
Relaxant Effect of Monoterpene (−)-Carveol on Isolated Human Umbilical Cord Arteries and the Involvement of Ion Channels

 ,
,  ,
,  , and
, and 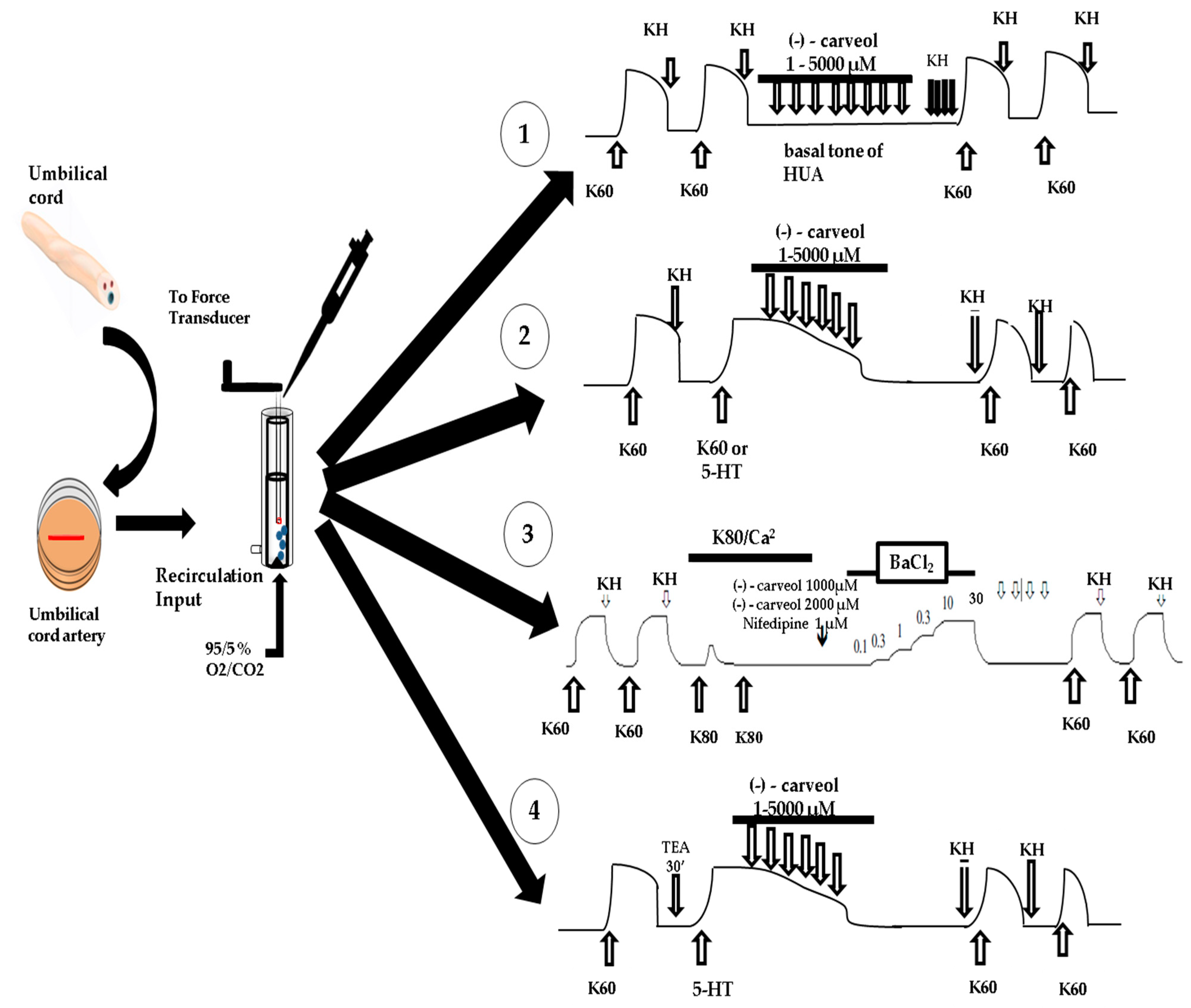{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Experimental Section
2.1. Solutions and Drugs
2.2. Tissue Preparation and Isolation
2.3. Determination of the Tension Exerted on the HUA Rings
2.4. Statistical Analysis
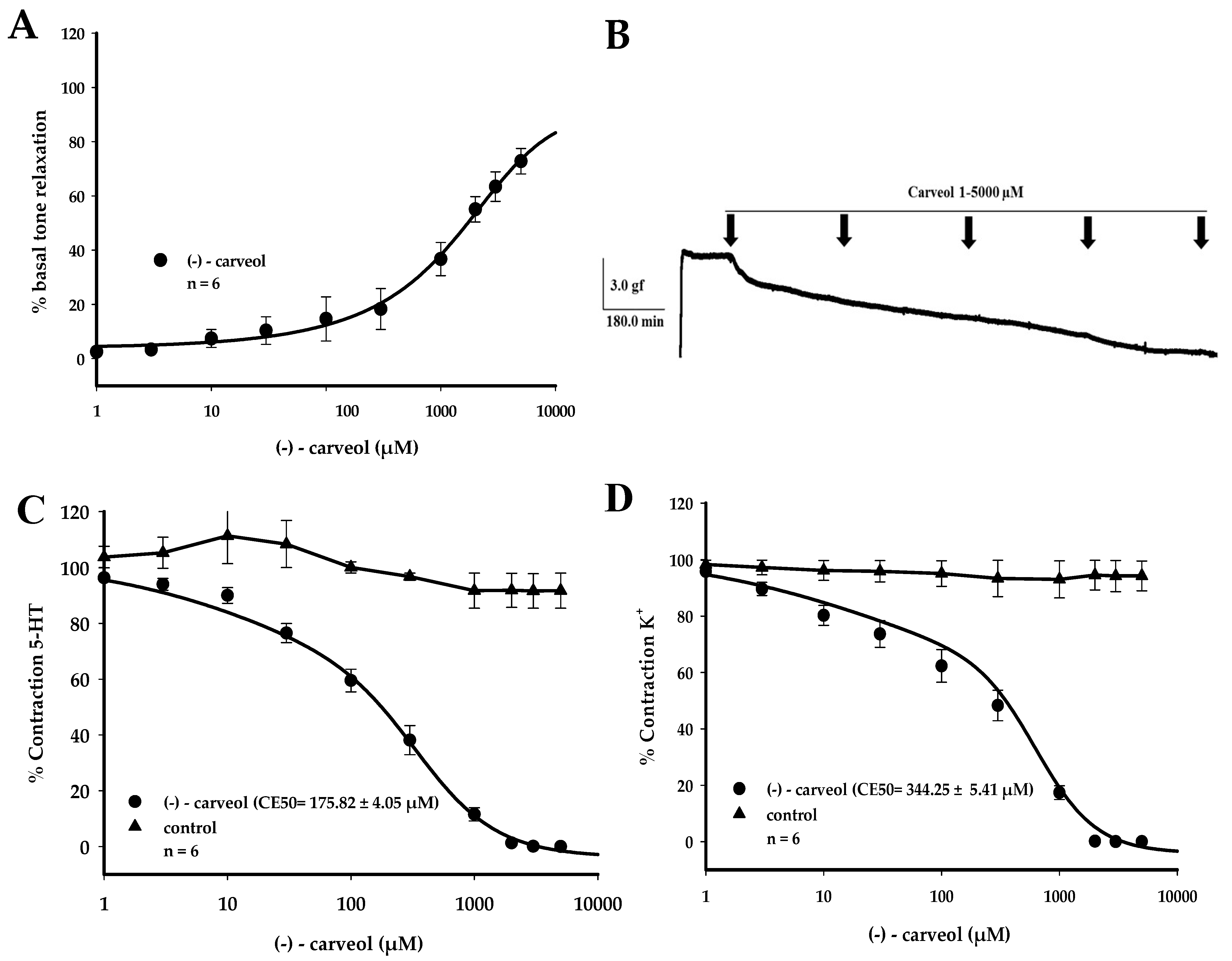
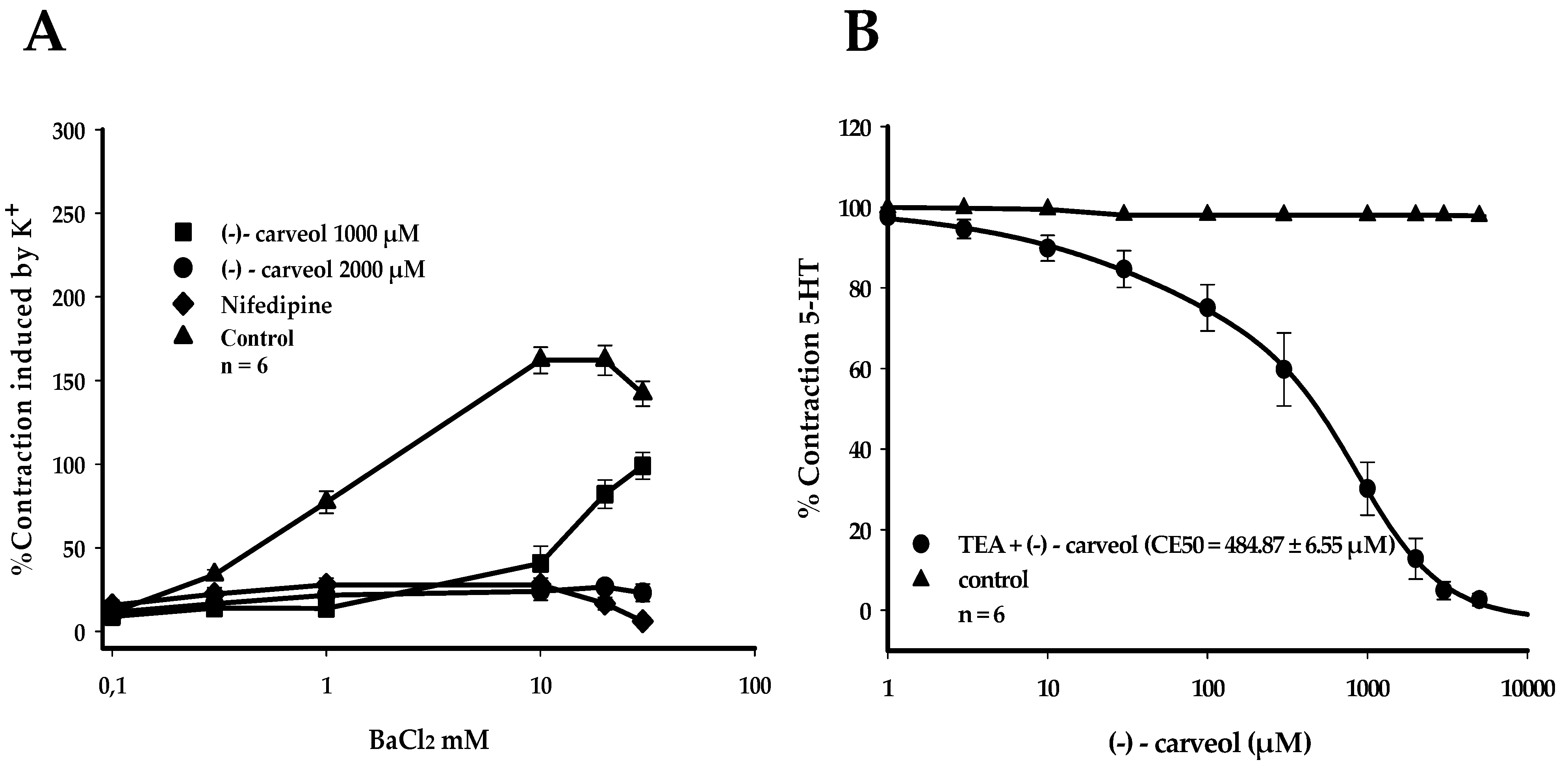
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lacerda-Neto, L.J.; Barbosa, A.G.R.; Quintans-Junior, L.J.; Coutinho, H.D.M.; Da Cunha, F.A.B. The complex pharmacology of natural products. Future Med. Chem. 2019, 11, 797–799. [Google Scholar] [CrossRef]
- Schmitt, E.K.; Moore, C.M.; Krastel, P.; Petersen, F. Natural products as catalysts for innovation: A pharmaceutical industry perspective. Curr. Opin. Chem. Biol. 2011, 15, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. The Mevalonate and Methylerythritol Phosphate Pathways: Terpenoids and Steroids. In Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 187–310. ISBN 9780470741689. [Google Scholar]
- Bhatia, S.P.; McGinty, D.; Letizia, C.S.; Api, A.M. Fragrance material review on carveol. Food Chem. Toxicol. 2008, 46, 85–87. [Google Scholar] [CrossRef] [PubMed]
- Omolo, M.O.; Okinyo, D.; Ndiege, I.O.; Lwande, W.; Hassanali, A. Repellency of essential oils of some Kenyan plants against Anopheles gambiae. Phytochemistry 2004, 65, 2797–2802. [Google Scholar] [CrossRef] [PubMed]
- Echeverrigaray, S.; Zacaria, J.; Beltrão, R. Nematicidal activity of monoterpenoids against the root-knot nematode Meloidogyne incognita. Phytopathology 2010, 100, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial activity of terpenes and terpenoids present in essential oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [PubMed]
- Rossi, Y.E.; Palacios, S.M. Fumigant toxicity of Citrus sinensis essential oil on Musca domestica L. adults in the absence and presence of a P450 inhibitor. Acta Tropica 2013, 127, 33–37. [Google Scholar] [CrossRef]
- Wagner, K.H.; Elmadfa, I. Biological relevance of terpenoids: Overview focusing on mono-, di- and tetraterpenes. Ann. Nutr. Metab. 2003, 47, 95–106. [Google Scholar] [CrossRef]
- Marques, F.M.; Figueira, M.M.; Schmitt, E.F.P.; Kondratyuk, T.P.; Endringer, D.C.; Scherer, R.; Fronza, M. In vitro anti-inflammatory activity of terpenes via suppression of superoxide and nitric oxide generation and the NF-κB signalling pathway. Inflammopharmacology 2019, 27, 281–289. [Google Scholar] [CrossRef]
- Li Lin, A.; Shangari, N.; Chan, T.S.; Remirez, D.; O’Brien, P.J. Herbal monoterpene alcohols inhibit propofol metabolism and prolong anesthesia time. Life Sci. 2006, 79, 21–29. [Google Scholar] [CrossRef]
- Cardoso-Teixeira, A.C.; Ferreira-Da-Silva, F.W.; Peixoto-Neves, D.; Oliveira-Abreu, K.; Pereira-Gonçalves, Á.; Coelho-De-Souza, A.N.; Leal-Cardoso, J.H. Hydroxyl group and vasorelaxant effects of perillyl alcohol, carveol, limonene on aorta smooth muscle of rats. Molecules 2018, 23, 1430. [Google Scholar] [CrossRef] [PubMed]
- Medina-Leyte, D.J.; Domínguez-Pérez, M.; Mercado, I.; Villarreal-Molina, M.T.; Jacobo-Albavera, L. Use of human umbilical vein endothelial cells (HUVEC) as a model to study cardiovascular disease: A review. Appl. Sci. 2020, 10, 938. [Google Scholar] [CrossRef]
- Velarde, F.; Castañeda, V.; Morales, E.; Ortega, M.; Ocaña, E.; Álvarez-Barreto, J.; Grunauer, M.; Eguiguren, L.; Caicedo, A. Use of Human Umbilical Cord and Its Byproducts in Tissue Regeneration. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Fei, J.Q.; Bin Zhou, H.; Shen, Y.L.; Chen, X.Z.; Wang, L.L. A comparison study on the responses of umbilical arteries and thoracic aorts to the adrenergic receptor agonists. Cell Biology Int. 2008, 32, S55. [Google Scholar] [CrossRef]
- Kaye, A.D.; Kaye, A.M.; Urman, R.D.; Banik, R.K.; Berger, J.S. Essentials of pharmacology for anesthesia, pain medicine, and critical care (Peripheral Vasodilators). In Essentials of Pharmacology for Anesthesia, Pain Medicine, and Critical Care; Springer: New York, NY, USA, 2015; pp. 1–904. ISBN 9781461489481. [Google Scholar]
- Stefos, T.; Sotiriadis, A.; Vasilios, D.; Tsirkas, P.; Korkontzelos, I.; Avgoustatos, F.; Lolis, D. Umbilical cord length and parity-The Greek experience. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 107, 41–44. [Google Scholar] [CrossRef]
- Yankowitz, J. Pharmacologic treatment of hypertensive disorders during pregnancy. J. Perinat. Neonatal Nurs. 2004, 18, 230–240. [Google Scholar] [CrossRef]
- Constantine, G.; Reynolds, A.L.; Luesley, D.M.; BEEVERS, D.G. Nifedipine as a second line antihypertensive drug in pregnancy. BJOG An. Int. J. Obstet. Gynaecol. 1987, 94, 1136–1142. [Google Scholar] [CrossRef]
- De Sousa, D.P.; De Farias nóbrega, F.F.; De Almeida, R.N. Influence of the chirality of (R)-(−)- and (S)-(+)-carvone in the central nervous system: A comparative study. Chirality 2007, 19, 264–268. [Google Scholar] [CrossRef]
- Santos-Silva, A.J.; Cairrão, E.; Morgado, M.; Álvarez, E.; Verde, I. PDE4 and PDE5 regulate cyclic nucleotides relaxing effects in human umbilical arteries. Eur. J. Pharmacol. 2008, 582, 102–109. [Google Scholar] [CrossRef]
- Mildenberger, E.; Biesel, B.; Siegel, G.; Versmold, H.T. Nitric oxide and endothelin in oxygen-dependent regulation of vascular tone of human umbilical vein. Am. J. Physiol. Circ. Physiol. 2003, 285, 1730–1737. [Google Scholar] [CrossRef][Green Version]
- Santos-silva, A.J.; Cairrão, E.; Verde, I. Study of the mechanisms regulating human umbilical artery contractility. Health (Irvine. Calif). 2010, 2, 321–331. [Google Scholar] [CrossRef]
- Rembold, C.M. Regulation of contraction and relaxation in arterial smooth muscle. Hypertension 1992, 20, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.; Menice, C.B.; Laporte, R.; Morgan, K.G. Mechanisms of smooth muscle contraction. Physiol. Rev. 1996, 76, 967–1003. [Google Scholar] [CrossRef] [PubMed]
- Lorigo, M.; Mariana, M.; Feiteiro, J.; Cairrao, E. How is the human umbilical artery regulated? J. Obstet. Gynaecol. Res. 2018, 44, 1193–1201. [Google Scholar] [CrossRef]
- Lovren, F.; Li, X.; Lytton, J.; Triggle, C. Functional characterization and m-RNA expression of 5-HT receptors mediating contraction in human umbilical artery. Br. J. Pharmacol. 1999, 127, 1247–1255. [Google Scholar] [CrossRef]
- Leung, S.W.S.; Quan, A.; Lao, T.T.; Man, R.Y.K. Efficacy of different vasodilators on human umbilical arterial smooth muscle under normal and reduced oxygen conditions. Early Hum. Dev. 2006, 82, 457–462. [Google Scholar] [CrossRef]
- Xie, H.; Triggle, C.R. Endothelium-lndependent Relaxations to Acetylcholine and A23187 in the Human Umbilical Artery. J. Vasc. Res. 1994, 31, 92–105. [Google Scholar] [CrossRef]
- Bertrand, C.; Duperron, L.; St-Louis, J. Umbilical and placental vessels: Modifications of their mechanical properties in preeclampsia. Am. J. Obstet. Gynecol. 1993, 168, 1537–1546. [Google Scholar] [CrossRef]
- Silva, R.E.R.; De Morais, L.P.; Silva, A.A.; Bastos, C.M.S.; Pereira-gonçalves, Á. Vasorelaxant effect of the Lippia alba essential oil and its major constituent, citral, on the contractility of isolated rat aorta. Biomed. Pharmacother. 2018, 108, 792–798. [Google Scholar] [CrossRef]
- Peixoto-Neves, D.; Silva-Alves, K.S.; Gomes, M.D.M.; Lima, F.C.; Lahlou, S.; Magalhães, P.J.C.; Ceccatto, V.M.; Coelho-De-Souza, A.N.; Leal-Cardoso, J.H. Vasorelaxant effects of the monoterpenic phenol isomers, carvacrol and thymol, on rat isolated aorta. Fundam Clin. Pharmacol. 2010, 24, 341–350. [Google Scholar]
- Perusquía, M.; Navarrete, E.; González, L.; Villalón, C.M. The modulatory role of androgens and progestins in the induction of vasorelaxation in human umbilical artery. Life Sci. 2007, 81, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.; Schmeda-Hirschmann, G.; Leiva, E.; Guzmán, L.; Orrego, R.; Fernández, P.; González, M.; Radojkovic, C.; Zuñiga, F.A.; Lamperti, L.; et al. Lemon grass (Cymbopogon citratus (D.C) Stapf) polyphenols protect human umbilical vein endothelial cell (HUVECs) from oxidative damage induced by high glucose, hydrogen peroxide and oxidised low-density lipoprotein. Food Chem. 2014, 151, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Lorigo, M.; Quintaneiro, C.; Lemos, M.C.; Martinez-De-Oliveira, J.; Breitenfeld, L.; Cairrao, E. UV-B filter octylmethoxycinnamate induces vasorelaxation by Ca2+ channel inhibition and guanylyl cyclase activation in human umbilical arteries. Int. J. Mol. Sci. 2019, 20, 1376. [Google Scholar] [CrossRef] [PubMed]
- Speroni, F.; Rebolledo, A.; Salemme, S.; Roldán-Palomo, R.; Rimorini, L.; Añón, M.C.; Spinillo, A.; Tanzi, F.; Milesi, V. Genistein effects on Ca2+ handling in human umbilical artery: Inhibition of sarcoplasmic reticulum Ca2+ release and of voltage-operated Ca2+ channels Efectos de la genisteína en los niveles del Ca2+ citosólico en células musculares de arteria umbilical hum. J. Physiol. Biochem. 2009, 65, 113–124. [Google Scholar] [CrossRef]
- Massaro, F.C.; Brooks, P.R.; Wallace, H.M.; Nsengiyumva, V.; Narokai, L.; Russell, F.D. Effect of australian propolis from stingless bees (Tetragonula carbonaria) on pre-contracted human and porcine isolated arteries. PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef]
- Marom, M.; Hagalili, Y.; Sebag, A.; Tzvier, L.; Atlas, D. Conformational changes induced in voltage-gated calcium channel Cav1.2 by BayK 8644 or FPL64176 modify the kinetics of secretion independently of Ca2+ influx. J. Biol. Chem. 2010, 285, 6996–7005. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.T.; Quayle, J.M. Channels in arterial smooth muscle\I. Am. J. Physiol. 1995, 268, C799–C822. [Google Scholar] [CrossRef] [PubMed]
- Gollasch, M.; Nelson, M.T. Voltage-Dependent Ca2+ channels in arterial smooth muscle cells. Kidney Blood Press. Res. 1997, 20, 355–371. [Google Scholar] [CrossRef]
- Salemme, S.; Rebolledo, A.; Speroni, F.; Milesi, V. L, P- / Q- and T-type Ca 2 + Channels in Smooth Muscle Cells from Human Umbilical Artery. Cell. Physiol. Biochem. 2007, 1900, 55–64. [Google Scholar] [CrossRef]
- Pereira-Gonçalves, Á.; Ferreira-da-Silva, F.W.; de Holanda-Angelin-Alves, C.M.; Cardoso-Teixeira, A.C.; Coelho-de-Souza, A.N.; Leal-Cardoso, J.H. 1,8-Cineole blocks voltage-gated L-type calcium channels in tracheal smooth muscle. Pflugers Arch. Eur. J. Physiol. 2018, 470, 1803–1813. [Google Scholar] [CrossRef]
- Carvalho, P.M.M.; Macêdo, C.A.F.; Ribeiro, T.F.; Silva, A.A.; Da Silva, R.E.R.; de Morais, L.P.; Kerntopf, M.R.; Menezes, I.R.A.; Barbosa, R. Effect of the Lippia alba (Mill.) N.E. Brown essential oil and its main constituents, citral and limonene, on the tracheal smooth muscle of rats. Biotechnol. Rep. 2018, 17, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Pereira-de-Morais, L.; de A. Silva, A.; da Silva, R.E.R.; da Costa, R.H.S.; Monteiro, Á.B.; dos S. Barbosa, C.R.; de S. Amorim, T.; de Menezes, I.R.A.; Kerntopf, M.R.; Barbosa, R. Tocolytic activity of the Lippia alba essential oil and its major constituents, citral and limonene, on the isolated uterus of rats. Chem. Biol. Interact. 2019, 297, 155–159. [Google Scholar] [PubMed]
- Milesi, V.; Raingo, J.; Rebolledo, A.; Grassi De Gende, A.O. Potassium channels in human umbilical artery cells. J. Soc. Gynecol. Investig. 2003, 10, 339–346. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evaristo Rodrigues da Silva, R.; de Alencar Silva, A.; Pereira-de-Morais, L.; de Sousa Almeida, N.; Iriti, M.; Kerntopf, M.R.; Menezes, I.R.A.d.; Coutinho, H.D.M.; Barbosa, R. Relaxant Effect of Monoterpene (−)-Carveol on Isolated Human Umbilical Cord Arteries and the Involvement of Ion Channels. Molecules 2020, 25, 2681. https://doi.org/10.3390/molecules25112681
Evaristo Rodrigues da Silva R, de Alencar Silva A, Pereira-de-Morais L, de Sousa Almeida N, Iriti M, Kerntopf MR, Menezes IRAd, Coutinho HDM, Barbosa R. Relaxant Effect of Monoterpene (−)-Carveol on Isolated Human Umbilical Cord Arteries and the Involvement of Ion Channels. Molecules. 2020; 25(11):2681. https://doi.org/10.3390/molecules25112681
Chicago/Turabian StyleEvaristo Rodrigues da Silva, Renata, Andressa de Alencar Silva, Luís Pereira-de-Morais, Nayane de Sousa Almeida, Marcello Iriti, Marta Regina Kerntopf, Irwin Rose Alencar de Menezes, Henrique Douglas Melo Coutinho, and Roseli Barbosa. 2020. "Relaxant Effect of Monoterpene (−)-Carveol on Isolated Human Umbilical Cord Arteries and the Involvement of Ion Channels" Molecules 25, no. 11: 2681. https://doi.org/10.3390/molecules25112681
APA StyleEvaristo Rodrigues da Silva, R., de Alencar Silva, A., Pereira-de-Morais, L., de Sousa Almeida, N., Iriti, M., Kerntopf, M. R., Menezes, I. R. A. d., Coutinho, H. D. M., & Barbosa, R. (2020). Relaxant Effect of Monoterpene (−)-Carveol on Isolated Human Umbilical Cord Arteries and the Involvement of Ion Channels. Molecules, 25(11), 2681. https://doi.org/10.3390/molecules25112681