
Replacement of l-Amino Acids by d-Amino Acids in the Antimicrobial Peptide Ranalexin and Its Consequences for Antimicrobial Activity and Biodistribution




Abstract
:1. Introduction
2. Results
2.1. Peptide Synthesis
2.2. Antimicrobial Susceptibility of Clinical Isolates
2.3. Antimicrobial Activity of Ranalexin and Danalexin
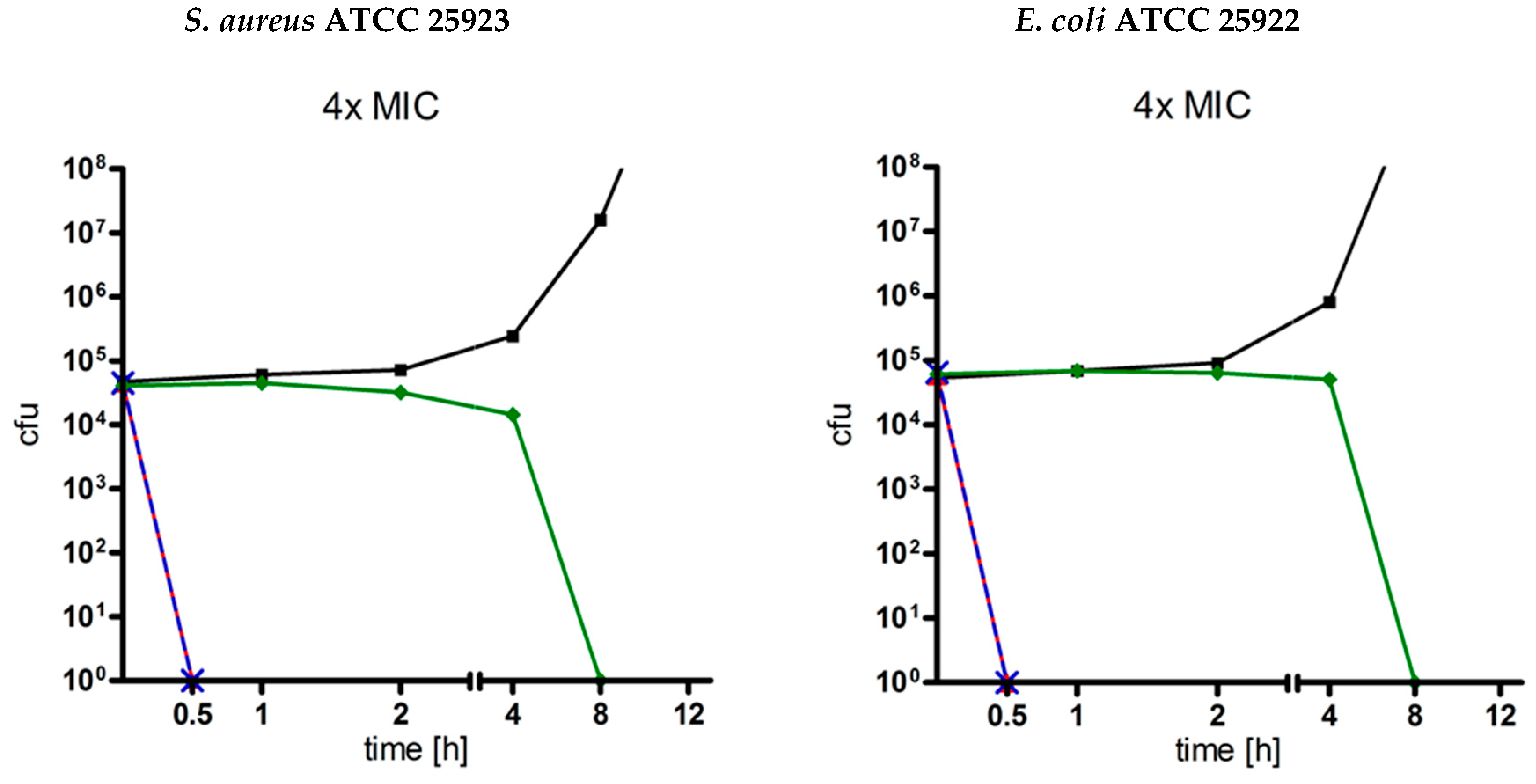
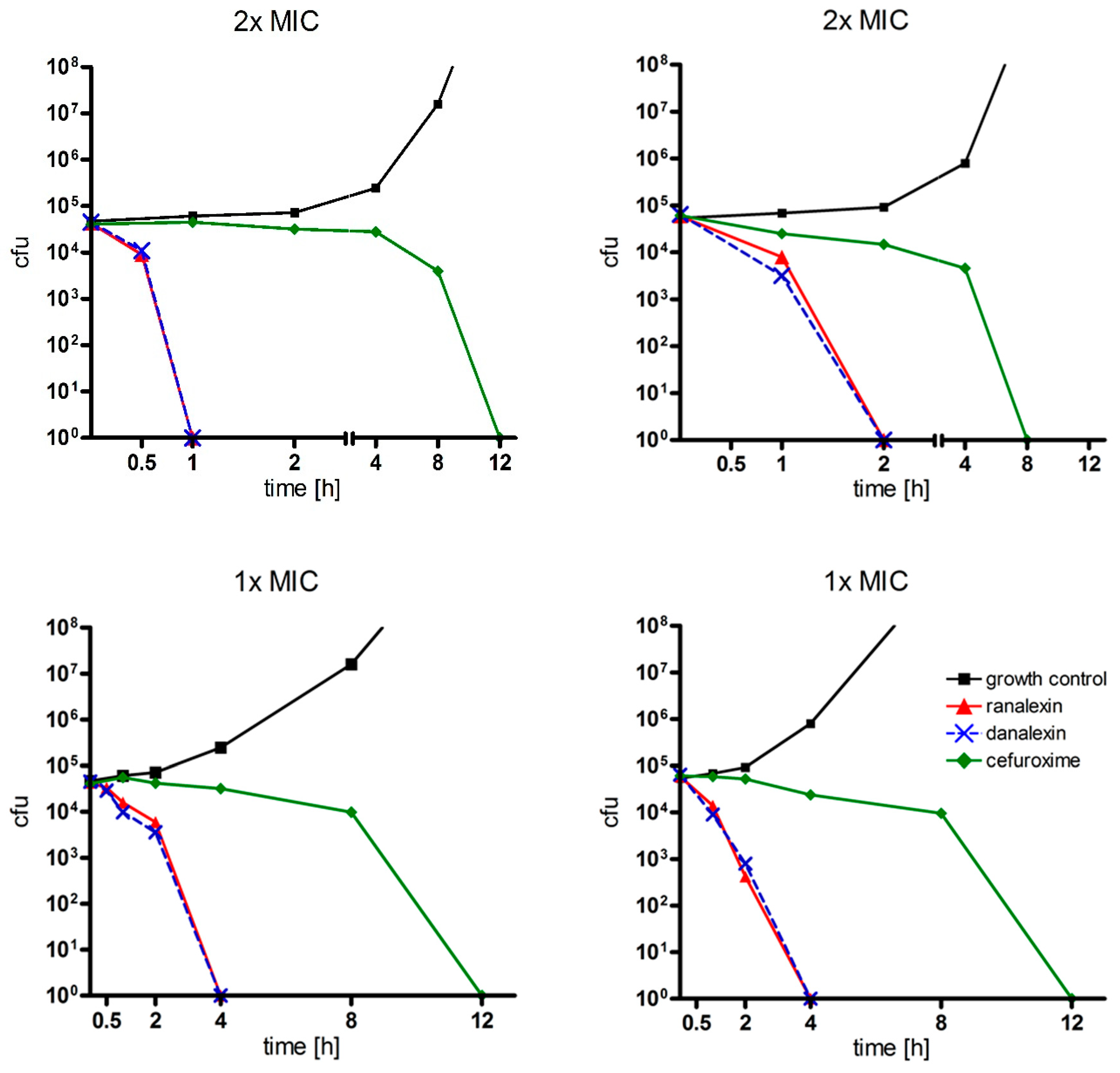
2.4. Time-Kill Curves
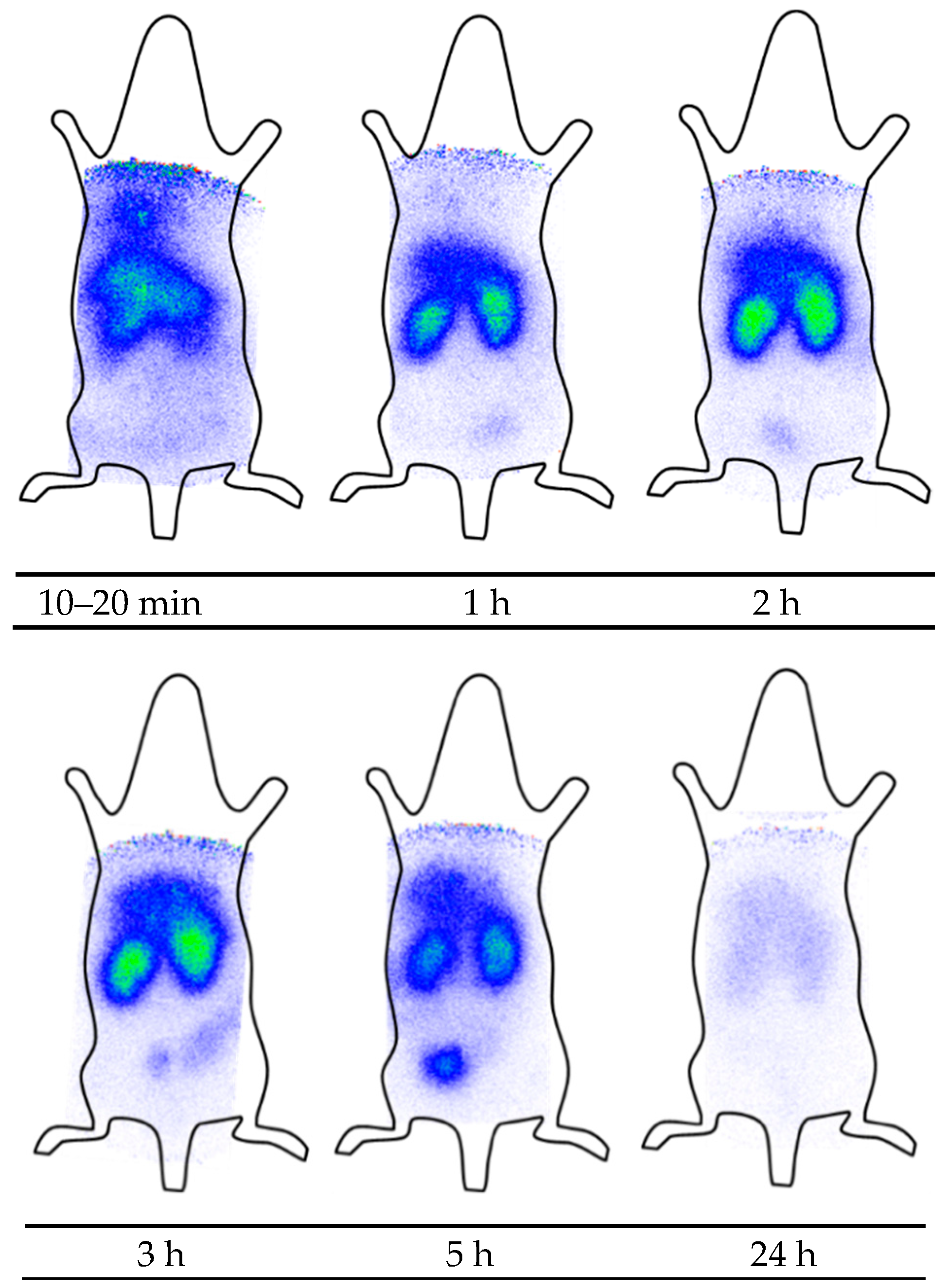
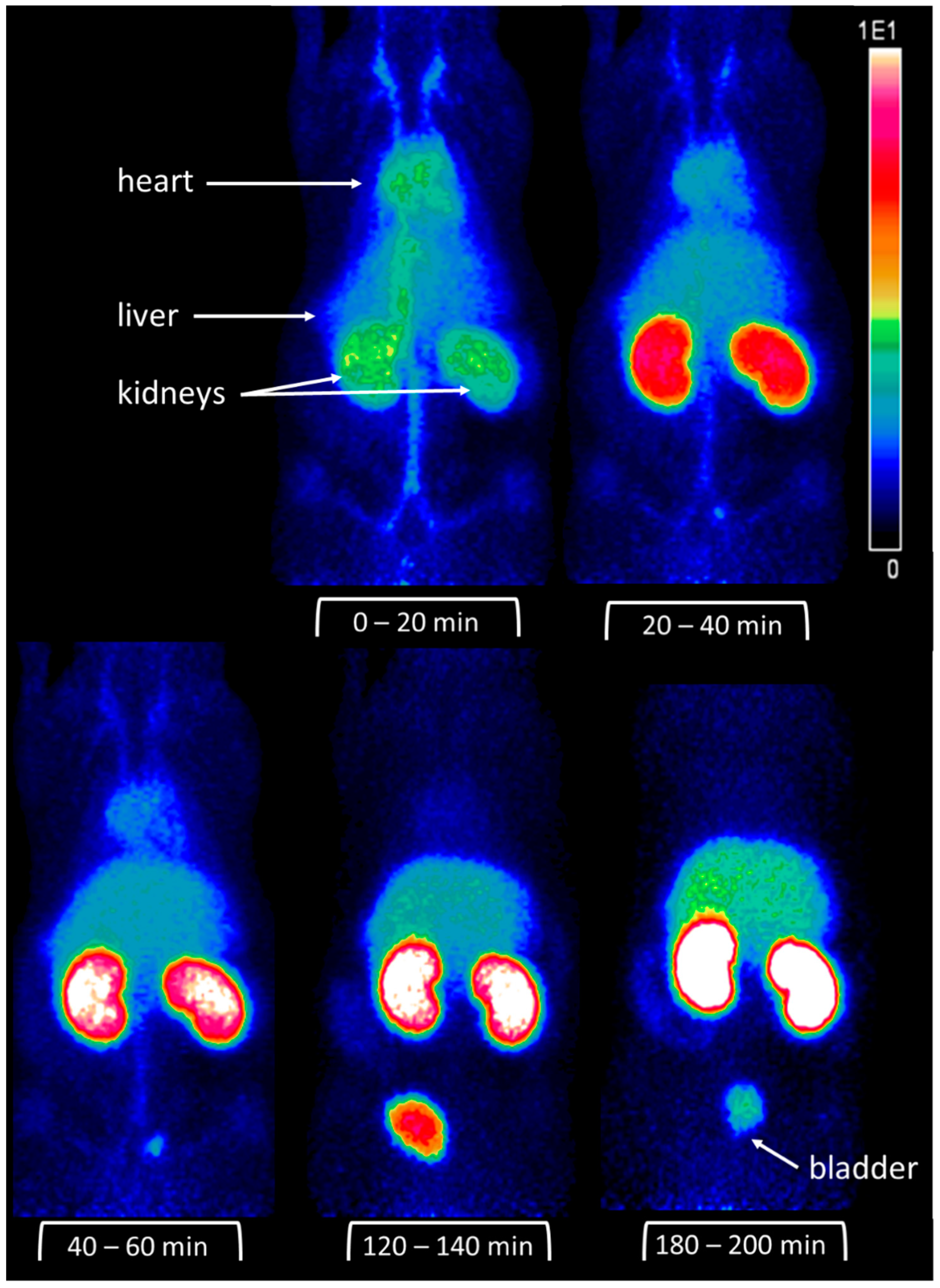
2.5. Scintigraphy
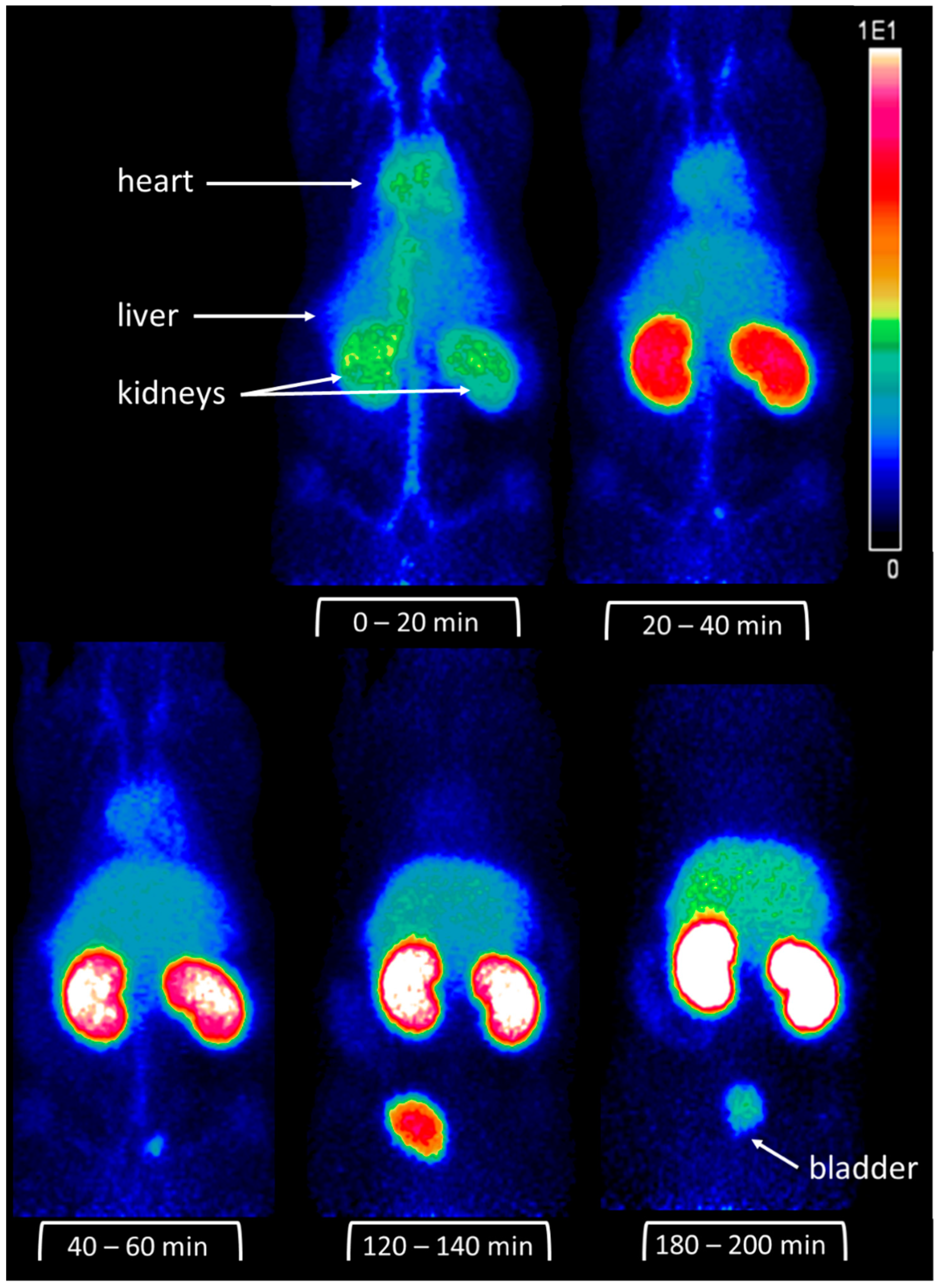
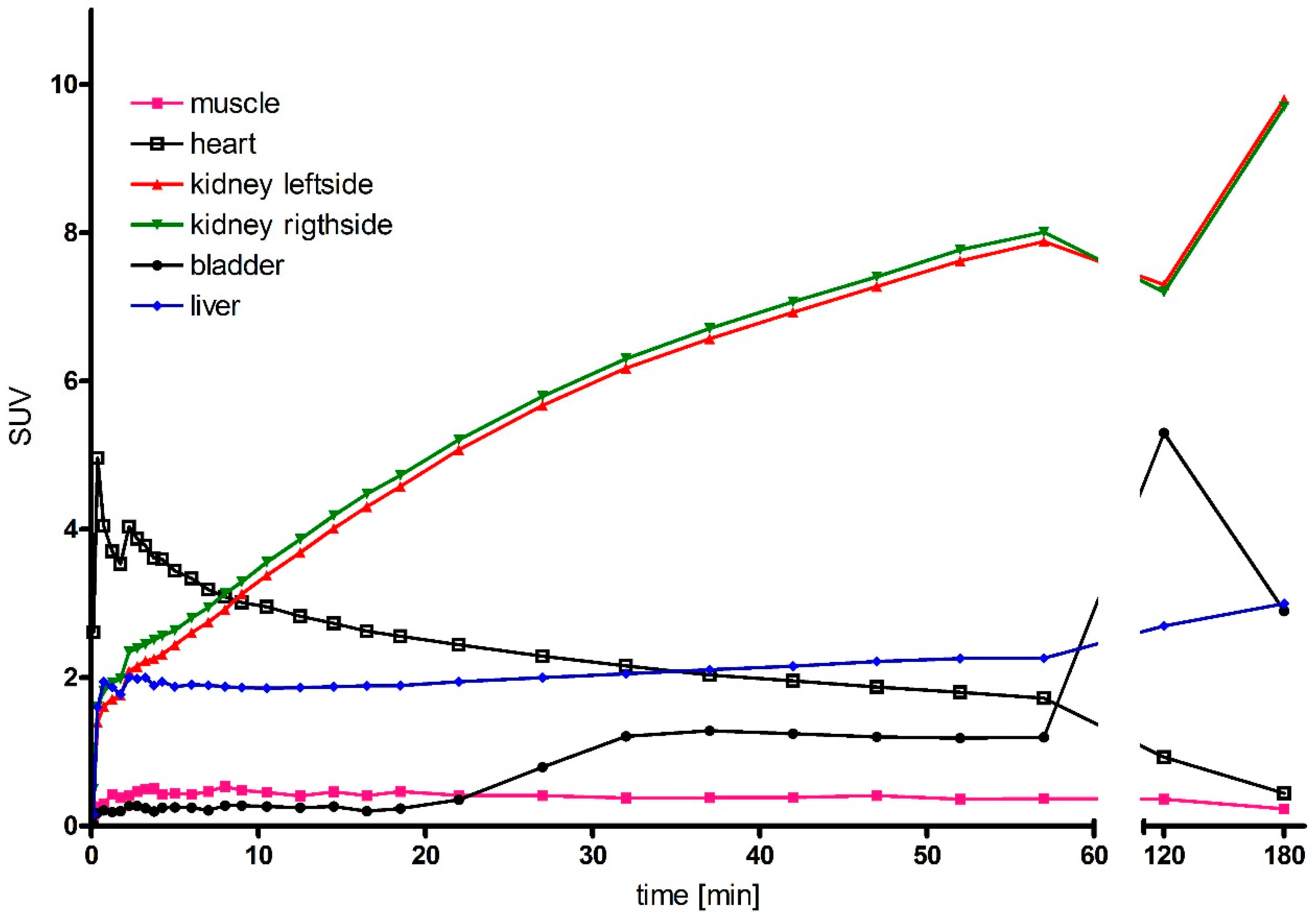
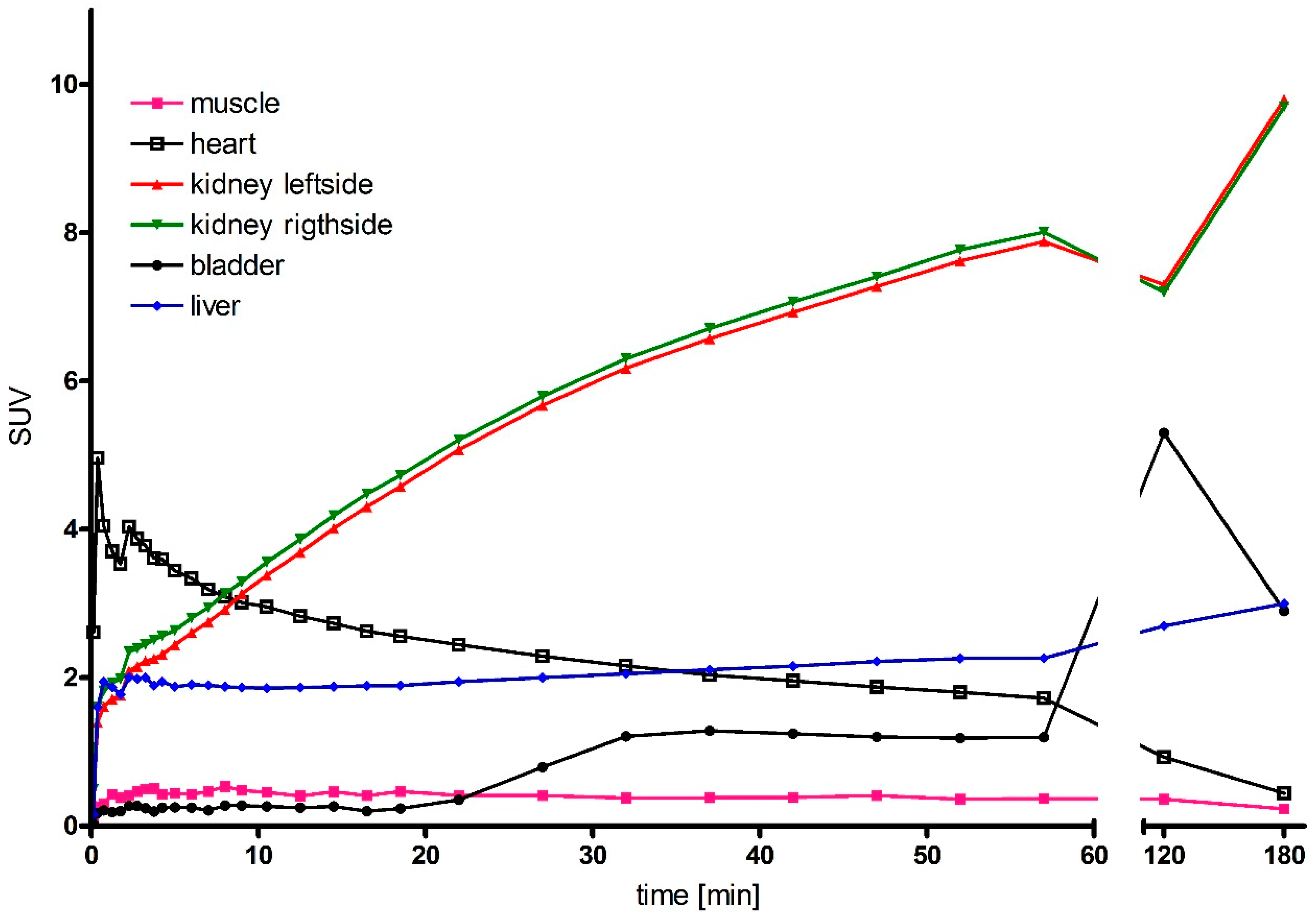
2.6. Micro-PET Imaging
3. Discussion
4. Material and Methods
4.1. Peptide Synthesis
4.2. Antimicrobial Activity
4.3. Time-Kill Curves
4.4. Radioactive Labeling and In Vivo Imaging
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Therap. 2015, 40, 277–283. [Google Scholar]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.L. Antimicrobial peptides stage a comeback. Nat. Biotechnol. 2013, 31, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.P.; Durell, S.; Maloy, W.L.; Zasloff, M. Ranalexin. A novel antimicrobial peptide from bullfrog (Rana catesbeiana) skin, structurally related to the bacterial antibiotic, polymyxin. J. Bio. Chem. 1994, 269, 10849–10855. [Google Scholar]
- Desbois, A.P.; Sattar, A.; Graham, S.; Warn, P.A.; Coote, P.J. MRSA decolonization of cotton rat nares by a combination treatment comprising lysostaphin and the antimicrobial peptide ranalexin. J. Antimicrob. Chemother. 2013, 68, 2569–2575. [Google Scholar] [CrossRef] [Green Version]
- Kosikowska, P.; Lesner, A. Antimicrobial peptides (AMPs) as drug candidates: A patent review (2003–2015). Expert. Opin. Ther. Pat. 2016, 26, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Kim, C.; Seo, C.H.; Park, Y. The therapeutic applications of antimicrobial peptides (AMPs): A patent review. J. Microbiol. 2017, 55, 1–12. [Google Scholar] [CrossRef]
- Hancock, R.E.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Hancock, R.E.; Chapple, D.S. Peptide antibiotics. Antimicrob. Agents Chemother. 1999, 43, 1317–1323. [Google Scholar] [CrossRef]
- Wakabayashi, H.; Matsumoto, H.; Hashimoto, K.; Teraguchi, S.; Takase, M.; Hayasawa, H. N-Acylated and D enantiomer derivatives of a nonamer core peptide of lactoferricin B showing improved antimicrobial activity. Antimicrob. Agents Chemother. 1999, 43, 1267–1269. [Google Scholar] [CrossRef]
- Domhan, C.; Uhl, P.; Meinhardt, A.; Zimmermann, S.; Kleist, C.; Lindner, T.; Leotta, K.; Mier, W.; Wink, M. A novel tool against multiresistant bacterial pathogens: Lipopeptide modification of the natural antimicrobial peptide ranalexin for enhanced antimicrobial activity and improved pharmacokinetics. Int. J. Antimicrob. Agents 2018, 52, 52–62. [Google Scholar] [CrossRef]
- CLSI. M100-S24: Performance Standards for Antimicrobial Susceptibility Testing. Available online: https://clsi.org/media/2663/m100ed29_sample.pdf (accessed on 15 July 2019).
- Zoller, F.; Schwaebel, T.; Markert, A.; Haberkorn, U.; Mier, W. Engineering and functionalization of the disulfide-constrained miniprotein min-23 as a scaffold for diagnostic application. ChemMedChem 2012, 7, 237–247. [Google Scholar] [CrossRef]
- Aleinein, R.A.; Hamoud, R.; Schafer, H.; Wink, M. Molecular cloning and expression of ranalexin, a bioactive antimicrobial peptide from Rana catesbeiana in Escherichia coli and assessments of its biological activities. Appl. Microbiol. Biotechnol. 2013, 97, 3535–3543. [Google Scholar] [CrossRef]
- Matsuzaki, K. Control of cell selectivity of antimicrobial peptides. Biochim. Biophys. Acta 2009, 1788, 1687–1692. [Google Scholar] [CrossRef] [Green Version]
- Papo, N.; Shahar, M.; Eisenbach, L.; Shai, Y. A novel lytic peptide composed of DL-amino acids selectively kills cancer cells in culture and in mice. J. Biol. Chem. 2003, 278, 21018–21023. [Google Scholar] [CrossRef]
- Zapotoczna, M.; Forde, E.; Hogan, S.; Humphreys, H.; O’Gara, J.P.; Fitzgerald-Hughes, D.; Devocelle, M.; O’Neill, E. Eradication of Staphylococcus aureus biofilm infections using synthetic antimicrobial peptides. J. Infect. Dis. 2017, 215, 975–983. [Google Scholar] [CrossRef]
- Thorne, K.J.; Thornley, M.J.; Glauert, A.M. Chemical analysis of the outer membrane and other layers of the cell envelope of Acinetobacter sp. J. Bacteriol. 1973, 116, 410–417. [Google Scholar]
- Jerala, R. Synthetic lipopeptides: A novel class of anti-infectives. Expert. Opin. Investig. Drugs 2007, 16, 1159–1169. [Google Scholar] [CrossRef]
- Wischnjow, A.; Sarko, D.; Janzer, M.; Kaufman, C.; Beijer, B.; Brings, S.; Haberkorn, U.; Larbig, G.; Kubelbeck, A.; Mier, W. Renal targeting: Peptide-based drug delivery to proximal tubule cells. Bioconjug. Chem. 2016, 27, 1050–1057. [Google Scholar] [CrossRef]
- Brings, S.; Fleming, T.; De Buhr, S.; Beijer, B.; Lindner, T.; Wischnjow, A.; Kender, Z.; Peters, V.; Kopf, S.; Haberkorn, U.; et al. A scavenger peptide prevents methylglyoxal induced pain in mice. Biochim. Biophys. Acta 2017, 1863, 654–662. [Google Scholar] [CrossRef]
- EUCAST. Determination of minimum inhibitory concentrations (MICs) of antimicrobial agents by broth dilution. Clin. Microbiol. Infect. 2003, ix–xv. [Google Scholar]
- CLSI. M11-A6: Methods for susceptibility testing of anaerobic bacteria. Available online: https://infostore.saiglobal.com/en-au/Standards/CLSI-M11-A6-6ED-2004-357434_SAIG_CLSI_CLSI_814197/ (accessed on 15 July 2019).
- Klein, S.; Zimmermann, S.; Kohler, C.; Mischnik, A.; Alle, W.; Bode, K.A. Integration of matrix-assisted laser desorption/ionization time-of-flight mass spectrometry in blood culture diagnostics: A fast and effective approach. J. Med. Microbiol. 2011, 61, 323–331. [Google Scholar] [CrossRef]
- Hofko, M.; Mischnik, A.; Kaase, M.; Zimmermann, S.; Dalpke, A.H. Detection of carbapenemases by Real-Time PCR and melt curve analysis on the BD max system. J. Clin. Microbiol. 2014, 52, 1701–1704. [Google Scholar] [CrossRef]
- NCCLS. M26: Methods for Determining Bactericidal Activity of Antimicrobial Agents. Available online: https://clsi.org/media/1462/m26a_sample.pdf (accessed on 15 July 2019).
- Uhl, P.; Helm, F.; Hofhaus, G.; Brings, S.; Kaufman, C.; Leotta, K.; Urban, S.; Haberkorn, U.; Mier, W.; Fricker, G. A liposomal formulation for the oral application of the investigational hepatitis B drug Myrcludex B. Eur. J. Pharm. Biopharm. 2016, 103, 159–166. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds tested in this study are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Amino Acid Sequence | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ranalexin | F | L | G | G | L | I | K | I | V | P | A | M | I | C | A | V | T | K | K | C |
| Ranalexin-d-Tyr | F | L | G | G | L | I | K | I | V | P | A | M | I | C | A | V | y1 | K | K | C |
| Danalexin | f | l | G | G | l | i | k | i | v | p | a | m | i | c | a | v | t | k | k | c |
| d-Tyr-danalexin | Y 1 | l | G | G | l | i | k | i | v | p | a | m | i | c | a | v | t | k | k | c |
| DOTA-d-Tyr-danalexin | y | l | G | G | l | i | k | i | v | p | a | m | i | c | a | v | t | k | K 2 | c |
| Peptide | Calculated Mass [Da] | Observed Mass [Da] | Detected Species |
|---|---|---|---|
| Ranalexin | 2103.1890 | 2104.1768 | [M + H]+ |
| Ranalexin-d-Tyr | 2165.0267 | 1083.6002 | [M + 2H]2+ |
| Danalexin | 2103.1890 | 2104.1047 | [M + H]+ |
| d-Tyr-danalexin | 2119.1839 | 2120.1700 | [M + H]+ |
| DOTA-d-Tyr-danalexin | 2506.0098 | 2506.1562 | [M]+ |
| A. baumannii SC303336 4-MRGN | A. baumannii SC411190 4-MRGN, OXA-23 | E. faecium UL407074 VanA | K. pneumoniae BL809453 4-MRGN, KPC | |
|---|---|---|---|---|
| Amoxicillin/Clavulanic acid | nt | R | R | nt |
| Piperacillin | R | R | nt | R |
| Piperacillin/Tazobactam | R | R | nt | R |
| Cefuroxime | nt | nt | R | R |
| Imipenem | R | R | R | R |
| Meropenem | R | R | nt | R |
| Ciprofloxacin | R | R | R | R |
| Gentamicin | R | R | nt | I |
| Tobramycin | R | R | nt | R |
| Amikacin | R | R | nt | nt |
| Tigecycline | I | nt | S | S |
| Trimethoprim/Sulfamethoxazole | R | R | R | R |
| Vancomycin | nt | nt | R | nt |
| Teicoplanin | nt | nt | R | nt |
| Erythromycin | nt | nt | R | nt |
| Linezolid | nt | nt | S | nt |
| Colistin | S | S | nt | S |
| Bacterium | MIC [mg/L] (µM) | ||
|---|---|---|---|
| Ranalexin | Danalexin | Positive Control | |
| Gram-positive bacteria | |||
| Bacillus megaterium DSM 32 | 4 (1.9) | 4 (1.9) | vancomycin 0.13 |
| B. subtilis DSM 10 | 4 (1.9) | 4 (1.9) | vancomycin 0.13 |
| Clostridium pasterianum DSM 525 | 16 (7.6) | 8 (3.8) | vancomycin 0.25 |
| Corynebacterium spheniscorum DSM 44757 | 16 (7.6) | 8 (3.8) | vancomycin 0.50 |
| Enterococcus casseliflavus ATCC 700327 VanC 1 | 8 (3.8) | 8 (3.8) | vancomycin 8 |
| E. faecalis ATCC 29212 | 16 (7.6) | 16 (7.6) | vancomycin 1 |
| E. faecium UL4070742 VanA 3 | 16 (7.6) | 8 (3.8) | vancomycin 640 |
| Staphylococcus aureus ATCC 25923 | 8 (3.8) | 4 (1.9) | vancomycin 1 |
| S. aureus NCTC 10442 MRSA 4 | 8 (3.8) | 8 (3.8) | vancomycin 1 |
| S. epidermidis ATCC 14990 | 16 (7.6) | 16 (7.6) | vancomycin 2 |
| S. saprophyticus ATCC 15305 | 8 (3.8) | 16 (7.6) | vancomycin 2 |
| Gram-negative bacteria | |||
| Acinetobacter baumannii SC3033362 4-MRGN 5 | 4 (1.9) | 4 (1.9) | colistin 0.25 6 |
| A. baumannii SC3223332 4-MRGN 5 | 8 (3.8) | 16 (7.6) | colistin 1 6 |
| A. baumannii SC4111902 4-MRGN 5 | 4 (1.9) | 8 (3.8) | colistin 0.25 6 |
| Escherichia coli ATCC 25922 | 32 (15.2) | 32 (15.2) | colistin 0.25 6 |
| E. coli 0157:H7 ATCC 35150 EHEC 7 | 32 (15.2) | 32 (15.2) | colistin 0.50 6 |
| Klebsiella pneumoniae ATCC 700603 | >64 (>30.4) | >64 (>30.4) | colistin 1 6 |
| K. pneumoniae BL809453 2 | >64 (>30.4) | >64 (>30.4) | colistin 0.25 6 |
| Pseudomonas aeruginosa ATCC 27853 | 64 (30.4) | 64 (30.4) | colistin 0.25 6 |
| P. fluorescens DSM 50090 | >64 (>30.4) | >64 (>30.4) | doxycycline 0.50 |
| Yersinia mollaretii DSM 18520 | >64 (>30.4) | >64 (>30.4) | colistin 0.25 6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domhan, C.; Uhl, P.; Kleist, C.; Zimmermann, S.; Umstätter, F.; Leotta, K.; Mier, W.; Wink, M. Replacement of l-Amino Acids by d-Amino Acids in the Antimicrobial Peptide Ranalexin and Its Consequences for Antimicrobial Activity and Biodistribution. Molecules 2019, 24, 2987. https://doi.org/10.3390/molecules24162987
Domhan C, Uhl P, Kleist C, Zimmermann S, Umstätter F, Leotta K, Mier W, Wink M. Replacement of l-Amino Acids by d-Amino Acids in the Antimicrobial Peptide Ranalexin and Its Consequences for Antimicrobial Activity and Biodistribution. Molecules. 2019; 24(16):2987. https://doi.org/10.3390/molecules24162987
Chicago/Turabian StyleDomhan, Cornelius, Philipp Uhl, Christian Kleist, Stefan Zimmermann, Florian Umstätter, Karin Leotta, Walter Mier, and Michael Wink. 2019. "Replacement of l-Amino Acids by d-Amino Acids in the Antimicrobial Peptide Ranalexin and Its Consequences for Antimicrobial Activity and Biodistribution" Molecules 24, no. 16: 2987. https://doi.org/10.3390/molecules24162987
APA StyleDomhan, C., Uhl, P., Kleist, C., Zimmermann, S., Umstätter, F., Leotta, K., Mier, W., & Wink, M. (2019). Replacement of l-Amino Acids by d-Amino Acids in the Antimicrobial Peptide Ranalexin and Its Consequences for Antimicrobial Activity and Biodistribution. Molecules, 24(16), 2987. https://doi.org/10.3390/molecules24162987