
Red Beetroot and Betalains as Cancer Chemopreventative Agents


Abstract
:1. Introduction
2. Bioactive Compounds in Beetroot
2.1. Betaine
2.2. Polyphenols
2.2.1. Phenolic Acids
2.2.2. Flavonoids
2.2.3. The Triterpene Saponins
3. Carotenoids
4. Non-Defined Red Beetroot Components with Chemopreventive Activity
5. Betanins
5.1. Betanins Bioavailability
5.2. Anti-Oxidant and Anti-Inflammatory Activity
5.3. Phase II Detoxifying Enzyme Activity
5.4. Betanins Cytotoxicity of Cultured Cells
5.5. Chemoprevention with Betanins
6. Conclusions and Future Reaearch
Funding
Conflicts of Interest
References
- Wang, L.S.; Kuo, C.T.; Huang, Y.W.; Stoner, G.D.; Lechner, J.F. Gene-Diet Interactions on Colorectal Cancer Risk. Curr. Nutr. Rep. 2012, 1, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, Inflammation, and Cancer: How are they Linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- Meitzler, J.; Antony, S.; Wu, Y.; Juhasz, A.; Liu, H.; Jiang, G.; Lu, J.; Roy, K.; Doroshow, J.H. NADPH Oxidases: A Perspective on Reactive Oxygen Species Production in Tumor Biology. Antioxidants Redox. Signal. 2014, 20, 2873–2889. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhou, Z.; Min, W. Mitochondria, Oxidative Stress and Innate Immunity. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Vallée, A.; Lecarpentier, Y. Crosstalk Between Peroxisome Proliferator-Activated Receptor Gamma and the Canonical WNT/β-Catenin Pathway in Chronic Inflammation and Oxidative Stress During Carcinogenesis. Front. Immunol. 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Truong, T.H.; Carroll, K.S. Redox Regulation of Protein Kinases. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 332–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reczek, C.R.; Chandel, N.S. ROS-dependent Signal Transduction. Curr. Opin. Cell Biol. 2015, 33, 8–13. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell. Long. 2016, 18. [Google Scholar] [CrossRef]
- Hofman, P.M. Pathobiology of the Neutrophil-intestinal Epithelial Cell Interaction: Role in Carcinogenesis. World J. Gastroenterol. 2010, 16, 5790–5800. [Google Scholar] [CrossRef]
- Manda, G.; Isvoranu, G.; Comanescu, M.V.; Manea, A.; Debelec Butuner, B.; Korkmaz, K.S. The Redox Biology Network in Cancer Pathophysiology and Therapeutics. Redox. Biol. 2015, 5, 347–357. [Google Scholar] [CrossRef]
- Casey, S.C.; Vaccari, M.; Al-Mulla, F.; Al-Temaimi, R.; Amedei, A.; Barcellos-Hoff, M.H.; Brown, D.G.; Chapellier, M.; Christopher, J.; Curran, C.S.; et al. The Effect of Environmental Chemicals on the Tumor Microenvironment. Carcinogenesis 2015, 36, S160–S183. [Google Scholar] [CrossRef] [PubMed]
- Munn, L.L. Cancer and Inflammation. Wiley Interdiscip. Rev. Syst. Biol. Med. 2017, 9. [Google Scholar] [CrossRef]
- Malzev, V.N. Hallmarks of Cancer: Interpretation of Carcinogenic Hypercycle. J. Carcinog. Mutagene. 2016, 7. [Google Scholar] [CrossRef]
- Loo, G. Redox-sensitive Mechanisms of Phytochemical-mediated Inhibition of Cancer Cell Proliferation (review). J. Nutr. Biochem. 2003, 14, 64–73. [Google Scholar] [CrossRef]
- Pandey, M.K.; Gupta, S.C.; Nabavizadeh, A.; Aggarwal, B.B. Regulation of Cell Signaling Pathways by Dietary Agents for Cancer Prevention and Treatment. Semin Cancer Biol. 2017, 46, 158–181. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.K. Beta vulgaris. In Edible Medicinal and Non-Medicinal Plants; Lim, T.K., Ed.; Springer: Dordrecht, The Netherlands, 2016; Volume 10, pp. 26–68. [Google Scholar]
- Chhikara, N.; Kushwaha, K.; Sharma, P.; Gat, Y.; Panghal, A. Bioactive Compounds of Beetroot and Utilization in Food Processing Industry: A Critical Review. Food Chem. 2019, 272, 192–200. [Google Scholar] [CrossRef]
- Zhao, G.; He, F.; Wu, C.; Li, P.; Li, N.; Deng, J.; Zhu, G.; Ren, W.; Peng, Y. Betaine in Inflammation: Mechanistic Aspects and Applications. Front. Immunol. 2018, 24. [Google Scholar] [CrossRef]
- Slow, S.; Elmslie, J.; Lever, M. Dietary Betaine and Inflammation. Am. J. Clin. Nutr. 2008, 88, 247–248. [Google Scholar] [CrossRef]
- Detopoulou, P.; Panagiotakos, D.B.; Antonopoulou, S.; Pitsavos, C.; Stefanadis, C. Dietary Choline and Betaine Intakes in Relation to Concentrations of Inflammatory Markers in Healthy Adults: The ATTICA Study. Am. J. Clin. Nutr. 2008, 87, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, M.; Sudheer, A.R.; Menon, V.P. Ferulic Acid: Therapeutic Potential Through its Antioxidant Property. J. Clin. Biochem. Nutr. 2007, 40, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Perez-Ternero, C.; Werner, C.M.; Nickel, A.G.; Herrera, M.D.; Motilva, M.J.; Böhm, M.; Alvarez de Sotomayor, M.; Laufs, U. Ferulic Acid, a Bioactive Component of Rice Bran, Improves Oxidative Stress and Mitochondrial Biogenesis and Dynamics in Mice and in Human Mononuclear Cells. J. Nutr. Biochem. 2017, 48, 51–61. [Google Scholar] [CrossRef]
- Damasceno, S.S.; Dantas, B.B.; Ribeiro-Filho, J.; de Araújo, D.A.; da Costa, G.M.J. Chemical Properties of Caffeic and Ferulic Acids in Biological System: Implications in Cancer Therapy. A Review. Curr. Pharm. Des. 2017, 23, 3015–3023. [Google Scholar] [CrossRef]
- Wang, S.J.; Zeng, J.; Yang, B.K.; Zhong, Y.M. Bioavailability of Caffeic Acid in Rats and its Absorption Properties in the Caco-2 Cell Model. Pharm. Biol. 2014, 52, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Prasad, N.R.; Jeyanthimala, K.; Ramachandran, S. Caffeic Acid Modulates Ultraviolet Radiation-B Induced Oxidative Damage in Human Blood Lymphocytes. J. Photochem. Photobiol. B 2009, 95, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.Q.; Khan, R.; Qamar, W.; Lateef, A.; Ali, F.; Tahir, M.; Muneeb-U-Rehman; Sultana, S. Caffeic Acid Attenuates 12-O-tetradecanoyl-phorbol-13-acetate (TPA)-induced NF-κB and COX-2 Expression in Mouse Skin: Abrogation of Oxidative Stress, Inflammatory Responses and Proinflammatory Cytokine Production. Food Chem. Toxicol. 2012, 50, 175–183. [Google Scholar] [CrossRef]
- Sharma, S.H.; Rajamanickam, V.; Nagarajan, S. Antiproliferative Effect of p-Coumaric Acid Targets UPR Activation by Downregulating Grp78 in Colon Cancer. Chem. Biol. Interact. 2018, 291, 16–28. [Google Scholar] [CrossRef]
- Sharma, S.H.; Chellappan, D.R.; Chinnaswamy, P.; Nagarajan, S. Protective Effect of p-Coumaric Acid Against 1,2 Dimethylhydrazine Induced Colonic Preneoplastic Lesions in Experimental Rats. Biomed. Pharmacother. 2017, 94, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.H.; Rajamanickam, V.; Nagarajan, S. Supplementation of p-Coumaric Acid Exhibits Chemopreventive Effect via Induction of Nrf2 in a Short-term Preclinical Model of Colon Cancer. Eur. J. Cancer Prev. 2018. [Google Scholar] [CrossRef]
- Bhavani, P.; Subramanian, P.; Kanimozhi, S. Preventive Efficacy of Vanillic Acid on Regulation of Redox Homeostasis, Matrix Metalloproteinases and Cyclin D1 in Rats Bearing Endometrial Carcinoma. Indian J. Clin. Biochem. 2017, 32, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Lamprecht, S.A.; Liu, Y.; Shinozaki, H.; Fan, K.; Leung, D.; Newmark, H.; Steele, V.E.; Kelloff, G.J.; Lipkin, M. Chemoprevention Studies of the Flavonoids Quercetin and Rutin in Normal and Azoxymethane-treated Mouse Colon. Carcinogenesis 2000, 21, 1655–1660. [Google Scholar] [CrossRef] [PubMed]
- Nafees, S.; Rashid, S.; Ali, N.; Hasan, S.K.; Sultana, S. Rutin Ameliorates Cyclophosphamide Induced Oxidative Stress and Inflammation in Wistar Rats: Role of NF-κB/MAPK Pathway. Chem. Biol. Interact. 2015, 231, 98–107. [Google Scholar] [CrossRef]
- Chen, A.Y.; Chen, Y.C. A Review of the Dietary Flavonoid, Kaempferol on Human Health and Cancer Chemoprevention. Food Chem. 2013, 138, 2099–2107. [Google Scholar] [CrossRef]
- Wu, P.; Meng, X.; Zheng, H.; Zeng, Q.; Chen, T.; Wang, W.; Zhang, X.; Su, J. Kaempferol Attenuates ROS-Induced Hemolysis and the Molecular Mechanism of Its Induction of Apoptosis on Bladder Cancer. Molecules 2018, 23, 2592. [Google Scholar] [CrossRef]
- Ahmed, H.; Moawad, A.; Owis, A.; AbouZid, S.; Ahmed, O. Flavonoids of Calligonum Polygonoides and their Cytotoxicity. Pharm. Biol. 2016, 54, 2119–2126. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cai, F.; Zha, D.; Wang, X.; Zhang, W.; He, Y.; Huang, Q.; Zhuang, H.; Hua, Z.C. Astragalin-induced Cell Death is Caspase-dependent and Enhances the Susceptibility of Lung Cancer Cells to Tumor Necrosis Factor by Inhibiting the NF-кB Pathway. Oncotarget 2017, 8, 26941–26958. [Google Scholar] [CrossRef]
- Koczurkiewicz, P.; Czyż, J.; Podolak, I.; Wójcik, K.; Galanty, A.; Janeczko, Z.; Michalik, M. Multidirectional Effects of Triterpene Saponins on Cancer Cells—Mini-review of in vitro Studies. Acta. Biochim. Pol. 2015, 62, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Ayeleso, T.B.; Matumba, M.G.; Mukwevho, E. Oleanolic Acid and Its Derivatives: Biological Activities and Therapeutic Potential in Chronic Diseases. Molecules 2017, 22, 1915. [Google Scholar] [CrossRef]
- Zhu, Y.Y.; Huang, H.Y.; Wu, Y.L. Anticancer and Apoptotic Activities of Oleanolic Acid are Mediated Through Cell Cycle Arrest and Disruption of Mitochondrial Membrane Potential in HepG2 Human Hepatocellular Carcinoma Cells. Mol. Med. Rep. 2015, 12, 5012–5018. [Google Scholar] [CrossRef] [PubMed]
- Faggian, M.; Sut, S.; Perissutti, B.; Baldan, V.; Grabnar, I.; Dall’Acqua, S. Natural Deep Eutectic Solvents (NADES) as a Tool for Bioavailability Improvement: Pharmacokinetics of Rutin Dissolved in Proline/Glycine after Oral Administration in Rats: Possible Application in Nutraceuticals. Molecules 2016, 21, 1531. [Google Scholar] [CrossRef] [PubMed]
- Bobek, P.; Galbavý, S.; Mariássyová, M. The Effect of Red Beet (Beta vulgaris var. rubra) Fiber on Alimentary Hypercholesterolemia and Chemically Induced Colon Carcinogenesis in Rats. Nahrung 2000, 44, 184–187. [Google Scholar] [CrossRef]
- von Elbe, J.H.; Schwartz, S.J. Absence of Mutagenic Aactivity and a Short-term Toxicity Study of Beet Pigments as Food Colorants. Arch. Toxicol. 1981, 49, 93–98. [Google Scholar] [CrossRef]
- Kapadia, G.J.; Rao, G.S. Anticancer Effects of Red Beet Pigments. In Red Beet Biotechnology; Neelwarne, B., Ed.; Springer: Boston, MA, USA, 2013; pp. 125–154. [Google Scholar]
- Kapadia, G.J.; Azuine, M.A.; Sridhar, R.; Okuda, Y.; Tsuruta, A.; Ichiishi, E.; Mukainake, T.; Takasaki, M.; Konoshima, T.; Nishino, H.; et al. Chemoprevention of DMBA-induced UV-B Promoted, NOR-1-induced TPA Promoted Skin Carcinogenesis, and DEN-induced Phenobarbital Promoted Liver Tumors in Mice by Extract of Beetroot. Pharmacol. Res. 2003, 47, 141–148. [Google Scholar] [CrossRef]
- Ninfali, P.; Antonini, E.; Frati, A.; Scarpa, E.S. C-Glycosyl Flavonoids from Beta vulgaris Cicla and Betalains from Beta vulgaris rubra: Antioxidant, Anticancer and Antiinflammatory Activities-A Review. Phytother. Res. 2017, 31, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Zielińska-Przyjemska, M.; Olejnik, A.; Dobrowolska-Zachwieja, A.; Łuczak, M.; Baer-Dubowska, W. DNA Damage and Apoptosis in Blood Neutrophils of Inflammatory Bowel Disease Patients and in Caco-2 Cells in vitro Exposed to Betanin. Postepy Hig Med. Dosw (Online) 2016, 70, 265–271. [Google Scholar] [CrossRef]
- Lee, C.H.; Wettasinghe, M.; Bolling, B.W.; Ji, L.L.; Parkin, K.L. Betalains, Phase II Enzyme-inducing Components from Red Beetroot (Beta vulgaris L.) Extracts. Nutr. Cancer 2005, 53, 91–103. [Google Scholar] [CrossRef]
- Zou, D.M.; Brewer, M.; Garcia, F.; Feugang, J.M.; Wang, J.; Zang, R.; Liu, H.; Zou, C. Cactus Pear: A Natural Product in Cancer Chemoprevention. Nutr. J. 2005, 4. [Google Scholar] [CrossRef]
- Rahimi, P.; Abedimanesh, S.; Mesbah-Namin, S.A.; Ostadrahimi, A. Betalains, the Nature-inspired Pigments, in Health and Diseases. Crit. Rev. Food Sci. Nutr. 2018. [Google Scholar] [CrossRef]
- Lechner, J.F.; Wang, L.S.; Rocha, C.M.; Larue, B.; Henry, C.; McIntyre, C.M.; Riedl, K.M.; Schwartz, S.J.; Stoner, G.D. Drinking Water with Red Beetroot Food Color Antagonizes Esophageal Carcinogenesis in N-Nitrosomethylbenzylamine-treated Rats. J. Med. Food. 2010, 13, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Yi, E.Y.; Kim, Y.J. Betaine Inhibits in vitro and in vivo Angiogenesis Through Suppression of the NF-kB and Akt Signaling Pathways. Int. J. Oncol. 2012, 41, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Perez-Ternero, C.; Macià, A.; de Sotomayor, M.A.; Parrado, J.; Motilva, M.J.; Herrera, M.D. Bioavailability of the Ferulic Acid-derived Phenolic Compounds of a Rice Bran Enzymatic Extract and their Activity Against Superoxide Production. Food Funct. 2017, 8, 2165–2174. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, P.; Zhao, P.; Wang, D.; Zhang, Y.; Wang, J.; Chen, L.; Guo, W.; Gao, H.; Jiao, Y. Pretreatment of Ferulic Acid Attenuates Inflammation and Oxidative Stress in a Rat Model of Lipopolysaccharide-induced Acute Respiratory Distress Syndrome. Int. J. Immunopathol. Pharmacol. 2018. [Google Scholar] [CrossRef]
- Pei, K.; Ou, J.; Huang, J.; Ou, S. p-Coumaric Acid and its Conjugates: Dietary Sources, Pharmacokinetic Properties and Biological Activities. J. Sci. Food. Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef] [PubMed]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.M.; Suresh Kumar, C. Syringic acid (SA) —A Review of Its Occurrence, Biosynthesis, Pharmacological and Industrial Importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Ha, S.J.; Lee, J.; Park, J.; Kim, Y.H.; Lee, N.H.; Kim, Y.E.; Song, K.M.; Chang, P.S.; Jeong, C.H.; Jung, S.K. Syringic Acid Prevents Skin Carcinogenesis via Regulation of NoX and EGFR Signaling. Biochem. Pharmacol. 2018, 154, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J. Rhamnetin Attenuates Melanogenesis by Suppressing Oxidative Stress and Pro-inflammatory Mediators. Biol. Pharm. Bull. 2013, 36, 1341–1347. [Google Scholar] [CrossRef]
- Hu, T.; Liu, Q.M.; He, X.W.; Huang, F.; Zhang, M.W.; Jiang, J.G. Identification of Bioactives from Astragalus chinensis L.f. and their Antioxidant, Anti-inflammatory and Anti-proliferative Effects. Food Sci. Technol. 2017, 54, 4315–4323. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.J.; Jo, H.J.; Lee, K.J.; Choi, J.W.; An, J.H. Oleanolic Acid Induces p53-dependent Apoptosis via the ERK/JNK/AKT Pathway in Cancer Cell Lines in Prostatic Cancer Xenografts in Mice. Oncotarget 2018, 9, 26370–26386. [Google Scholar] [CrossRef] [PubMed]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, Pharmacology and Treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [PubMed]
- Concha, J.; Weinstein, C.; Zuniga, M.E. Production of Pectic Extracts from Sugar Beet Pulp with Antiproliferative Activity on a Breast Cancer Cell Line. Front. Chem. Sci. Eng. 2013, 7, 482–489. [Google Scholar] [CrossRef]
- Clifford, T.; Howatson, G.; West, D.J.; Stevenson, E.J. The Potential Benefits of Red Beetroot Supplementation in Health and Disease. Nutrients 2015, 7, 2801–2822. [Google Scholar] [CrossRef] [Green Version]
- Esatbeyoglu, T.; Wagner, A.E.; Schini-Kerth, V.B.; Rimbach, G. Betanin-A Food Colorant with Biological Activity. Mol. Nutr. Food Res. 2015, 59, 36–47. [Google Scholar] [CrossRef]
- Schwartz, S.J.; von Elbe, J.H.; Pariza, M.W.; Goldsworthy, T. Pitot HC. Inability of Red Beet Betalain Pigments to Initiate or Promote Hepatocarcinogenesis. Food Chem. Toxicol. 1983, 21, 531–535. [Google Scholar] [CrossRef]
- Klewicka, E. Fermented Beetroot Juice as a Factor Limiting Chemical Mutations Induced by MNNG in Salmonella typhimurium TA98 and TA100 Strains. Biotechnology 2010, 48, 229–233. [Google Scholar]
- Paluszczak, J.; Krajka-Kuźniak, V.; Baer-Dubowska, W. The Effect of Dietary Polyphenols on the Epigenetic Regulation of Gene Expression in MCF7 Breast Cancer Cells. Toxicol. Lett. 2010, 192, 119–125. [Google Scholar] [CrossRef]
- Tesoriere, L.; Allegra, M.; Butera, D.; Livrea, M.A. Absorption, Excretion, and Distribution of Dietary Antioxidant Betalains in LDLs: Potential Health Effects of Betalains in Humans. Am. J. Clin. Nutr. 2004, 80, 941–945. [Google Scholar] [CrossRef] [PubMed]
- Clifford, T.; Constantinou, C.M.; Keane, K.M.; West, D.J.; Howatson, G.; Stevenson, E.J. The Plasma Bioavailability of Nitrate and Betanin from Beta vulgaris rubra in Humans. Eur. J. Nutr. 2017, 56, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I. Plant Betalains: Safety, Antioxidant Activity, Clinical Efficacy, and Bioavailability. Compre. Rev. Food Sci. Food Safety 2016, 15, 316–330. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Wagner, A.E.; Motafakkerazad, R.; Nakajima, Y.; Matsugo, S.; Rimbach, G. Free Radical Scavenging and Antioxidant Activity of Betanin: Electron Spin Resonance Spectroscopy Studies and Studies in Cultured Cells. Food Chem. Toxicol. 2014, 73, 119–126. [Google Scholar] [CrossRef]
- Nowacki, L.; Vigneron, P.; Rotellini, L.; Cazzola, H.; Merlier, F.; Prost, E.; Ralanairina, R.; Gadonna, J.P.; Rossi, C.; Vayssade, M. Betanin-Enriched Red Beetroot (Beta vulgaris L.) Extract Induces Apoptosis and Autophagic Cell Death in MCF-7 Cells. Phytother. Res. 2015, 29, 1964–1973. [Google Scholar] [CrossRef]
- Zielińska-Przyjemska, M.; Olejnik, A.; Kostrzewa, A.; Łuczak, M.; Jagodziński, P.P.; Baer-Dubowska, W. The Beetroot Component Betanin Modulates ROS Production, DNA Damage and Apoptosis in Human Polymorphonuclear Neutrophils. Phytother. Res. 2012, 26, 845–852. [Google Scholar] [CrossRef]
- Martinez, R.M.; Longhi-Balbinot, D.T.; Zarpelon, A.C.; Staurengo-Ferrari, L.; Baracat, M.M.; Georgetti, S.R.; Sassonia, R.C.; Verri, W.A., Jr.; Casagrande, R. Anti-inflammatory Activity of Betalain-rich Dye of Beta vulgaris: Effect on Edema, Leukocyte Recruitment, Superoxide Anion and Cytokine Production. Arch. Pharm. Res. 2015, 38, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Allegra, M.; Furtmüller, P.G.; Jantschko, W.; Zederbauer, M.; Tesoriere, L.; Livrea, M.A.; Obinger, C. Mechanism of Interaction of Betanin and Indicaxanthin with Human Myeloperoxidase and Hypochlorous Acid. Biochem. Biophys. Res. Commun. 2005, 332, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G. HOCl and the Control of Oncogenesis. J. Inorg. Biochem. 2018, 179, 10–23. [Google Scholar] [CrossRef]
- Aratani, Y. Myeloperoxidase: Its Role for Host Defense, Inflammation, and Neutrophil Function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef]
- El Gamal, A.A.; AlSaid, M.S.; Raish, M.; Al-Sohaibani, M.; Al-Massarani, S.M.; Ahmad, A.; Hefnawy, M.; Al-Yahya, M.; Basoudan, O.A.; Rafatullah, S. Beetroot (Beta vulgaris L.) Extract Ameliorates Gentamicin-induced Nephrotoxicity Associated Oxidative Stress, Inflammation, and Apoptosis in Rodent Model. Mediators Inflamm. 2014. [Google Scholar] [CrossRef]
- Krajka-Kuźniak, V.; Paluszczak, J.; Szaefer, H.; Baer-Dubowska, W. Betanin, a Beetroot Component, Induces Nuclear Factor Erythroid-2-related Factor 2-mediated Expression of Detoxifying/antioxidant Enzymes in Human Liver Cell Lines. Br. J. Nutr. 2013, 110, 2138–2149. [Google Scholar] [CrossRef] [PubMed]
- Chyau, C.C.; Chu, C.C.; Chen, S.Y.; Duh, P.D. The Inhibitory Effects of Djulis (Chenopodium formosanum) and Its Bioactive Compounds on Adipogenesis in 3T3-L1 Adipocytes. Molecules 2018, 23, 1780. [Google Scholar] [CrossRef]
- Klewicka, E.; Nowak, A.; Zduńczyk, Z.; Cukrowska, B.; Błasiak, J. Protective Effect of Lactofermented Beetroot Juice Against Aberrant Crypt Foci Formation and Genotoxicity of Fecal Water in Rats. Exp. Toxicol. Pathol. 2012, 64, 599–604. [Google Scholar] [CrossRef]
- Kapadia, G.J.; Tokuda, H.; Konoshima, T.; Nishino, H. Chemoprevention of Lung and Skin Cancer by Beta vulgaris (beet) Root Extract. Cancer Lett. 1996, 100, 211–214. [Google Scholar] [CrossRef]
- Kapadia, G.J.; Azuine, M.A.; Rao, G.S.; Arai, T.; Iida, A.; Tokuda, H. Cytotoxic effect of the red beetroot (Beta vulgaris L.) extract compared to doxorubicin (Adriamycin) in the human prostate (PC-3) and breast (MCF-7) cancer cell lines. Anticancer Agents Med. Chem. 2011, 11, 280–284. [Google Scholar] [CrossRef]
- Zhang, Q.; Pan, J.; Wang, Y.; Lubet, R.; You, M. Beetroot Red (Betanin) Inhibits Vinyl Carbamate- and Benzo(a)pyrene-induced Lung Tumorigenesis Through Apoptosis. Mol. Carcinog. 2013, 52, 686–691. [Google Scholar] [CrossRef]
- Guldiken, B.; Toydemir, G.; Nur Memis, K.; Okur, S.; Boyacioglu, D.; Capanoglu, E. Home-Processed Red Beetroot (Beta vulgaris L.) Products: Changes in Antioxidant Properties and Bioaccessibility. Int. J. Mol. Sci. 2016, 17, 858. [Google Scholar] [CrossRef]
- Singh, C.K.; Siddiqui, I.A.; El-Abd, S.; Mukhtar, H.; Ahmad, N. Combination Chemoprevention with Grape Antioxidants. Mol. Nutr. Food Res. 2016, 60, 1406–1415. [Google Scholar] [CrossRef] [Green Version]


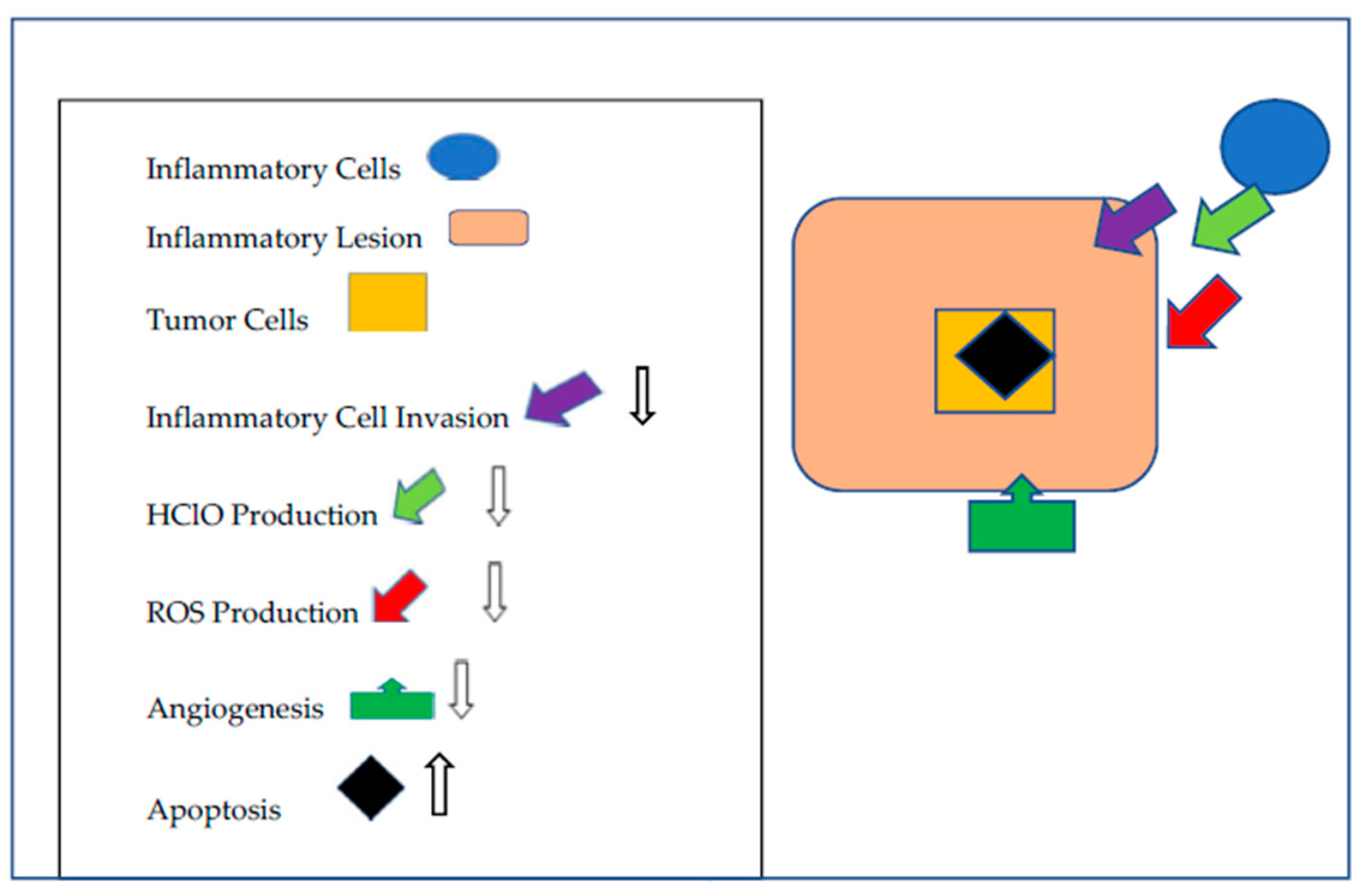{kind=link}
| Compound | Beneficial Effects | References |
|---|---|---|
| Betaine | Disrupt inflammation | [18,19,20] |
| Ferulic Acid | Membrane antioxidant, reduce NADPH oxidase activity and superoxide production | [21,22,23] |
| Caffeic Acid | Reduced lipid peroxidation, oxidative stress, inflammation | [24,25,26] |
| p-Coumaric Acid | Inhibits AKT and ERK signaling, angiogenesis, up-regulation of phase II enzymes, apoptosis | [27,28,29] |
| Syringic Acid | Chemoprevention activity in mice, Inhibit EGFR, Akt and NADPH oxidase activities | [30] |
| Rutin | Chemoprevention activity in mice, down-regulation of inflammation | [31] |
| Kaempferol | Apoptosis in vitro | [32,33] |
| Rhamnetin | Anti-oxidant, anti-inflammatory | [34] |
| Rhamnocitrin | Cytotoxicity | [35] |
| Astragalin | Inhibited growth of xenotransplanted tumor cells in nude mice | [36] |
| Oleanolic Acid | Inhibited growth of xenotransplanted tumor cells in nude mice | [37,38,39] |
| β-carotene | Anti-oxidant, increased apoptosis | [40] |
| Lutein | Anti-oxidant, activated macrophages, up-regulated pro-apoptotic genes | [40] |
| Non-defined | Reduced pre-cancerous lesions in rat | [41] |
| Betalains | Anti-oxidant, decreased inflammation, increased apoptosis, anti-mutagenic, induce phase II enzymes, chemopreventive activity | [42,43,44,45,46,47,48,49,50] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lechner, J.F.; Stoner, G.D. Red Beetroot and Betalains as Cancer Chemopreventative Agents. Molecules 2019, 24, 1602. https://doi.org/10.3390/molecules24081602
Lechner JF, Stoner GD. Red Beetroot and Betalains as Cancer Chemopreventative Agents. Molecules. 2019; 24(8):1602. https://doi.org/10.3390/molecules24081602
Chicago/Turabian StyleLechner, John F., and Gary D. Stoner. 2019. "Red Beetroot and Betalains as Cancer Chemopreventative Agents" Molecules 24, no. 8: 1602. https://doi.org/10.3390/molecules24081602
APA StyleLechner, J. F., & Stoner, G. D. (2019). Red Beetroot and Betalains as Cancer Chemopreventative Agents. Molecules, 24(8), 1602. https://doi.org/10.3390/molecules24081602