1. Introduction
Inflammatory skin lesions caused by various bacterial infectious diseases, such as acne, folliculitis, furuncles, and carbuncles, mainly occur on the epidermis as nodules, pustules, and papules [
1,
2,
3,
4]. Reactive oxygen species (ROS) have been found to contribute to epidermal inflammation by damaging the follicular epithelium and inducing melanin deposition [
1]. The goal of epidermal inflammation treatment is to control existing lesions and prevent permanent scarring as much as possible [
2]. Epidermal inflammation therapies combine systematic and local treatments such as non-steroid anti-inflammatory drugs (NSAIDs), retinoids, antibiotics, hormonal therapy, and other complementary medicines [
5]. These medicines generally regulate the epidermal cell growth, reduce hyperpigmentation, and have anti-inflammatory, anti-bacterial, and anti-oxidative effects. In order to achieve a good therapeutic effect, it is essential to use various medicines cooperatively [
6]. Therefore, it will be reasonable to search for an alternative drug with both anti-inflammatory and anti-oxidative effects to protect the epidermis from infectious skin diseases, rather than using the current complex drug combinations.
Feruloylserotonin (FS;
N-feruloyl-5-hydroxy-tryptamine) is isolated from safflower seeds and has been shown to have anti-atherogenic, anti-oxidative, and anti-melanogenesic activities [
7,
8]. FS is comprised of 5-hydroxytryptamine (5-HT) and ferulic acid which may contribute to its anti-oxidative and anti-inflammatory properties [
9]. It has been reported that the combined therapy of FS and methotrexate can reduce inflammation in adjuvant arthritis [
10]. Nuclear factor-κB (NF-κB) is an important transcriptional regulator of inflammatory damage that is often considered to be a therapeutic target for inflammatory diseases. FS can inhibit NF-κB activation in human aortic endothelial cells treated with high levels of glucose [
11]; however, the anti-inflammatory effects of FS were only proven by detecting its anti-inflammatory efficiency. No detailed mechanism has yet been elucidated for the action of FS against LPS (lipopolysaccharides) induced epidermal inflammation. It has also been found through DCF-DA and XTT (2,3-
bis(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazxolium-5-carboxanilide inner salt) assays that FS can inhibit oxidative stress in response to cytokines, bacterial invasion, and other factors [
7]. The anti-oxidative activity of FS can also inhibit H
2O
2-induced melanogenesis by down-regulating the microphthalmia-associated transcription factor expression via extracellular signal-regulated kinase phosphorylation [
12]. Nrf2 (nuclear factor erythroid 2-related factor 2) is thought to be a master regulator of cellular survival, whose translocation during oxidative stress induces genes to produce antioxidant enzymes. Although Nrf2 is an important regulator that controls many pathways in response to oxidative stress, there is no direct evidence that it is affected by FS. Both NF-κB and Nrf2 are important transcription factors in almost all cell lines that regulate cellular responses to inflammation and oxidation, respectively. Since functional cross-talk exists between these two pathways, specific anti-inflammatory or anti-oxidative functions must be confirmed.
Keratinocytes comprise almost 95% of the cell mass of the human epidermis, the inflammation of which is considered as skin problem [
13]. LPS are the main component of Gram-negative bacterial membranes and are one of the most common stimulators of inflammation [
14]. In this study, we established an epidermal bacterial infection inflammatory model in human keratinocyte HaCaT cells treated with LPS. In order to determine the effects of FS on epidermal inflammation, the anti-inflammatory and anti-oxidative properties of FS was assessed in LPS-induced HaCaT cells. Evidence was provided that FS not only inhibits the Toll-like receptor 4-mediated nuclear factor-κB (TLR4/NF-κB) pathway, but also promotes the translocation and stability of Nrf2 by analyzing cell ability, prostaglandin E2 (PGE
2) release, ROS production, cell apoptosis, TLR4/NF-κB-related protein expression, and Nrf2 translocation. FS can be used as a component of topical drugs to manage inflammatory skin lesions in infectious diseases because of its anti-oxidative and anti-inflammatory properties.
3. Discussion
FS is a compound extracted from safflower seeds that exhibits anti-oxidative and anti-inflammatory properties. Here, we examined these two activities of FS in LPS-induced HaCaT cells.
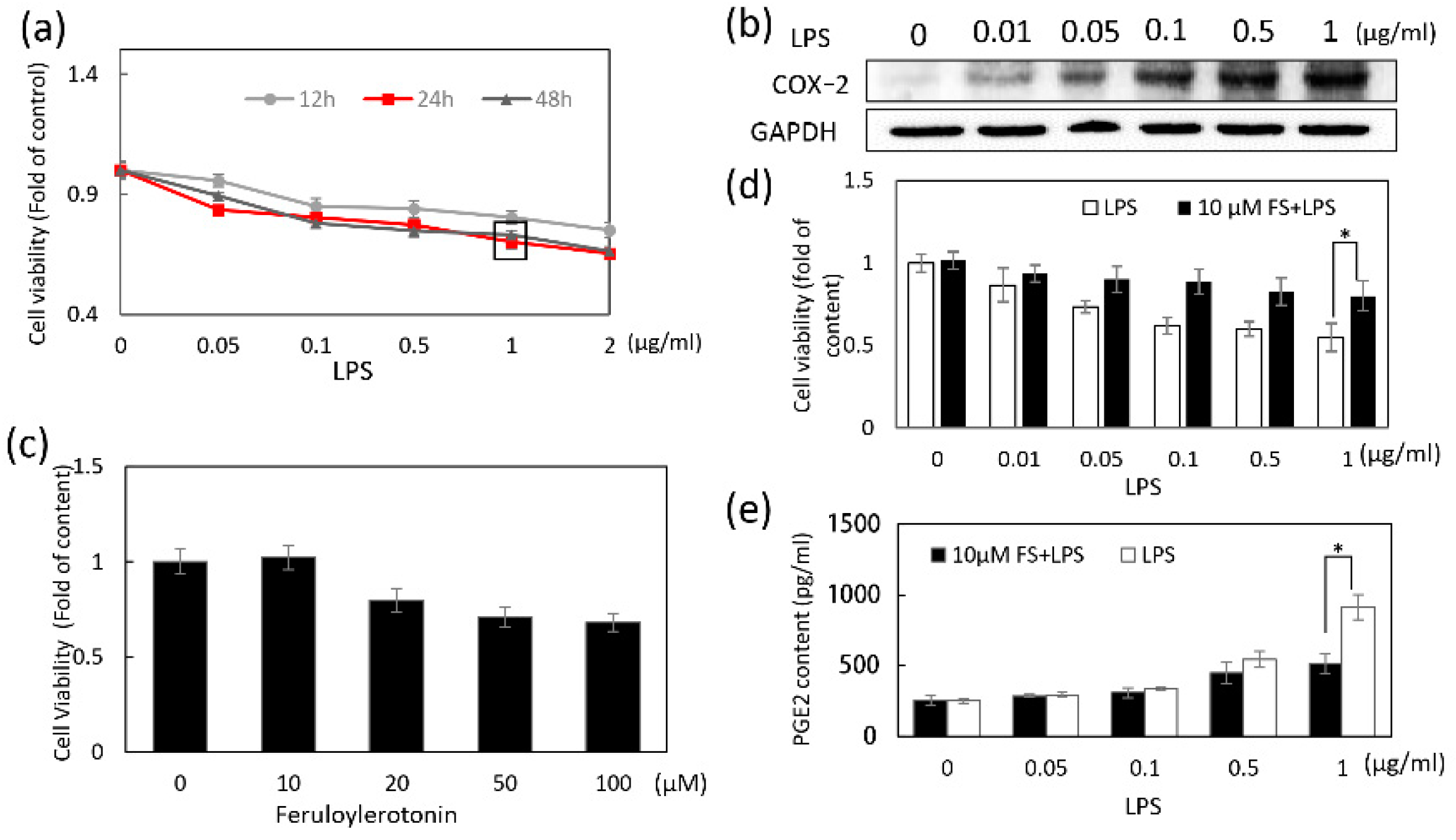
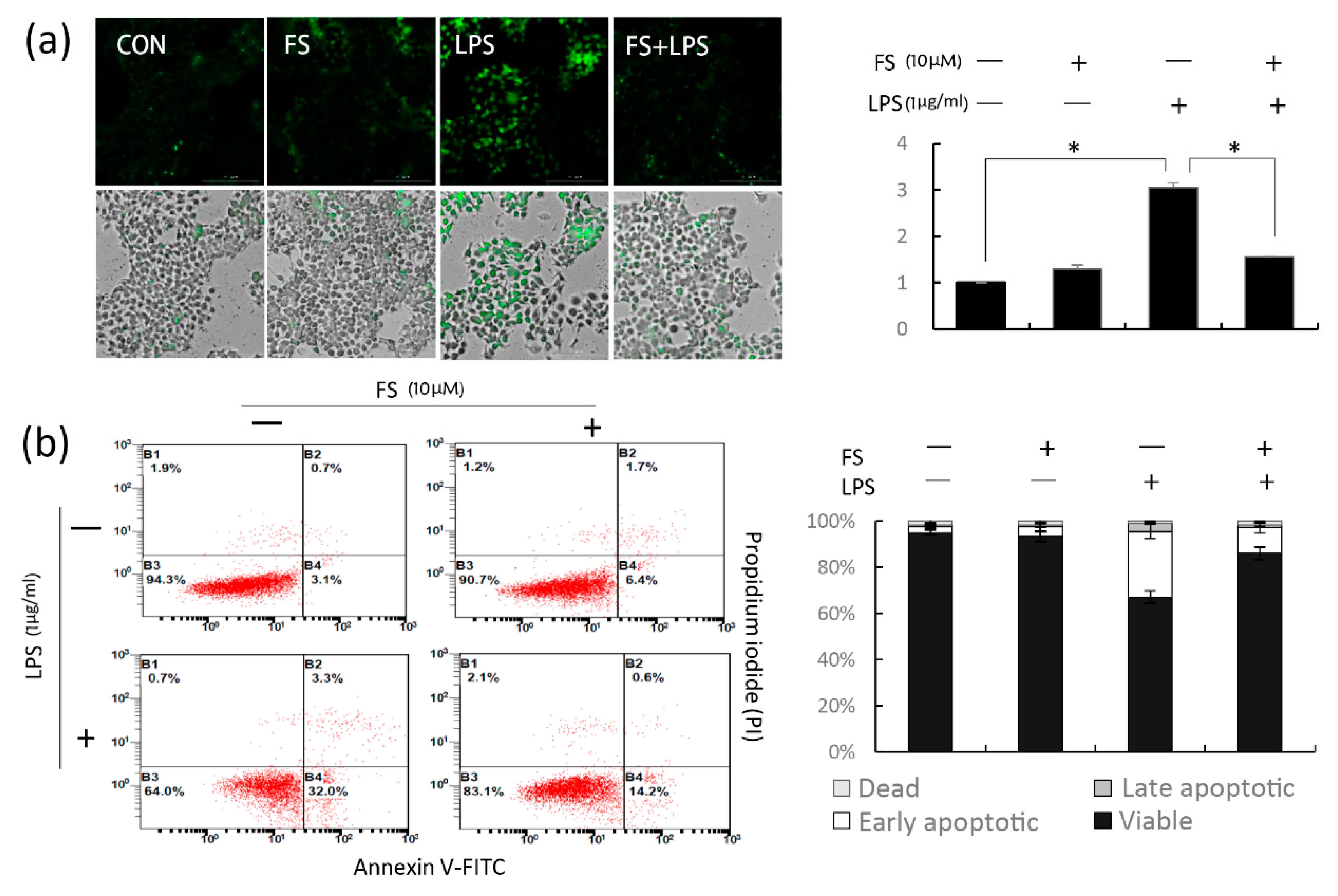
The viability of HaCaT cells treated with 1 μg/mL LPS for 24 h was approximately 60%; however, the reduction in apoptosis because of FS was relevant in this study. LPS reduced the viability of pretreated cells with or without FS in a dose-dependent manner; however, the decrease was less in the FS-pretreated group than in the group without FS (
Figure 1d), indicating that FS pretreatment maintained cell stability. Similarly, flow cytometry result suggested that FS pretreatment reduce the LPS-induced apoptosis of HaCaT cells (
Figure 4a).
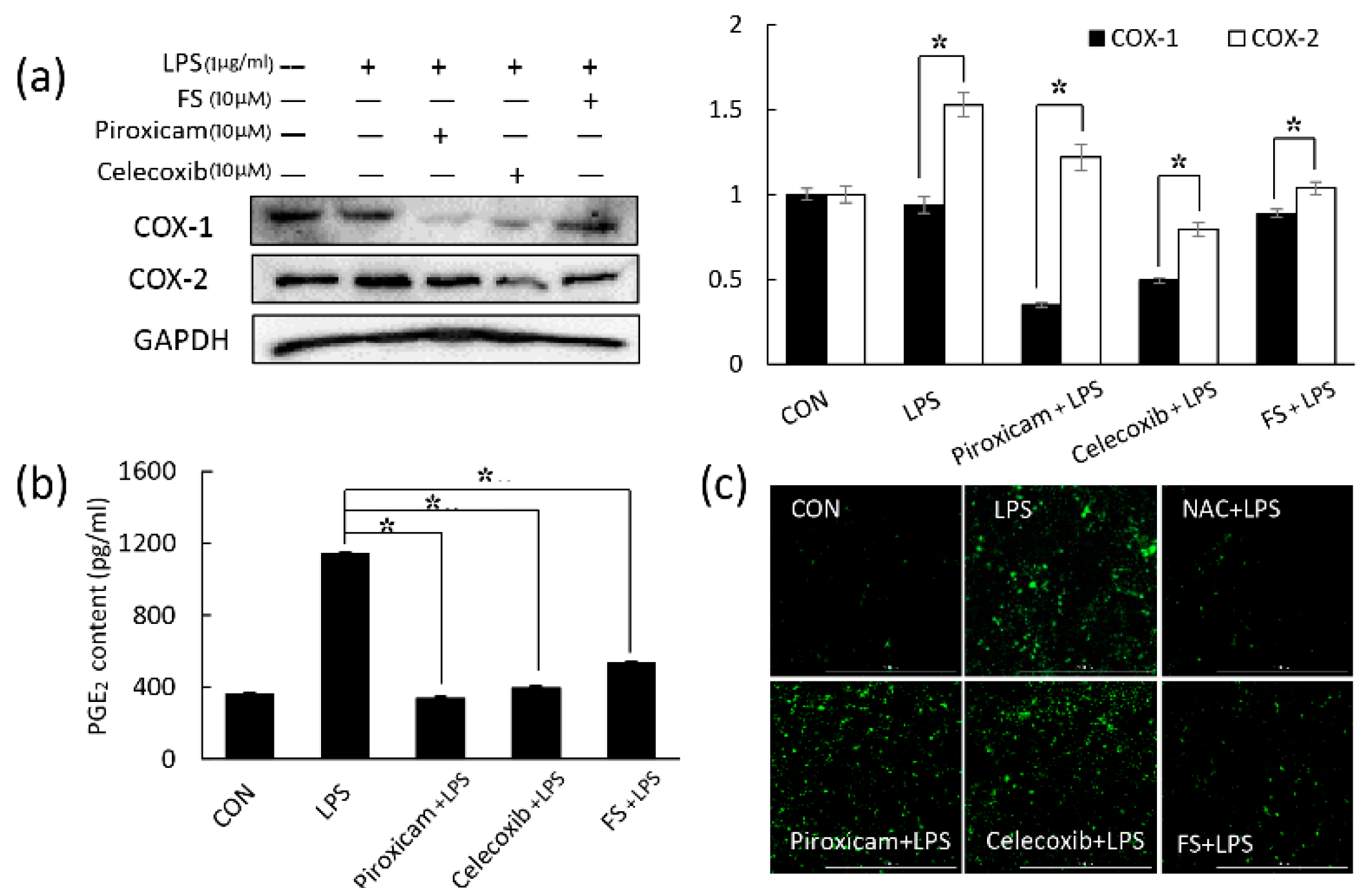
PGE
2 plays an important role in generating inflammatory responses and is induced by the inflammatory stimulator COX-2. COX-1 may also be a dominant source of PGE
2 for housekeeping functions [
16,
17]. The NSAIDs piroxicam and celecoxib were used to inhibit COX-1 and COX-2, respectively. FS did not affect COX-1 expression compared to piroxicam (
Figure 2a); therefore, we focused on COX-2 in this study. Western blotting and a PGE
2 assay revealed that FS exhibited a weaker anti-inflammatory activity than piroxicam and celecoxib (
Figure 2a,b), whereas the ROS assay suggested that FS exhibited better anti-oxidative effects than the two NASIDs (
Figure 2c). Thus, FS was found to have the potential to manage epidermal inflammation. LPS-induced COX-2 expression and PGE
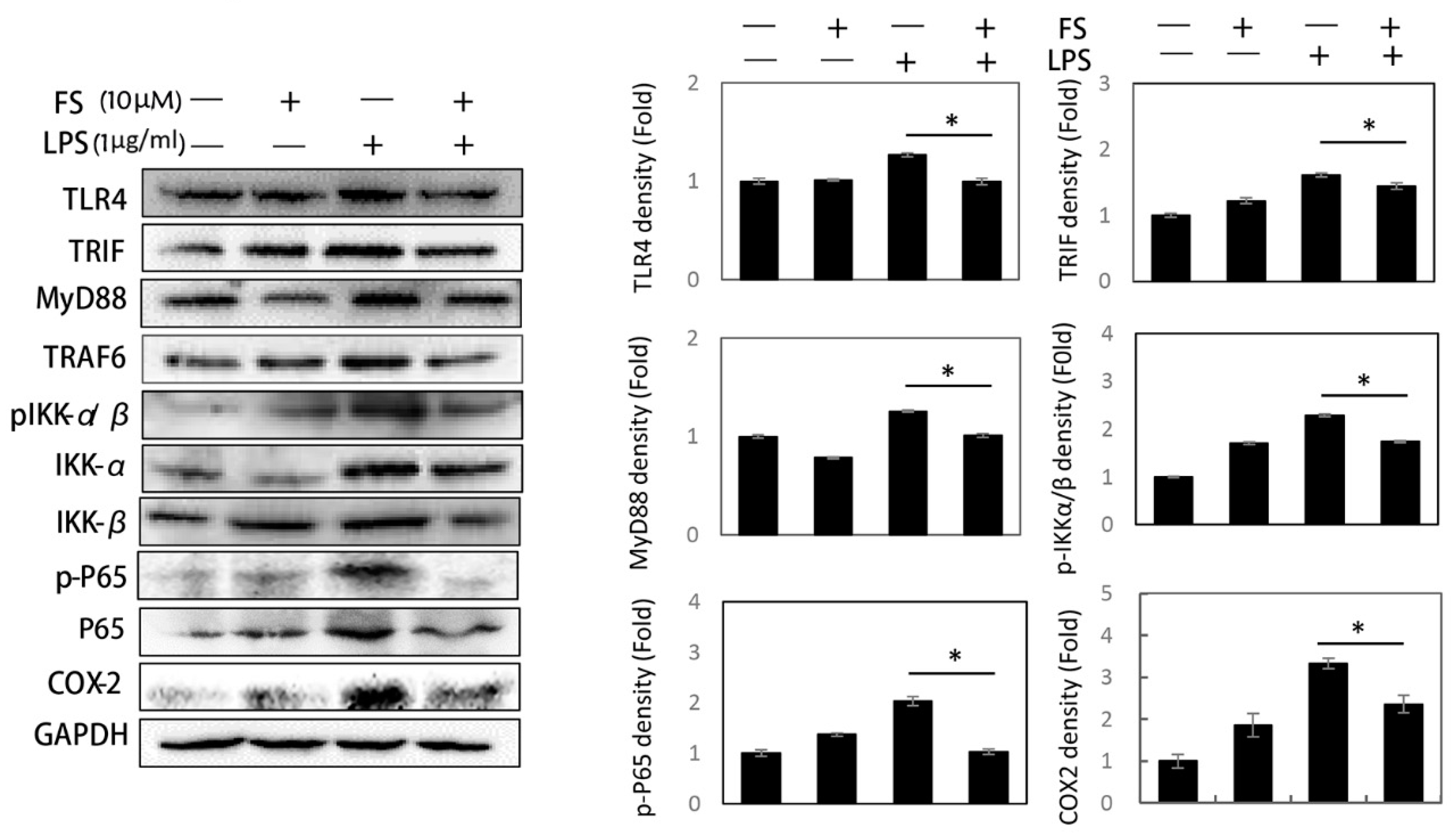
2 release occurred via the TLR4/NF-κB pathway and were both inhibited by FS pretreatment. It has been reported that LPS can induce TLR4 expression and lead to NF-κB pathway activation [
18]. The LPS-induced activation of the TLR4/NF-κB pathway was related to the following proteins: TLR4, MyD88, TRIF, TRAF6, pIKK-α/β, IKK-α, IKK-β, p-P65, and P65 (
Figure 3) [
19]. The FS pretreatment of HaCaT cells inhibited the expression of TLR4/NF-κB pathway proteins, suggesting that FS be beneficial on LPS-induced inflammation. Some protein expressions in FS group as a positive group were higher than that in control group. This difference was inferred to be caused by the basal condition of the HaCaT cells. Once the HaCaT cells were treated with FS, the basal condition was different from control group, followed by the changes of protein levels. In the experiment of detecting TLR4/NF-κB pathway, the difference between LPS group and FS-LPS group was remarkable.
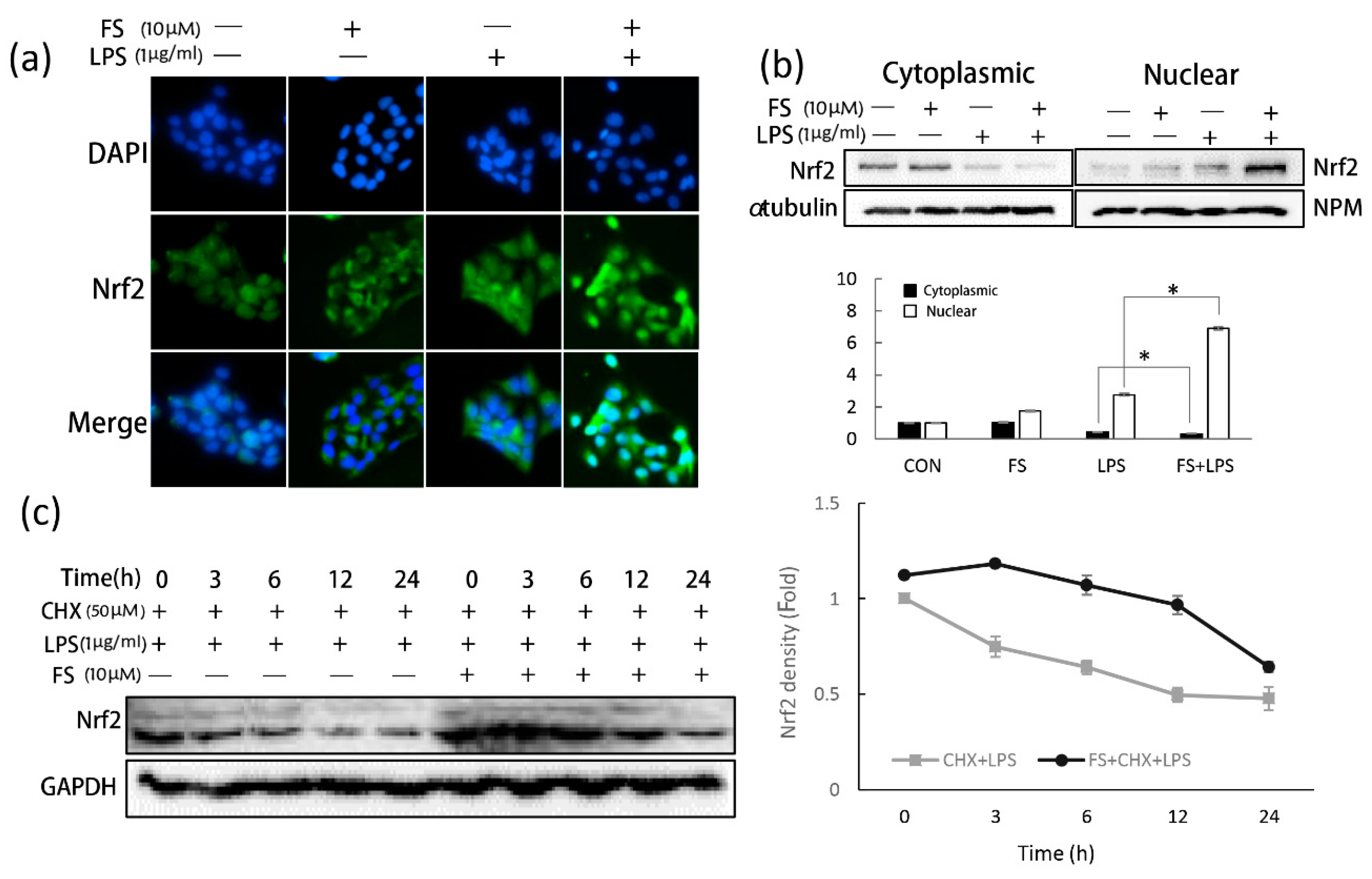
The anti-inflammatory effects of FS were confirmed by analyzing total ROS level (
Figure 4a). Nrf2 is thought to be a master regulator of cell survival [
20]. Under basal conditions, Nrf2 resides in the cytoplasm and translocates inactively. If cells experience ROS-induced oxidative damage, Nrf2 is actively translocated to the nucleus [
21]. FS exerted anti-oxidative effects by stimulating Nrf2 translocation. In LPS-treated HaCaT cells, Nrf2 translocated into the nucleus a little; however, when treated with both FS and LPS, nuclear Nrf2 levels increased greatly. In FS-pretreated HaCaT cells, Nrf2 accumulated around the nucleus; however, translocation did not occur until the HaCaT cells were exposed to LPS (
Figure 5a). These results suggest that when pretreated with FS, Nrf2 translocation was directly induced by LPS, with FS acting as a stimulator, concordant with the western blotting results (
Figure 5b). It was also shown that cytoplasmic Nrf2 levels were slightly higher in FS-pretreated HaCaT cells than in the control group. To explain this observation, CHX was used to block Nrf2 synthesis and investigate the role of FS on Nrf2 stability (
Figure 5c). CHX is often used to block messenger RNA translocation and thus block protein synthesis [
22]. Within 24 h, FS increased Nrf2 stability in CHX-blocked HaCaT cells, particularly when exposed to LPS for 3-12 h, indicating that FS prevented Nrf2 ubiquitination and degradation [
23].
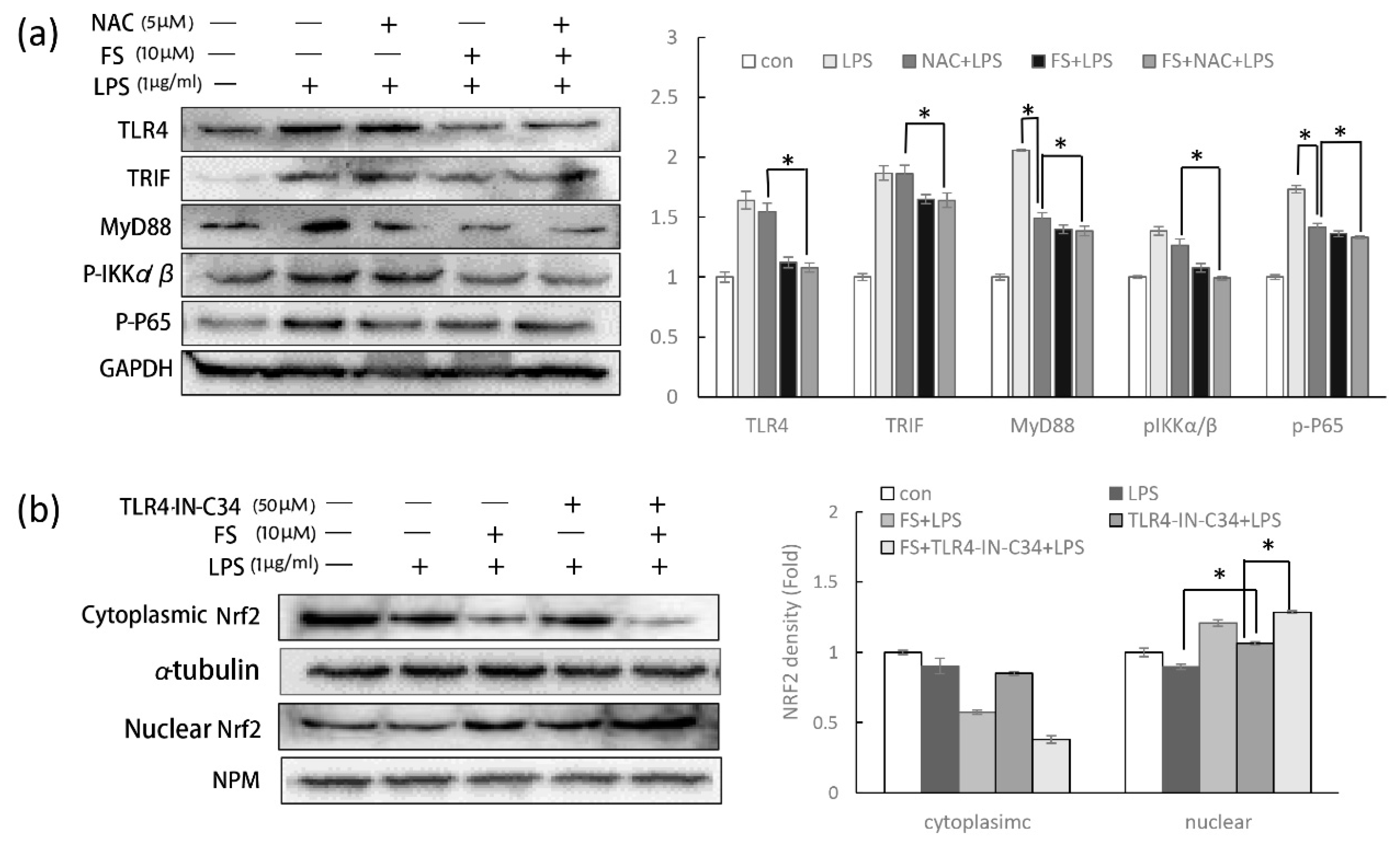
Furthermore, it has been reported that LPS-induced ROS can increase cellular inflammation, while ROS-induced Nrf2 transcription can produce anti-oxidant enzymes to reduce COX-2 production [
24]. To evade the complex relationship described above and detect the specific function of FS in the TLR4/NF-κB pathway, NAC was used to clear LPS-induced ROS. The cleared ROS avoid the effect of Nrf2 translocation on evaluating the action of FS on TLR4/NF-κB pathway. Notably, without the effect of Nrf2 on inflammation, FS still performed the activity on the anti-inflammation (
Figure 6a).
The anti-inflammatory role of Nrf2 in the TLR4/NF-κB pathway has been discussed above; however, it should be noted that NF-κB can also regulate Nrf2 activity. It has been well established that CREB-binding protein is a transcriptional co-activator of both Nrf2 and NF-κB, with NF-κB overexpression thought to limit Nrf2 complex formation by competing with CBP (CREB-binding protein) [
15]. In order to eliminate the effect of the TLR4/NF-κB pathway on Nrf2 translocation, TLR4 inhibitor was used. The Nrf2 translocation in HaCaT cells pretreated with FS and TLR4-IN-C34 was still promoted, suggesting that the promotion of FS on Nrf2 translocation is independent from its anti-inflammatory property (
Figure 6b). Nuclear Nrf2 level in the TLR4-C34-IN pretreated HaCaT cells were slightly lower than the group without TLR4-C34-IN, suggesting that Nrf2 translocation was slightly regulated by the activity of TLR4/NF-κB pathway.
In this study, the mechanism of FS on anti-inflammation and anti-oxidation was investigated via TLR4/NF-κB pathway and Nrf2 translocation. Although the inhibition of FS on TLR4/NF-κB pathway was proved, its target needs to be confirmed. FS has both anti-inflammatory and anti-oxidative properties compared to the existing NSAIDs, as shown in our results. Further investigation is required to see if FS can be used in clinical applications.
In summary, FS exhibited anti-inflammatory and anti-oxidative activities in LPS-induced human keratinocyte HaCaT cells by inhibiting the TLR4/NF-κB pathway and promoting Nrf2 translocation. These two properties benefited each other, but independently; thus, FS has the potential of being a component to manage epidermal inflammation because of its anti-inflammatory and anti-oxidative properties.
4. Materials and Methods
4.1. Chemicals and Antibodies
LPS was purchased from Sigma (Sigma-Aldrich, St. Louis, MO, USA). Specific antibodies for Nrf2, NPM, α-tubulin, GAPDH, COX2, TLR4, MyD88, TRAF6, and p-IKKα/β were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibodies for TRIF, IKK-α, IKK-β, NF-kB (P65), and pNF-kB (p-P65) were purchased from Cell Signaling Technology (Cell Signaling Technology, Beverly, MA, USA). FS was synthesized by reacting activated hydroxycinnamic acid esters with serotonin hydrochloride under alkaline conditions.
4.2. Cell Culture
HaCaT cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum (FBS; JR Scientific, Woodland, CA, USA) and penicillin/streptomycin (Wel gene, Gyeongsan, Korea) at 37 °C in a humid 5% CO2 atmosphere for 24 h. Cells were then pretreated with FS at the indicated concentration for 24 h and exposed to LPS (Sigma-Aldrich, St. Louis, MO, USA) for 24 h. To detect the effects of FS on Nrf2 stability, HaCaT cells were pretreated with or without FS for 24 h and 50 µM cycloheximide (CHX, Sigma-Aldrich, St. Louis, MO, USA) for 4 h. To inhibit the TLR4/NF-kB pathway, cells were pretreated with 50 µM TLR4 inhibitor (TLR4-IN-C34, Sigma-Aldrich, St. Louis, MO, USA) before LPS. To clear ROS, cells were pretreated with 5 µM N-acetyl-L-cysteine (NAC, Sigma-Aldrich, St. Louis, MO, USA) before LPS. To compare the anti-inflammatory activity of FS with NSAIDs, COX-1 inhibitor (Piroxicam) and COX-2 inhibitor (Celecoxib) were used to pretreat the HaCaT cells at a concentration of 10 µM. (The cells without any treatment: control group; only FS-treatment: FS group; merely LPS-treatment: LPS group; FS pretreatment and LPS treatment: FS-LPS group).
4.3. Cell Viability Assay
HaCaT cells (1 × 103 cells/well) were seeded in 96-well plates (Corning, NY, USA) with DMEM containing 10% FBS and 1% penicillin and streptomycin at 37 °C in a humid 5% CO2 atmosphere and treated with 0, 10, 20, 50, or 100 µM/mL FS for 24 h. MTT reagent (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (Sigma-Aldrich, St. Louis, MO, USA) was added to each well and incubated for a further 4 h. The MTT reagent was discarded, cells were incubated with DMSO for 10 min, and their absorbance was measured at 570 nm using a microplate reader (Bio-rad, Hercules, CA, USA). Viable cell percentage was calculated as follows: cell viability = mean absorbance in test well/mean absorbance in control well. The same method was used to assess the viability of HaCaT cells pretreated with 10 µM/mL FS and 0, 10, 50, 100, 500, or 1000 ng/mL LPS.
4.4. PGE2 Release Assay
HaCaT cells were pretreated with or without FS and treated with 0, 50, 100, 500, or 1000 ng/mL LPS for 24 h. All supernatants were then collected and their PGE2 levels were measured using a commercially available ELISA kit (R & D Systems, Minneapolis, MN, USA) with a microplate reader.
4.5. Cellular Reactive Oxygen Species (ROS) Detection
Total ROS production in HaCaT cells was determined by the 2′,7′-dichlorodihydrofluorescein diacetate (DCF-DA, Sigma-Aldrich, St. Louis, MO, USA) fluorescence method. Briefly, HaCaT cells (approximately 70% confluence in a 6-well plate) were incubated at 25 °C in PBS with DCF-DA away from light for 20 min. The PBS containing DCF-DA was then aspirated and cells were rinsed twice with cold PBS. The fluorescence of oxidized DCF-DA in the cell lysates, an index of ROS formation, was measured by fluorescence microscopy with 488 nm excitation and 530 nm emission filters (LionheartTM FX, Winoski, VT, USA).
4.6. Apoptosis Analysis
LPS-induced HaCaT cell apoptosis was analyzed by Annexin V-FITC/propidium iodide (PI) (Invitrogen, Carlsbad, CA, USA) staining. Appropriately treated HaCaT cells were harvested and washed in cold PBS, centrifuged, and re-suspended in annexin-binding solution to which a working solution of FITC-annexin V and PI was added. After 15 min incubation at 25 °C, the samples were immediately analyzed under single laser emitting excitation by flow cytometry.
4.7. Immunofluorescence Staining
To analyze intracellular Nrf2 localization, HaCaT cells were grown as polarized monolayers on 12 mm tissue culture inserts (Nunc, NY, USA), fixed with 4% paraformaldehyde in PBS for 20 min at 25 °C, and incubated with 0.1% Triton X-100 in PBS for 15 min. The cells were then incubated in 1% BSA-PBS for 1 h at 25 °C and with anti-Nrf2 antibodies (1:100 in 1% BSA-PBS, m/v) overnight at 4 °C. Cells were washed three times with 1% BSA-PBS followed by staining with goat anti-rabbit IgG secondary antibodies. And the nucleus was stained with diluted DAPI (Molecular Probes). The stained cells were mounted and imaged using a laser confocal scanning microscope (Lionheart FX, Winoski, VT, USA).
4.8. Western Blotting Analysis
At the indicated time points, HaCaT cells were lysed with modified radioimmunoprecipitation assay (RIPA) buffer (50 mM Tris-Cl, 150 mM NaCl, 1% Nonidet P-40, and 0.1% SDS) with PMSF and a protease inhibitor cocktail. Extracted proteins were subjected to 7.5% SDS-PAGE gel electrophoresis and transferred to polyvinylidene difluoride (PVDF) membranes (Millipore) followed by being blocked with 2% milk-TBS buffer at 25 °C for 1 h. After being blocked, the membranes were probed with primary antibodies at 4 °C overnight using the following antibodies related to the TLR4/NF-κB pathway and Nrf2: TLR4 (1:1000), MyD88 (1:500), TRIF (1:1000), TRAF6 (1:500), pIKK-α/β (1:1000), IKK-α (1:1000), IKK-β (1:1000), pNF-κB (1:1000), NF-κB (1:1000), COX-2 (1:500), GAPDH (1:1000), Nrf2 (1:1000), NPM(1:1000), and α-tubulin (1:1000). After being washed with TBST four times, the membranes were incubated with secondary anti-rabbit IgG (sc-2004) or anti-mouse IgG (sc-2005) antibodies for 2 h at 25 °C and visualized using an enhanced chemiluminescence kit.
4.9. Statistical Analysis
All the results were expressed as mean ± standard deviation. The statistical analysis was performed by one-way ANOVA test using SPSS software. All data were repeated at least three times. If p < 0.05, the result was considered significant difference.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





