
Mushrooms of the Genus Ganoderma Used to Treat Diabetes and Insulin Resistance



Abstract
1. Introduction
2. Hypoglicemic Activity of Ganoderma Extracts
3. Polysaccharides Isolated from Ganoderma Species
3.1. Polysaccharides Isolated from G. lucidum
3.2. Polysaccharides Isolated from G. atrum
4. Terpenoids Isolated from Ganoderma Species
4.1. Triterpenoids Isolated from Ganoderma Species
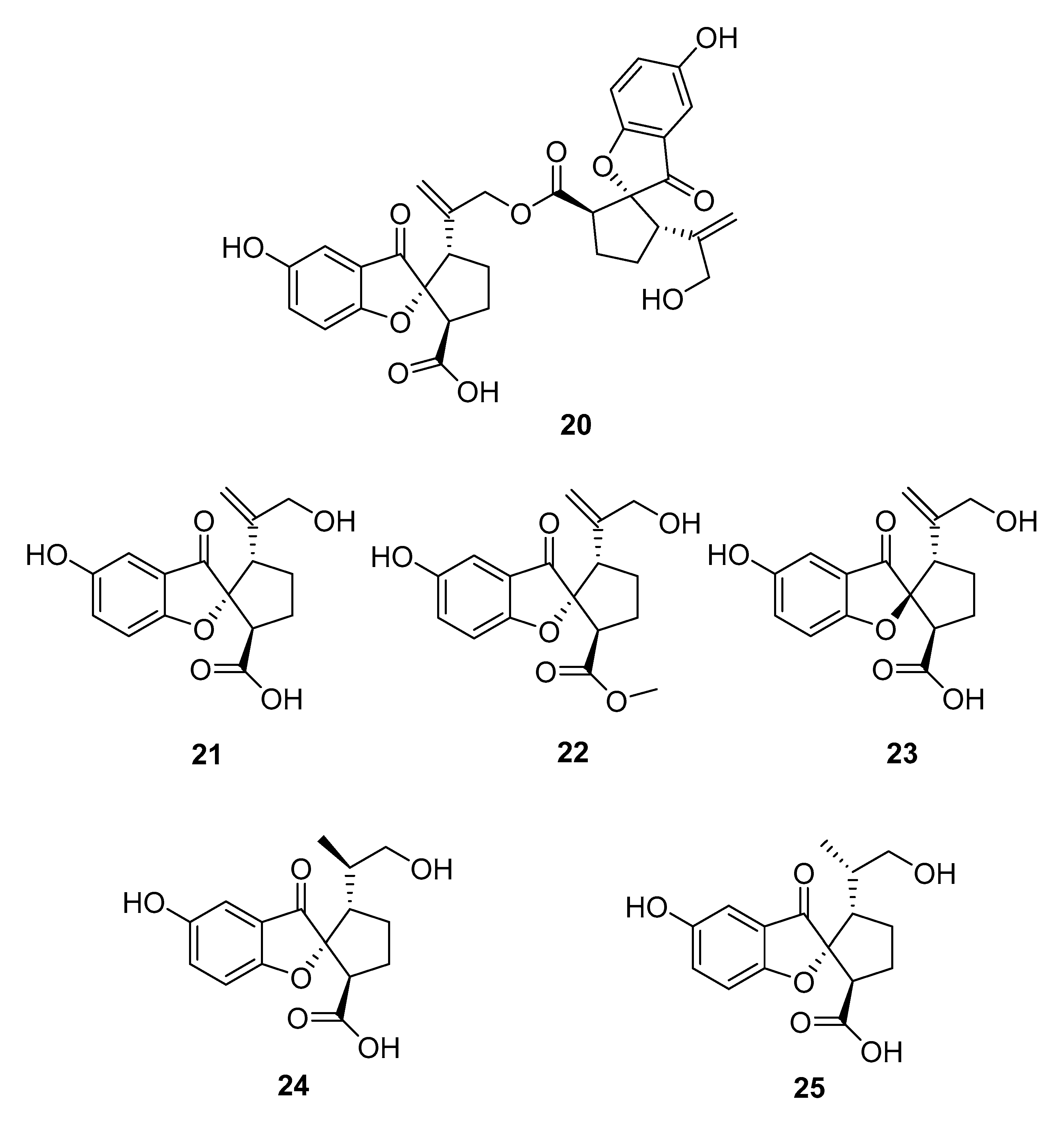
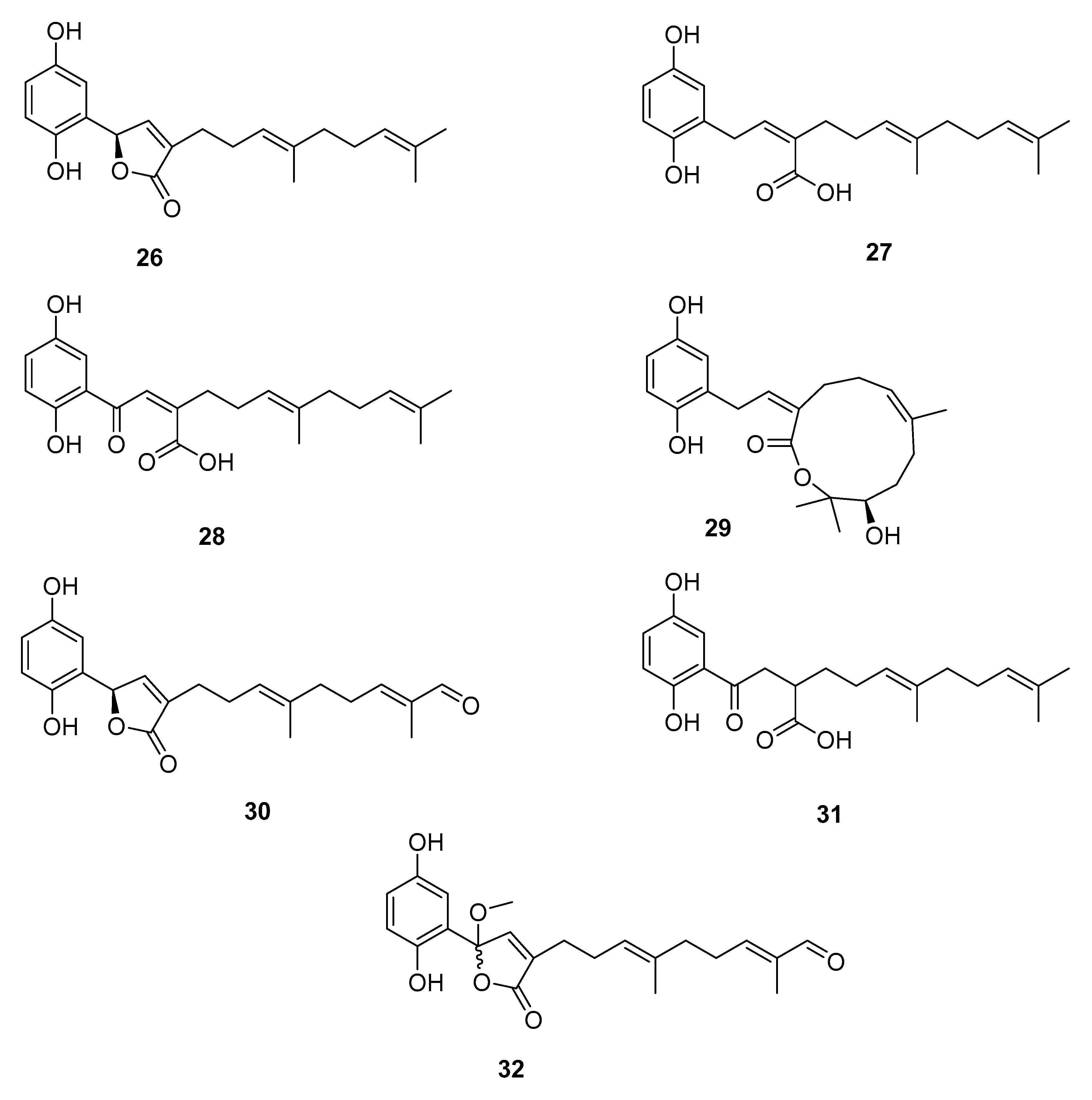
4.2. Meroterpenoids Isolated from Ganoderma Species
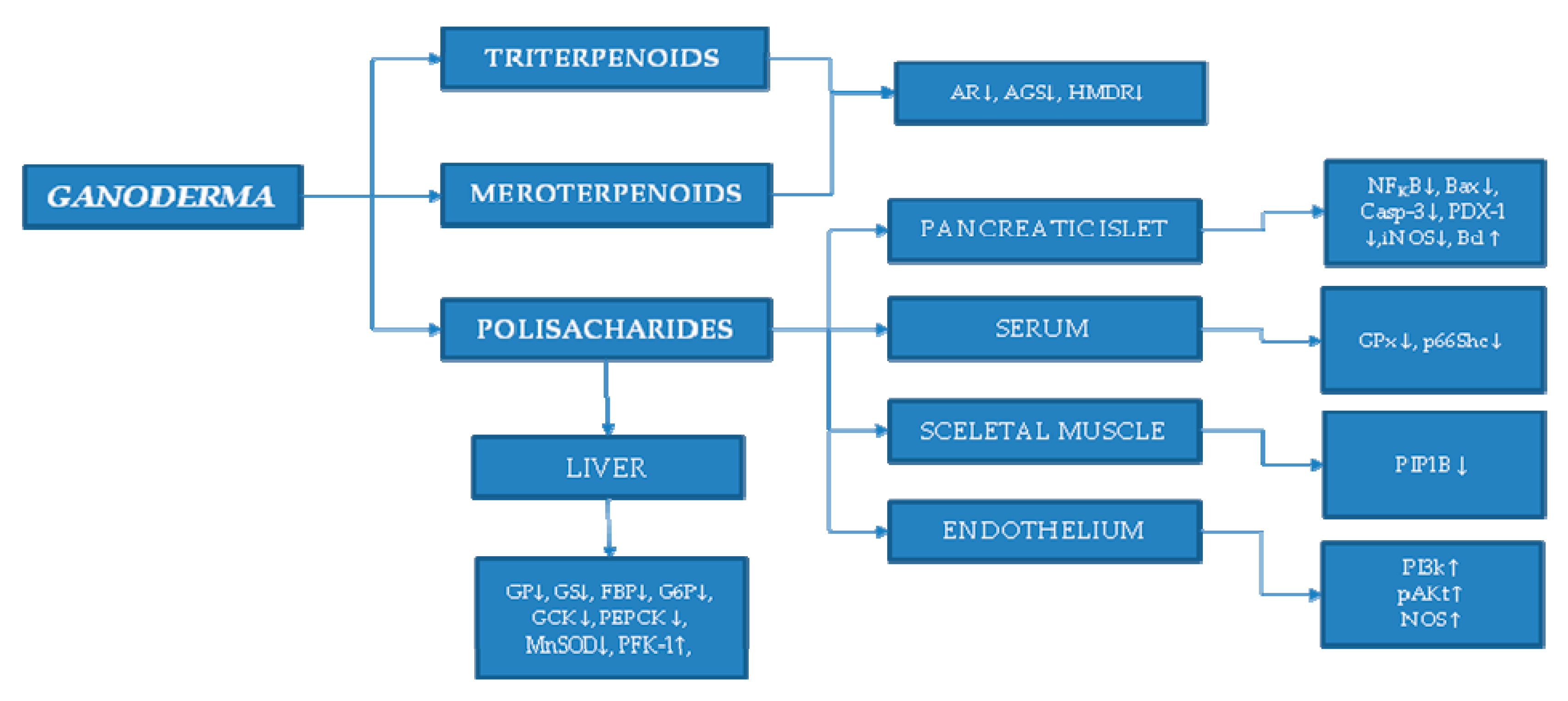
5. Summarized Treatment Mechanism by Ganoderma
6. Future Prospects
- standardization of Ganoderma species sources;
- the relationship between the structure and function of Ganoderma species individual polysaccharides and triterpenoids is not yet fully clear;
- investigation of the possible synergistic or antagonistic effects of active substances with food ingredients;
- developing more efficient and economic approaches for the preparation and modification of the most active compounds;
- polysaccharides structure determination methods are far from reaching the protein and nucleic acid structure determination as automated, micro-quantified, and standardized;
- polysaccharides in vivo mechanism of action is not yet fully clear, especially lacking in full toxicological profile.
Author Contributions
Funding
Conflicts of Interest
References
- Zheng, Y.; Bai, L.; Zhou, Y.; Tong, R.; Zeng, M.; Li, X.; Shi, J. Polysaccharides from Chinese herbal medicine for anti-diabetes recent advances. Int. J. Biol. Macromol. 2019, 121, 1240–1253. [Google Scholar] [CrossRef] [PubMed]
- Ahlqvist, E.; Storm, P.; Käräjämäki, A.; Martinell, M.; Dorkhan, M.; Carlsson, A.; Vikman, P.; Prasad, R.B.; Aly, D.M.; Almgren, P.; et al. Novel subgroups of adult-onset diabetes and their association with outcomes: A data-driven cluster analysis of six variables. Lancet Diabetes Endocrinol. 2018, 6, 361–369. [Google Scholar] [CrossRef]
- Gloyn, A.L.; Drucker, D.J. Precision medicine in the management of type 2 diabetes. Lancet Diabetes Endocrinol. 2018, 6, 891–900. [Google Scholar] [CrossRef]
- Khan, R.M.M.; Chua, Z.J.Y.; Tan, J.C.; Yang, Y.; Liao, Z.; Zhao, Y. From pre-diabetes to diabetes: Diagnosis, treatments and translational research. Medicina 2019, 55, 546. [Google Scholar] [CrossRef] [PubMed]
- Fatmawati, S.; Shimizu, K.; Kondo, R. Structure-activity relationships of ganoderma acids from Ganoderma lucidum as aldose reductase inhibitors. Bioorg. Med. Chem. Lett. 2011, 21, 7295–7297. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, H.E. Alpha-glucosidase inhibitors. Endocrinol. Metab. Clin. N. Am. 1997, 26, 539–551. [Google Scholar] [CrossRef]
- Rena, G.; Hardie, D.G.; Pearson, E.R. The mechanisms of action of metformin. Diabetologia 2017, 60, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Fuchtenbusch, M.; Standl, E.; Schatz, H. Clinical efficacy of new thiazolidinediones and glinides in the treatment of type 2 diabetes mellitus. Exp. Clin. Endocrinol. Diabetes 2000, 108, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Montagnani, A.; Gonnelli, S. Antidiabetic therapy effects on bone metabolism and fracture risk. Diabetes Obes. Metab. 2013, 15, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Egan, A.G.; Blind, E.; Dunder, K.; de Graeff, P.A.; Hummer, T.; Bourcier, T.; Rosebraugh, C. Pancreatic safety of incretin- based drugs – FDA and EMA assessment. N. Engl, J. Med. 2014, 370, 794–797. [Google Scholar] [CrossRef] [PubMed]
- Bistola, V.; Lambadiari, V.; Dimitriadis, G.; Ioannidis, I.; Makrilakis, K.; Tentolouris, N.; Tsapas, A.; Parissis, J. Possible mechanisms of direct cardiovascular impact of GLP-1 agonists and DPP4 inhibitors. Heart Fail. Rev. 2018, 23, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.; Wittstein, K.; Kirk, P.M.; Stadler, M. An assessment of the taxonomy and chemotaxonomy of Ganoderma. Fungal Divers. 2015, 71, 1–15. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Chen, H.; Chen, X.; Lan, J.; Liu, C. Complete mitochondrial genome of the medicinal mushroom Ganoderma lucidum. PLoS ONE 2013, 8, e72038. [Google Scholar] [CrossRef] [PubMed]
- Boh, B. Ganoderma lucidum: A potential for biotechnological production of anti-cancer and immunomodulatory drugs. Rec. Pat. Anti-cancer Drug Discov. 2013, 8, 255–287. [Google Scholar] [CrossRef]
- Paterson, R.R. Ganoderma — a therapeutic fungal biofactory. Phytochemistry 2006, 67, 1985–2001. [Google Scholar] [CrossRef] [PubMed]
- Fatmawati, S.; Shimizu, K.; Kondo, R. Ganoderic acid Df, a new triterpenoid with aldose reductase inhibitory activity from the fruiting body of Ganoderma lucidum. Fitoterapia 2010, 81, 1033–1036. [Google Scholar] [CrossRef] [PubMed]
- Sliva, D. Ganoderma lucidum in cancer research. Leuk. Res. 2006, 30, 767–768. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Qiu, M. Meroterpenoids from Ganoderma species: A review of last five years. Nat. Prod. Bioprosp. 2018, 8, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Shim, S.H.; Kim, J.S.; Shin, K.H.; Kang, S.S. Aldose reductase inhibitors from the fruiting bodies of Ganoderma applanatum. Biol. Pharm. Bull. 2005, 28, 1103–1105. [Google Scholar] [CrossRef] [PubMed]
- Ngai, P.H.K.; Ng, T.B. A mushroom (Ganoderma capense) lectin with spectacular thermostability, potent mitogenic activity on splenocytes, and antiproliferative activity toward tumor cells. Biochem. Biophys. Res. Commun. 2004, 314, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Raman, J.; Malek, S.N.A.; John, P.A.; Vikineswary, S. Green synthesis of silver nanoparticles using Ganoderma neo-japonicum Imazeki: A potential cytotoxic agent against breast cancer cells. Int. J. Nanomed. 2013, 8, 4399–4413. [Google Scholar]
- Seto, S.W.; Lam, T.Y.; Tam, H.L.; Au, A.L.S.; Chan, S.W.; Wu, J.H.; Yu, P.H.F.; Leung, G.P.H.; Ngai, S.M.; Yeung, J.H.K.; et al. Novel hypoglycemic effects of Ganoderma lucidum water-extract in obese/diabetic (+db/+db) mice. Phytomedicine 2009, 16, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Ratnaningtyas, N.I.; Hernayanti, H.; Andarwanti, S.; Ekowati, N.; Purwanti, E.S.; Sukmawati, D. Effects of Ganoderma lucidum extract on diabetic rats. Biosaintifika: J. Biol. Biol. Edu. 2018, 10, 642–647. [Google Scholar] [CrossRef]
- Oluba, O.M.; Onyeneke, E.C.; Ojieh, G.C.; Idonije, B.O.; Ojiezeh, T.I. Hepatoprotective potential of aqueous extract of Ganoderma lucidum against carbon tetrachloride intoxication in rats. Pharm. Lett. 2010, 2, 432–439. [Google Scholar]
- Sarker, M.M.R. Antihyperglycemic, insulin-sensitivity and anti-hyperlipidemic potential of Ganoderma lucidum, a dietary mushroom, onalloxan-and glucocorticoid-induced diabetic Long-Evans rats. Funct. Foods Health Dis. 2015, 5, 450–466. [Google Scholar]
- Bach, E.; Hi, E.; Martins, A.; Nascimento, P.; Wadt, N. Hypoglicemic and hypolipedimic efects of Ganoderma lucidum in streptozotocin-induced diabetic rats. Medicines 2018, 5, 78. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F. Ganoderma lucidum: Persuasive biologically active constituents and their health endorsement. Biomed. Pharmacother 2018, 107, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Sirisidthi, K.; Kosai, P.; Jiraungkoorskul, W. Antidiabetic activity of the lingzhi or reishi medicinal mushroom Ganoderma lucidum: A review. S. Afr. Pharm. J. 2016, 83, 45–47. [Google Scholar]
- Tie, L.; Yang, H.Q.; An, Y.; Liu, S.Q.; Han, J.; Xu, Y.; Hu, M.; Li, W.D.; Chen, A.F.; Lin, Z.B.; et al. Ganoderma lucidum polysaccharide accelerates refractory wound healing by inhibition of mitochondrial oxidative stress in type 1 diabetes. Cell. Physiol. Biochem. 2012, 29, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-C.; Zhao, S.; Yang, B.-Y.; Wang, Q.-H.; Kuang, H.-X. Anti-diabetic polysaccharides from natural sources: A review. Carbohydr. Polym. 2016, 148, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Oliver-Krasinski, J.M.; Kasner, M.T.; Yang, J.; Crutchlow, M.F.; Rustgi, A.K.; Kaestner, K.H.; Stoffers, D.A. The diabetes gene Pdx1 regulates the transcriptional network of pancreatic endocrine progenitor cells in mice. J. Clin. Investig. 2009, 119, 1888–1898. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.-T.; Hsieh, J.-F.; Chen, S.-T. Anti-diabetic effects of Ganoderma lucidum. Phytochemistry 2015, 114, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Wu, Q.; Zhang, J.; Xie, Y.; Cai, W.; Tan, J. Antidiabetic activity of Ganoderma lucidum polysaccharides F31 down-regulated hepatic glucose regulatory enzymes in diabetic mice. J. Ethnopharmacol. 2017, 196, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Wu, Q.; Xie, Y.; Tan, J.; Ding, Y.; Bai, L. Hypoglycemic mechanisms of Ganoderma lucidum polysaccharides F31 in db/db mice via RNA-seq and iTRAQ. Food Funct. 2018, 9, 6495–6507. [Google Scholar] [CrossRef] [PubMed]
- Hikino, H.; Ishiyama, M.; Suzuki, Y.; Konno, C. Mechanisms of hypoglycemic activity of ganoderan B: A glycan of Ganoderma lucidum fruit body. Planta Med. 1989, 55, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Teng, B.-S.; Wang, C.-D.; Zhang, D.; Wu, J.; Pan, D.; Pan, L.; Yang, H.; Zhou, P. Hypoglycemic effect and mechanism of a proteoglycan from Ganoderma lucidum on streptozotocin-induced type 2 diabetic rats. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 166–175. [Google Scholar] [PubMed]
- Zhu, K.; Nie, S.; Li, C.; Lin, S.; Xing, M.; Li, W. A newly identified polysaccharide from Ganoderma atrum attenuates hyperglycemia and hyperlipidemia. Int. J. Biol. Macromol. 2013, 57, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.X.; Nie, S.P.; Li, C.; Gong, D.; Xie, M.Y. Ganoderma atrum polysaccharide improves aortic relaxation in diabetic rats via PI3K/Akt pathway. Carbohydr. Polym. 2014, 103, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Skalicka-Woźniak, K.; Szypowski, J.; Łoś, R.; Siwulski, M.; Sobieralski, K.; Głowniak, K.; Malm, A. Evaluation of polysaccharides content in fruit bodies and their antimicrobial activity of four Ganoderma lucidum (W Curt.: Fr.) P. Karst. strains cultivated on different wood type substrates. Acta Soc. Bot. Pol. 2012, 81, 17–21. [Google Scholar] [CrossRef]
- Hwang, I.-W.; Kim, B.-M.; Kim, Y.-C.; Lee, S.-H.; Chung, S.-K. Improvement in β-glucan extraction from Ganoderma lucidum with high-pressure steaming and enzymatic pre-treatment. Appl. Biol. Chem. 2018, 61, 235–242. [Google Scholar] [CrossRef]
- Zhang, H.N.; He, J.H.; Yuan, L.; Lin, Z.B. In vitro and in vivo protective effect of Ganoderma lucidum polysaccharides on alloxan-induced pancreaticis lets damage. Life Sci. 2003, 73, 2307–2319. [Google Scholar] [CrossRef]
- Zheng, J.; Yang, B.; Yu, Y.; Chen, Q.; Huang, T.; Li, D. Ganoderma lucidum polysaccharides exert anti-hyperglycemic effect on streptozotocin-induced diabetic rats through affecting β-cells. Comb. Chem. High Throughput Screen 2012, 15, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.N.; Lin, Z.B. Hypoglycemic effect of Ganoderma lucidum polysaccharides. Acta Pharmacol. Sin. 2004, 25, 191–195. [Google Scholar] [PubMed]
- Xiao, C.; Wu, Q.P.; Cai, W.; Tan, J.B.; Yang, X.B.; Zhang, J.M. Hypoglycemic effects of Ganoderma lucidum polysaccharides in type 2 diabetic mice. Arch. Pharmacal Res. 2012, 35, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Hikino, H.; Konno, C.; Mirin, Y.; Hayashi, T. Isolation and hypoglycemic activity of ganoderans A and B, glycans of Ganoderma lucidum fruit bodies. Planta Med. 1985, 51, 339–340. [Google Scholar] [CrossRef] [PubMed]
- Tomoda, M.; Gonda, R.; Kasahara, Y.; Hikino, H. Glycan structures of ganoderans B and C, hypoglycemic glycans of Ganoderma lucidum fruit bodies. Phytochemistry 1986, 25, 2817–2820. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Zhang, W.; Sun, M.; Zhang, Z. Hypoglycemic effect of inulin combined with Ganoderma lucidum polysaccharides in T2DM rats. J. Funct. Foods 2019, 55, 381–390. [Google Scholar] [CrossRef]
- Gao, Y.; Lan, J.; Dai, X.; Ye, J.; Zhou, S. A phase I/II study of Ling Zhi mushroom Ganoderma lucidum (W. Curt.: Fr.) Lloyd (Aphyllophoromycetideae) extract in patients with type II diabetes mellitus. Int. J. Med. Mushrooms 2004, 6, 3–9. [Google Scholar]
- Zhang, H.; Li, W.-J.; Nie, S.-P.; Chen, Y.; Wang, Y.-X.; Xie, M.-Y. Structural characterisation of a novel bioactive polysaccharide from Ganoderma atrum. Carbohydr. Polym. 2012, 88, 1047–1054. [Google Scholar] [CrossRef]
- Perveen, S.; Al-Taweel, A. Introductory chapter: Terpenes and terpenoids. IntechOpen 2018. [Google Scholar] [CrossRef]
- Fatmawati, S.; Kurashiki, K.; Takeno, S.; Kim, Y.; Shimizu, K.; Sato, M.; Imaizumi, K.; Takahashi, K.; Kamiya, S.; Kaneko, S.; et al. The Inhibitory Effect on Aldose Reductase by an Extract of Ganoderma lucidum. Phytother. Res. 2009, 23, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Fatmawati, S.; Shimizua, K.; Kondoa, R. Ganoderol B: A potent -glucosidase inhibitor isolated from the fruiting body of Ganoderma lucidum. Phytomedicine 2011, 18, 1053–1055. [Google Scholar] [CrossRef] [PubMed]
- Fatmawati, S.; Kondo, R.; Shimizu, K. Structure-activity relationships of lanostane-type triterpenoids from Ganoderma lingzhi as α-glucosidase inhibitors. Bioorg. Med. Chem. Lett. 2013, 23, 5900–5903. [Google Scholar] [CrossRef] [PubMed]
- Satria, D.; Tamrakar, S.; Suhara, H.; Kaneko, S.; Shimizu, K. Mass spectrometry-based untargeted metabolomics and α-glucosidase inhibitory activity of Lingzhi (Ganoderma lingzhi) during the developmental stages. Molecules 2019, 24, 2044. [Google Scholar] [CrossRef] [PubMed]
- Satria, D.; Amena, Y.; Niwa, Y.; Ashoura, A.; Allama, A.E.; Shimizua, K. Lucidumol D, a new lanostane-type triterpene from fruiting bodies of Reishi (Ganoderma lingzhi). Nat. Prod. Res. 2019, 33, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Bao, L.; Xiong, W.; Ma, K.; Han, J.; Wang, W.; Yin, W.; Liu, H. Lanostane triterpenes from the Tibetan medicinal mushroom Ganoderma leucocontextum and their inhibitory effects on HMG-CoA reductase and α-glucosidase. J. Nat. Prod. 2015, 78, 1977–1989. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ma, K.; Chen, H.; Wang, K.; Xiong, W.; Bao, L.; Liu, H. A novel polycyclic meroterpenoid with aldose reductase inhibitory activity from medicinal mushroom Ganoderma leucocontextum. J. Antibiot. 2017, 70, 915–917. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Bao, L.; Ma, K.; Zhang, J.; Chen, B.; Han, J.; Ren, J.; Luo, H.; Liu, H. A novel class of α-glucosidase and HMG-CoA reductase inhibitors from Ganoderma leucocontextum and the anti-diabetic properties of ganomycin I in KK-Ay mice. Eur. J. Med. Chem. 2017, 127, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Mittal, A.; Dabur, R. Mechanistic approach of anti-diabetic compounds identified from natural sources. Chem. Biol. Lett. 2018, 5, 63–99. [Google Scholar]
- Bhatnagar, A.; Srivastava, S.K. Aldose reductase: Congenial and injurious profiles of an enigmatic enzyme. Biochem. Med. Metab. Biol. 1992, 48, 91–121. [Google Scholar] [CrossRef]
- Schemmel, K.E.; Padiyara, R.S.; D’Souza, J.J. Aldose reductase inhibitors in the treatment of diabetic peripheral neuropathy: A review. J. Diabetes Compl. 2010, 24, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Anti-diabetic effects and mechanisms of dietary polysaccharides. Molecules 2019, 24, 2556. [Google Scholar] [CrossRef] [PubMed]
- Franz, M.J.; Boucher, J.L.; Rutten-Ramos, S.; VanWormer, J.J. Lifestyle weight-loss intervention outcomes in overweight and obese adults with type 2 diabetes: A systematic review and meta-analysis of randomized clinical trials. J. Acad. Nutr. Diet. 2015, 115, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.-D.; Cheng, K.-C. From nutraceutical to clinical trial: Frontiers in Ganoderma development. Appl. Microbiol. Biotechnol. 2018, 102, 9037–9051. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.H.; Zhou, S.F.; Jiang, W.Q.; Huang, M.; Dai, X.H. Effects of Ganopoly (R) (a Ganoderma lucidum polysaccharide extract) on the immune functions in advanced-stage cancer patients. Immunol. Investig. 2003, 32, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Klupp, N.L.; Kiat, H.; Bensoussan, A.; Steiner, G.Z.; Chang, D.H. A double-blind, randomised, placebo-controlled trial of Ganoderma lucidum for the treatment of cardiovascular risk factors of metabolic syndrome. Sci. Rep. 2016, 6, 29540. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.S.; Kao, C.H.; Xu, Y.; Glucina, M.P.; Paterson, R.R.M.; Ferguson, L.R. From 2000 years of Ganoderma lucidum to recent developments in nutraceuticals. Phytochemistry 2015, 114, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Current findings, future trends, and unsolved problems in studies of medicinal mushrooms. Appl. Microbiol. Biotechnol. 2011, 89, 1323–1332. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Extract | Experimental Model | Dose | Concentration of Glucose | Reference |
|---|---|---|---|---|---|
| G. lucidum | Water extract of powder of the capsules of G. lucidum contains 95% Ganoderma extract (obtained from the whole fruit body) and 5% dextrin. | C57BL/KsJ mice (female; 6 months old) (normal mice) | 0 g/kg | 168.5 mg/dL | [22] |
| 0.003 g/kg | 161.6 mg/dL | ||||
| 0.03 g/kg | 126.5 mg/dL | ||||
| 0.3 g/kg | 68.5 mg/dL | ||||
| C57BL/KsJ mice (female; 6 months old) (diabetic mice) | 0 g/kg | 668.5 mg/dL | |||
| 0.003 g/kg | 645.9 mg/dL | ||||
| 0.03 g/kg | 441.5 mg/dL | ||||
| 0.3 g/kg | 288,4 mg/dL | ||||
| G. lucidum | Ethanol extract of powdered fruiting bodies | white rats (Rattus norvegicus) derived from Wistar line, male, 2–3 weeks old | 0 mg/kg | 435.75 mg/dL | [23] |
| 250 mg/kg | 312.00 mg/dL | ||||
| 0 mg/kg | 311.00 mg/dL | ||||
| 500 mg/kg | 203.50 mg/dL | ||||
| 0 mg/kg | 384.25 mg/dL | ||||
| 1000 mg/kg | 140.50 mg/dL | ||||
| G. lucidum | Water extract of fruiting bodies | male Wistar rats weighing 85 – 100 g, normal rats | 0 mg/kg | 90 mg/dL | [24] |
| 100 mg/kg | 60 mg/dL | ||||
| 200 mg/kg | 45 mg/dL | ||||
| male Wistar rats weighing 85 – 100 g, diabetic rats | 0 mg/kg | 200 mg/dL | |||
| 100 mg/kg | 150 mg/dL | ||||
| 200 mg/kg | 90 mg/dL | ||||
| G. lucidum | Methanol extract of fruiting bodies | Long-Evans male rats (4 weeks age, weight 83.5 g), induced with alloxan | 0 mg/kg | 14.5 mg/dL | [25] |
| 200 mg/kg | 12.5 mg/dL | ||||
| 400 mg/kg | 12 mg/dL | ||||
| 600 mg/kg | 10 mg/dL | ||||
| 800 mg/kg | 10 mg/dL | ||||
| Petroleum ether extract of fruiting bodies | 0 mg/kg | 14.5 mg/dL | |||
| 200 mg/kg | 10 mg/dL | ||||
| 400 mg/kg | 9.5 mg/dL | ||||
| 600 mg/kg | 7 mg/dL | ||||
| 800 mg/kg | 7 mg/dL | ||||
| Methanol extract of fruiting bodies | Long-Evans male rats (4 weeks age, weight 83.5 g), induced with corticosteroid | 0 mg/kg | 17.5 mg/dL | ||
| 200 mg/kg | 14 mg/dL | ||||
| 400 mg/kg | 12.5 mg/dL | ||||
| 600 mg/kg | 10 mg/dL | ||||
| 800 mg/kg | 9.5 mg/dL | ||||
| Petroleum ether extract of fruiting bodies | 0 mg/kg | 17.5 mg/dL | |||
| 200 mg/kg | 10 mg/dL | ||||
| 400 mg/kg | 7.5 mg/dL | ||||
| 600 mg/kg | 5 mg/dL | ||||
| 800 mg/kg | 4.8 mg/dL | ||||
| G. lucidum | Water extract of fruiting bodies (80%) and alcohol extract of fruiting bodies (20%) | Male Wistar rats aged four weeks and weighing 250 to 300 g | 0 mL/kg | 456 mg/dL | [26] |
| 1 mL/kg | 265 mg/dL |
| Polysaccharide | Tissue | Enzyme | Reference |
|---|---|---|---|
| Hot water extract | liver | glycogen phosphorylase (GP) ↓ | [27] |
| fructose-1,6-bisphosphatase (FBP) ↓ | |||
| glucose-6-phosphatase (G6P) ↓ | |||
| phosphoenolpyruvate carboxykinase (PEPCK) ↓ | [27,28] | ||
| glycogen synthase (GS) ↓ | [28] | ||
| manganese superoxide dismutase (MnSOD) ↓ | [29] | ||
| serum | glutathione peroxidase (GPx) ↓ | ||
| p66Shc ↓ | |||
| pancreatic islet | nuclear factor kappa (NF-κB) ↓ | [30] | |
| insulin promoter factor 1 (PDX-1) ↓ | [31,32] | ||
| Bax ↓ | |||
| calcium-insensitive nitric oxide synthase (iNOS) ↓ | |||
| caspase 3 (Casp-3) ↓ | |||
| F31 | liver | AMP-activated protein kinase (AMPK) ↓ | [1,33,34] |
| ganoderan B | liver | glucokinase (GCK) ↑ | [35] |
| Phosphofructokinase (PFK-1) ↑ | |||
| glucose-6-phosphate dehydrogenase (G6PDH) ↑ | |||
| glucose-6-phosphatase (G6P) ↓ | |||
| glycogen synthase (GS) ↓ | |||
| FYGL | skeletal muscle | tyrosine phosphatase 1B (PTP1B) ↓ | [36] |
| PSG-1 (G. atrum) | pancreatic cells | Bax ↓ | [37] |
| Bcl-2 ↑ | |||
| endothelium | phosphoinositide 3-kinase (PI3K) ↑ | [38] | |
| phospho-Akt (p-Akt) ↑ | |||
| nitric oxide synthase (eNOS) ↑ |
| Species | Terpenoids | Enzyme | Reference |
|---|---|---|---|
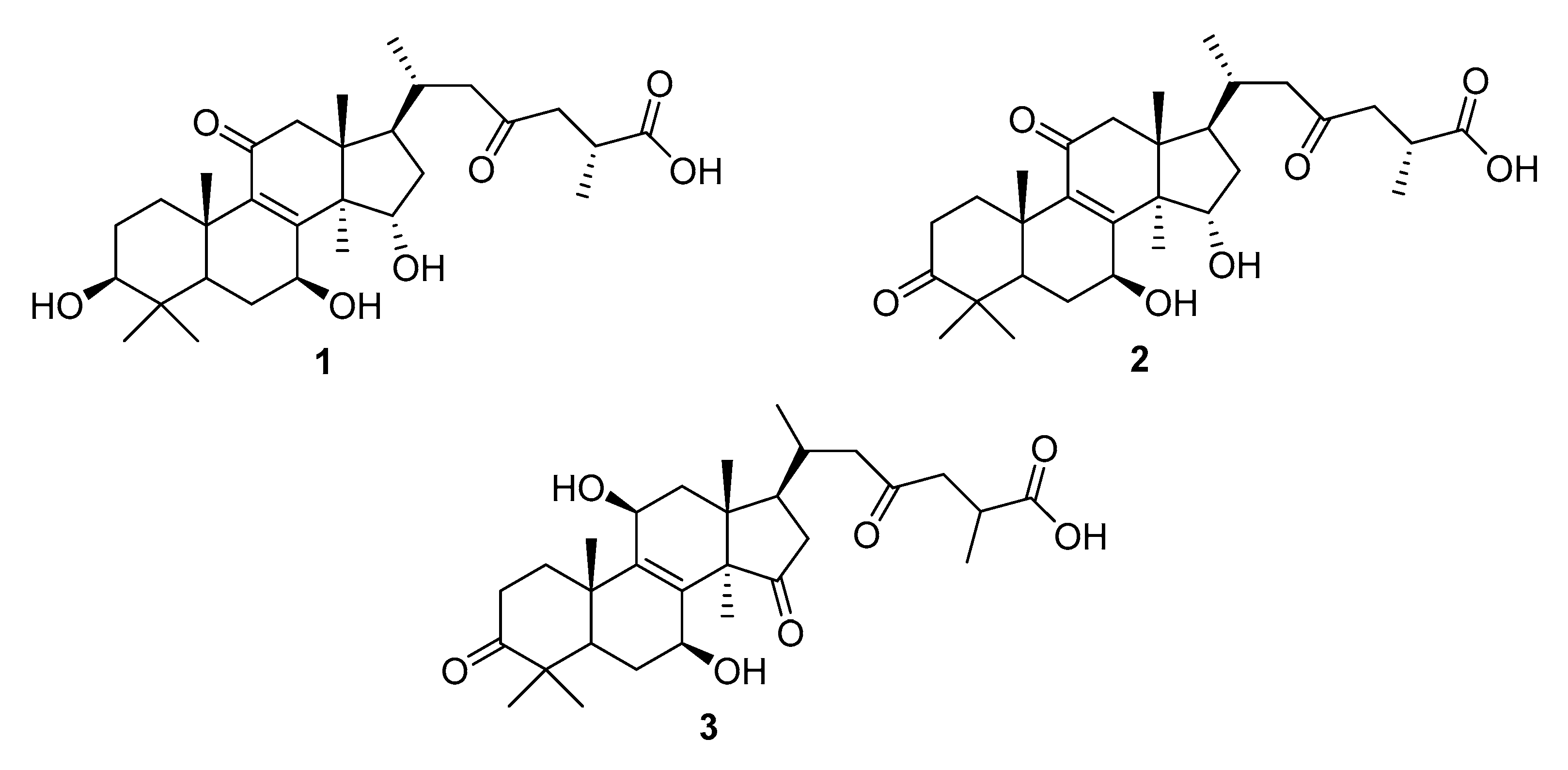
| G. lucidum | ganoderic acid C2 (1), ganoderenic acid A (2) ganoderic acid Df (3) | aldose reductase (AR) ↓ | [3,16,32,51] |
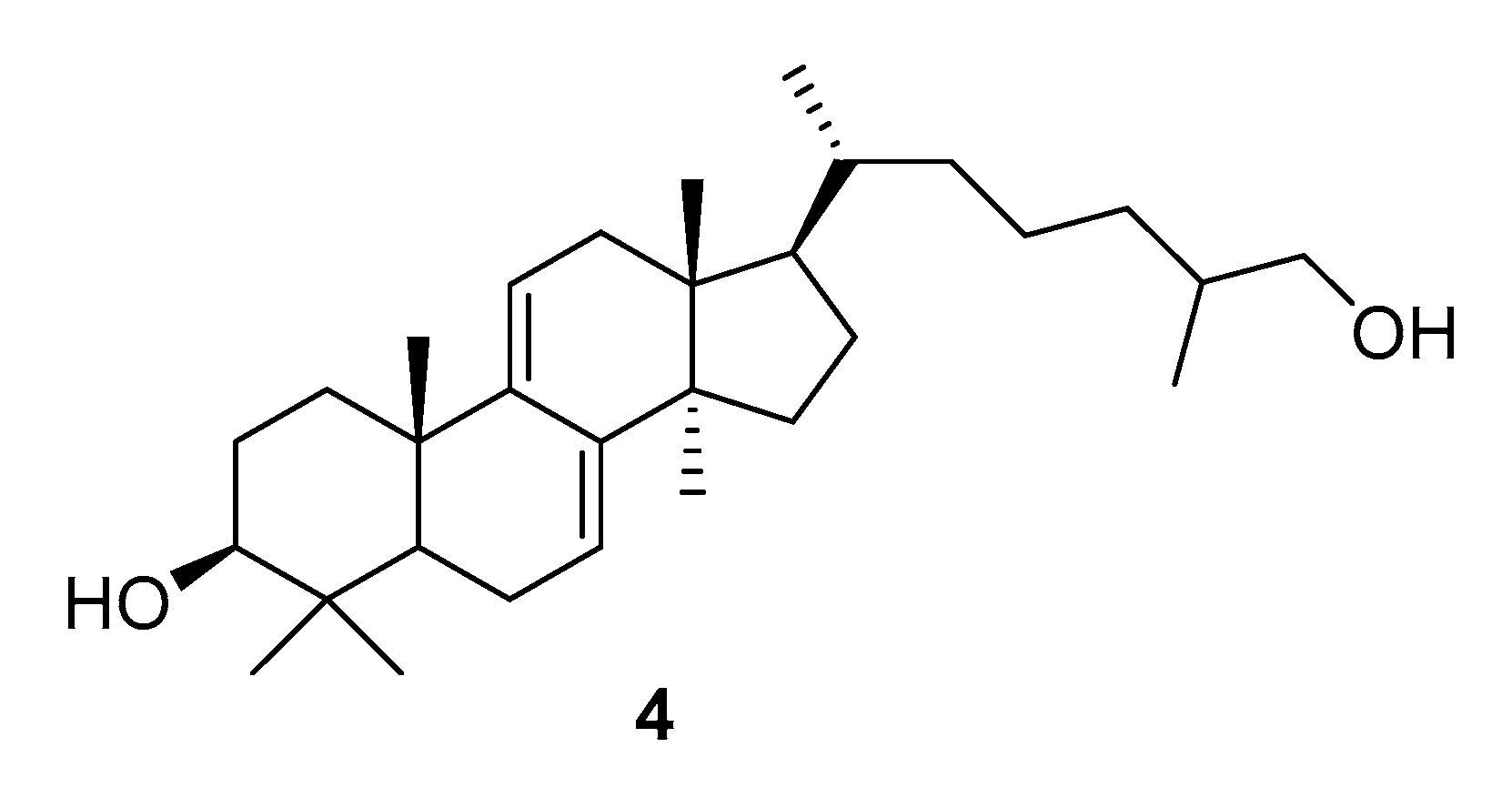
| G. lucidum | ganoderol B (4) | α-glucosidase (AGS) ↓ | [52,53] |
| G. lingzhi | 3,12,20-trihydroxy-7,11,15-trioxolanost-8,16,24- -trien-26-oic acid,ethanolic extract of G. lingzhi | [54,55] | |
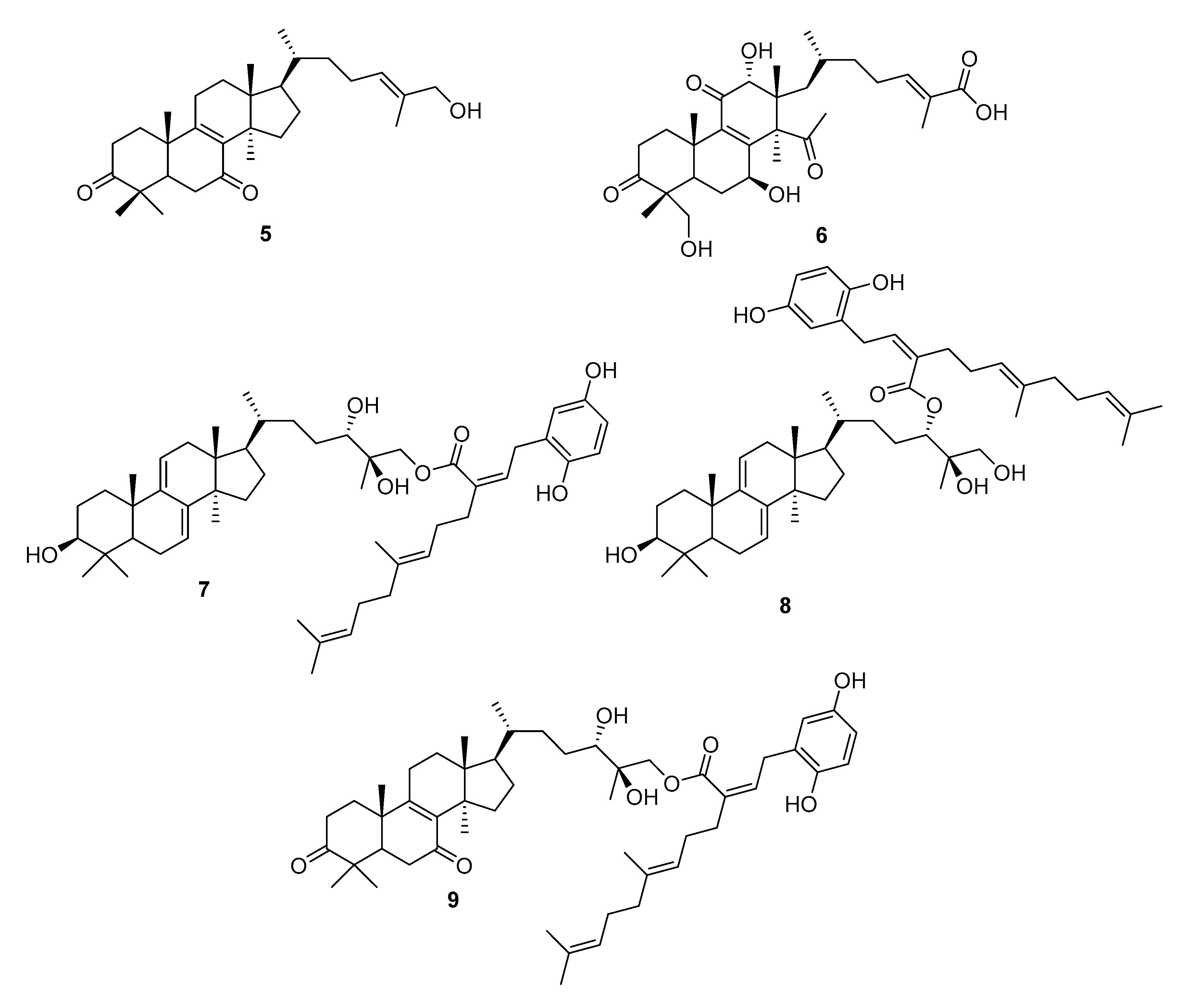
| G. leucocontexum | ganoderone A (5), ganoleuconin E, M, N, P, (6–9) | [56] | |
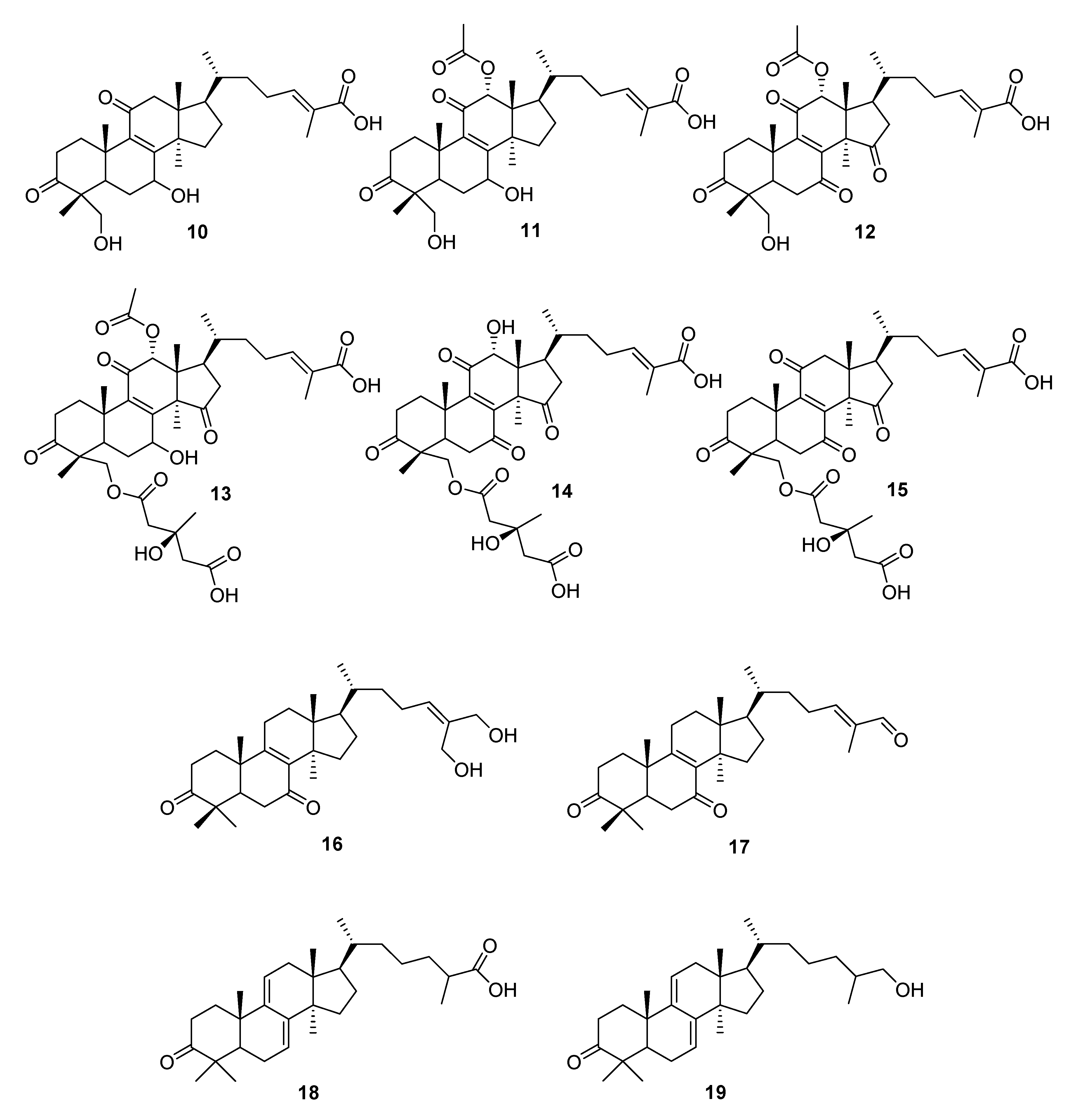
| G. leucocontexum | ganoleuconin A (10), C (11), F (12), J (13), K (14),L (15), M (7), N (8), ganoderiol J (16) | HMG-CoA reductase (HMGR) ↓ | [56] |
| G. leucocontextum | ganoleucin D (20), spiroapplanatumine L (22) (±)-spirolingzhine A (23, 24) spirolingzhine D (25) | aldose reductase ↓ | [57] |
| ganoleucin D (20) | HMG-CoA reductase ↓ | ||
| ganomycin I (26) | [58] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wińska, K.; Mączka, W.; Gabryelska, K.; Grabarczyk, M. Mushrooms of the Genus Ganoderma Used to Treat Diabetes and Insulin Resistance. Molecules 2019, 24, 4075. https://doi.org/10.3390/molecules24224075
Wińska K, Mączka W, Gabryelska K, Grabarczyk M. Mushrooms of the Genus Ganoderma Used to Treat Diabetes and Insulin Resistance. Molecules. 2019; 24(22):4075. https://doi.org/10.3390/molecules24224075
Chicago/Turabian StyleWińska, Katarzyna, Wanda Mączka, Klaudia Gabryelska, and Małgorzata Grabarczyk. 2019. "Mushrooms of the Genus Ganoderma Used to Treat Diabetes and Insulin Resistance" Molecules 24, no. 22: 4075. https://doi.org/10.3390/molecules24224075
APA StyleWińska, K., Mączka, W., Gabryelska, K., & Grabarczyk, M. (2019). Mushrooms of the Genus Ganoderma Used to Treat Diabetes and Insulin Resistance. Molecules, 24(22), 4075. https://doi.org/10.3390/molecules24224075