1. Introduction
The cyanotoxin cylindrospermopsin (CYN) is a worldwide health risk.
Cylindrospermopsis (
Raphidiopsis) raciborskii and other species capable of synthesizing this molecule have been reported in water bodies of tropical and temperate regions of Australia, North America, South America, New Zealand and Europe. However, cyanobacterial species capable of producing CYN and its structural variants are found in almost all of the world’s latitudes [
1,
2]. In Brazil, the problem of cyanobacteria and their toxins in water is widely known and is a matter of public health policies [
3,
4,
5].
Exposure to CYN produces a fatty liver and hepatonecrosis in experimental animals, with extrahepatic lesions of variable location and severity. The main toxic effects appear to be irreversible protein synthesis inhibition, oxidative stress and deoxyribonucleic acid (DNA) damage. Nevertheless, comprehensive and specific mechanisms of toxicity are yet unclear [
4,
6].
Cylindrospermopsins (CYNs) are structural analogs of the originally reported CYN molecule. Limited information is available on the distribution, occurrence and toxicity of other naturally occurring CYN-like secondary metabolites from cyanobacteria [
4,
6,
11,
12,
13,
14,
15,
16,
17].
Figure 1 shows the CYNs reported to date, along with their reported toxicity and calculated octanol/water partition coefficient (log P). CYN, 7-epi-cylindrospermopsin (7-epi-CYN) and 7-deoxy-cylindrospermopsin (7D-CYN) are the main zwitterions reported in freshwater samples and laboratory strains [
13,
16,
18]. These zwitterion variants consist of a negatively chargeable sulfate group, a positively chargeable tricyclic guanidine portion and a biologically active uracil group. In the case of CYN and 7-epi-CYN, the uracil moiety is bound to the heterocyclic rings by a hydroxylated asymmetric carbon. This hydroxylation is absent in 7D-CYN, as well as in 7D-desulfo-CYN and 7-deoxy-desulfo-12-acetyl-cylindrospermopsin. Additionally, the absence of the sulfate anion on these last two desulfo variants makes them less hydrophilic compared to the zwitterion variants.
Figure 1 also shows the structure of cylindrospermic acid and 5-chloro-cylindrospermopsin, two innocuous oxidation products reported in the literature. Desulfo-cylindrospermopsin (desulfo-CYN) is a degradation product that can be formed by acid hydrolysis.
Both 7D-CYN and 7D-desulfo-CYN are biosynthetic precursors of the originally reported CYN molecule. The functional guanidino moiety is the starting unit in the biosynthesis of the carbon backbone of CYN. An uncommon biosynthetic pathway of pyrimidine is responsible for the de novo formation of the uracil ring. Finally, sulfation and hydroxylation tailoring reactions add the last modifications that culminate in the fully bioactive cylindrospermopsin molecule [
19,
20].
Cylindrospermopsin is a widely studied molecule for which numerous extraction protocols and toxicity assays have been described. 7D-CYN is a zwitterion analog, structurally similar enough to be simultaneously isolated along with CYN in most frequently used extraction protocols [
11,
13,
21,
22]. However, other CYN-like metabolites are less reported. In 2014, Wimmer et al. reported the natural occurrence of two alkaloids with different hydrophobicity than the previously reported CYNs: 7D-desulfo-CYN and 7-deoxy-desulfo-12-acetylcylindrospermopsin [
15]. At present, there are no certified standards for 7D-desulfo-CYN and 7-deoxy-desulfo-12-acetylcylindrospermopsin. 7D-CYN-certified standards are available by several vendors. Nevertheless, 7D-CYN standards are currently sold in low concentrations that are inappropriate for the validation of analytical methods or biological testing purposes.
The chemical properties of the less-hydrophilic CYNs indicate that they may accumulate in other matrixes other than freshwater—for example, inside cyanobacterial cells, animal tissue, sediments or water filter resins. The distribution of desulfo CYNs in ecological systems is unknown.
7D-desulfo-CYN serves as an important analyte in order to study the importance of the C-7 hydroxylation in the lethal potency of CYN alkaloids that lack the sulfate group. Another desulfo analog, desulfo-CYN, can be synthesized by the acid hydrolysis of CYN at a constant temperature. Reports on the acute toxicity of desulfated CYNs are scarce, and the chronic effects of these desulfo variants are far from being elucidated. Here, we report a novel SPE workflow to isolate cylindrospermopsins (CYN, 7D-CYN and 7D-desulfo-CYN) simultaneously from the same aliquot of a natural producing strain in order to assess their lethal potencies on HepG2 cells.
3. Discussion
We present in this work new comparative toxicological studies in vitro of zwitterionic and desulfated CYN alkaloids. Comparative studies between naturally occurring CYN alkaloids are not common, mainly due to a lack of popular and accepted extraction protocols for these metabolites.
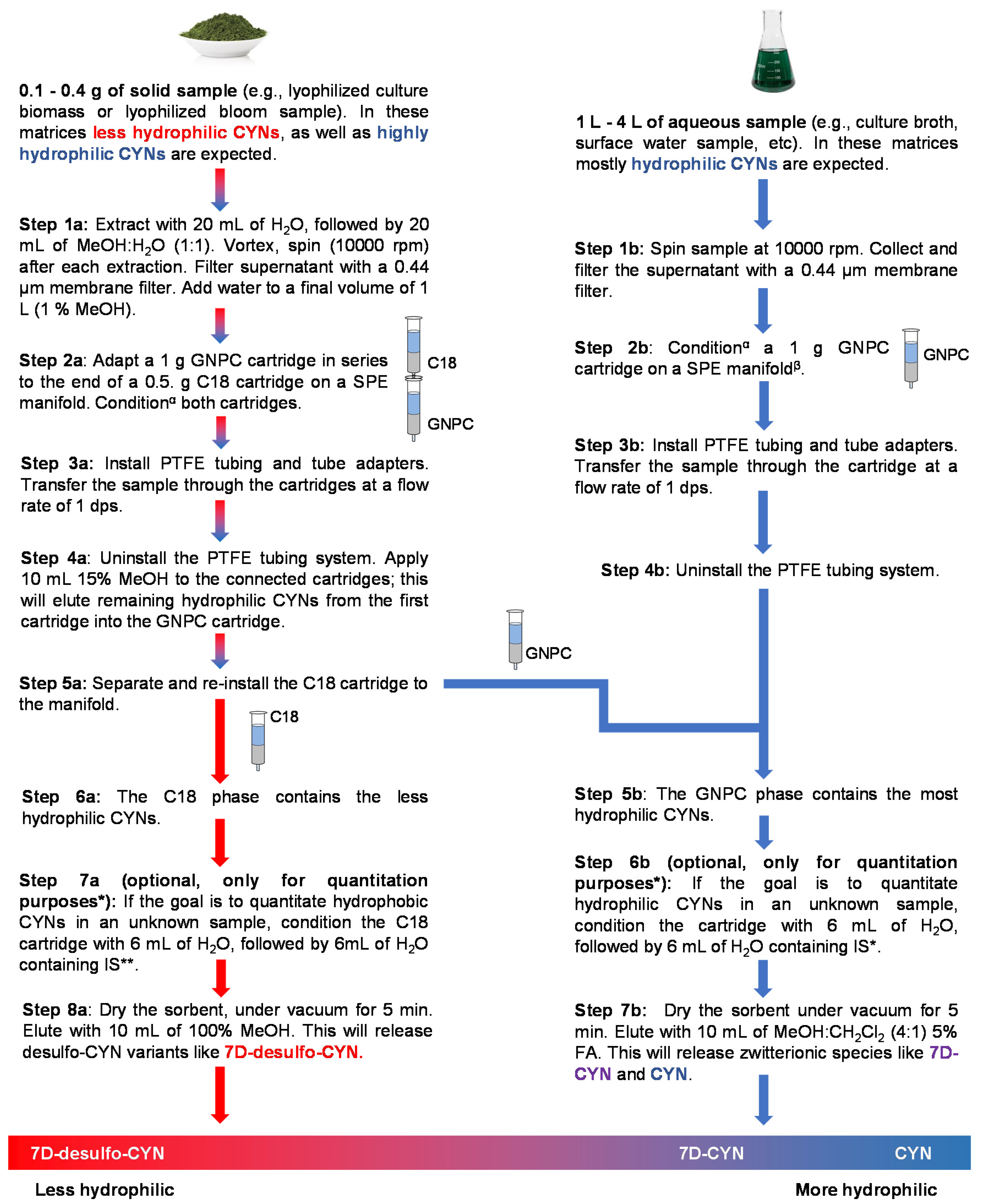
The workflow described in
Figure 2 aims to improve the monitoring of CYNs by concurrently isolating CYN variants with different water solubilities, depending on the nature of the sample. As evidenced in
Figures S1–S7, considerable effort was invested in testing different SPE sorbents’ abilities to retain the CYN analogs after applying various elution techniques. Our main concern was to implement efficient and selective elutions on the ideal sorbent, while ensuring the maximum recovery of each analyte.
Norris et al. (2001) documented the benefits of using graphitized carbon cartridges (instead of C18) to retain CYN zwitterions [
17]. Nevertheless, C18 SPE columns proved optimal in order to selectively adsorb less-hydrophilic CYNs [
15]. The coupling of these two SPE techniques while using selective eluents can improve the recovery of each unique CYN-like molecule.
Unknown solid samples composed mostly of the biomass may contain highly hydrophilic and, also, less-hydrophilic CYNs. As shown in
Figure 2 (Step 2a), after the application of the sample to the coupled cartridges, less-hydrophilic variants remain in the C18 phase, while zwitterions adsorb mainly on GNPC. Elution with 15% MeOH efficiently releases the remaining zwitterions from the C18 phase, and these, in turn, adsorb to the GNPC cartridge. Desulfo-CYNs in an unknown sample will remain in the C18 phase and can be subsequently eluted with pure MeOH. On the other hand, hydrophilic zwitterions bond strongly to the GNPC phase, and pure acidified MeOH is not sufficient to completely release these metabolites from the solid phase. However, the complete recovery of zwitterions from the carbon phase was reliably achieved with 10 mL of MeOH:CH
2Cl
2 4:1 with 5% FA; there was no need for successive elutions, even when extracting 0.4 g of the biomass from our toxin-producing strain.
The validation of the protocol only focused on CYN, due to the lack (at the time) of commercially available certified standards of 7D-CYN at concentrations useful to prepare spiked samples. As stated earlier, no certified standards of 7D-desulfo-CYN (or other desulfo alkaloids) are currently available. Consequently, we decided to only include in the validation plan steps 1b to 7b of the workflow (
Figure 2). As indicated in the Results (
Section 2.2) and
Supplementary Materials (Figures S8 and S9 and Tables S3 and S4), all the analytical performance parameters evaluated in the validation were under regulatory guidance requirements.
After confirming the quantity, purity and structure of the metabolites, we carried out toxicity assays on HepG2 cells. Fluorescence-activated cell sorting (FACS) flow cytometry was chosen as the ideal technique to test the purified CYNs’ toxicity. Some benefits of using FACS are that it monitors a representative sample of our cells (10,000 events), it is selective enough to identify the type of cell death (necrosis or apoptosis) and it is a highly sensitive technique because of the use of fluorescent markers.
Toxicity data resulting from our experiments matched the CYNs’ cell death results reported by Štraser et al. (2013) using annexin V and PI staining for FACS analysis. Their research found no significant difference in cell death between the vehicle control and treated HepG2 cells at 0.5 μg/mL for 12 and 24 h [
30]. Using a similar methodology (
Figure 5D), we did not find a significant difference in cell death between the control (100% viability) and cells treated with 1 µM (0.4 μg/mL) of the certified CYN standard during 12 and 24 h. Additionally consistent with Štraser et al.’s work, significant cell death was detected after a 48-h stimulation with 1 µM of the CYN-certified standard (
Figure 5D) and 1 µM of purified CYN (
Figure 6D). Intriguingly, a two-h treatment with 381 µM of CYN showed no significant viability decrease (data not shown). This was the highest concentration we could prepare with the available CYN-certified standard. Even though it is not a practical working concentration, the results with 381 µM of CYN served as evidence of the cyanotoxin’s delayed toxicity. In other words, the toxin’s apoptotic and/or necrotic effects are not immediate to HepG2 cells even at high concentrations.
In our current work, 7D-CYN caused no significant loss in viability at 1 µM (0.4 μg/mL) and 5 µM (2.0 μg/mL) after a 48-h treatment (
Figure 6D). These results are consistent with reports by Neumann et al. (2007). Their group implemented a cell proliferation assay using the 3-(4,5-dimethylthiazol-2-yl)-5-(-3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium inner salt (MTS) and the electron-coupling reagent phenazine ethosulfate (PES). They stimulated with 7D-CYN several cell lines—of which, HepG2 was the least affected—yielding an IC
50 of 3.2 μg/mL at 24 h and 3.1 μg/mL at 48 h [
12]. According to Looper et al. (2005), 7D-CYN is a potent inhibitor of protein synthesis in vitro. Nevertheless, a complete protein synthesis inhibition required 10 µM in rat hepatocytes, a concentration higher than the ones tested in our current work [
24].
Finally, we found that 7D-desulfo-CYN caused no significant cell death compared to the vehicle control at 1 µM (0.4 μg/mL) and 5 µM (2.0 μg/mL) after a 48-h treatment (
Figure 7D). In the same manner, no significant viability loss was detected using 1 µM and 5 µM of the in-house degradation product desulfo-CYN. The vehicle control cells were grown in culture medium containing 0.2% MeOH to account for the methanol content of the purified desulfo analogs’ fractions.
Several authors have stated the essential role of the guanidine, uracil and hydroxyl groups in the toxic mechanism of CYN [
23,
31,
32,
33]. On the other hand, it has been proposed that the sulfate group of CYN has no role in its toxicity or its transport through the rat hepatocyte’s membrane. According to Runnegar et al., the sulfate group at C-12 of CYN is not required to induce a protein synthesis inhibition, nor is it required for cellular uptake in primary rat hepatocytes (PRH) [
31]. In the current work, no significant apoptosis nor necrosis was detected in HepG2 cells after stimulation with up to 5 µM of two different desulfo variants. This incongruence can be explained by taking into account the cell type (PRH) and, also, the kind of biological activity assays (protein synthesis and cellular glutathione quantitation) used by Runnegar et al. (2002). Direct and indirect protein synthesis in vitro measurements are sensitive and specific of CYN alkaloids’ biological activity; these tests may not correlate well with cell viability assays (detection of the onset of necrosis and apoptosis). Likewise, PRH are known to be more sensitive to the toxic mechanisms of CYN. PRH display a wide variety of metabolic enzymes, membrane receptors and active transport proteins that resemble the liver parenchyma [
6,
34,
35]. The HepG2 hepatoma cell line lacks this diversity, making it less sensitive to the toxicity of CYN alkaloids [
12,
35]. Taking this into account, it is likely that higher doses of desulfo-CYN and 7D-desulfo-CYN are necessary to detect their lethal potency in HepG2 cells.
Interestingly, 7-deoxy-desulfo-CYNs tend to be less-hydrophilic than the zwitterionic alkaloids (according to the n-octanol and water partition coefficients in
Figure 1). Lipophilicity, commonly represented by the log P value, is the definitive factor in a molecule’s absorption, diffusion across cellular and tissue membranes, metabolism and excretion. Thus, a correlation between the lipophilicity and biological activity of small molecules is usually made [
36,
37]. In this way, the deoxy-desulfo analogs are favored with a better chance of passive membrane transport. Nevertheless, the enhanced capacity of desulfated analogs to penetrate cell membranes in comparison to zwitterionic species was not reflected in their lethality towards HepG2 cells. Only the original cylindrospermopsin alkaloid was able to significantly decrease viability in the hepatoma cell line at the tested conditions.
It has been demonstrated that 7D-desulfo-CYN and 7D-CYN correspond to the last two biosynthetic steps of CYN [
19]. In the last biosynthetic step, the cyrI gene of the cyr cluster encodes a 2-oxoglutarate-dependent hydroxylase that transforms 7-deoxy-cylindrospermopsin into the diastereomers CYN and 7-epi-CYN [
20]. The pharmacological significance of the C-7 hydroxyl group lies in being the last modification of a complex biosynthetic pathway in order to enable its biological activity. Previously, synthetic CYN analogs were used by other authors to test the pharmacological importance of the C-7 hydroxyl group [
32,
33]. Studies by Cartmell et al. (2017) compared the bioactivity of several synthetic analogs, revealing that the hydroxyl group at C-7 was indispensable in causing significant toxicity in human cells. The authors proposed that the C-7 hydroxyl group may be crucial for the active cellular transportation of CYN, analogous to the uptake of cholesterol. Evans et al. (2019) tested the biological activity of new synthetic CYN analogs with variations in their functional groups but all possessing the uracil ring. These experiments showed that the C-7 hydroxyl group, as well as the guanidine moiety, have a role in cylindrospermopsin’s toxicity and that both are essential for the molecule’s biological activity [
33].
The toxicity assays presented in this work indicate that, in our biological model, a combination of the guanidino, uracil, sulfate and C-7 hydroxyl functional groups is necessary for a CYN analog to efficiently induce cell death. Future investigations using desulfo CYN variants may include toxicity tests using 3D hepatocyte cultures, toxicity tests on PRH cultures and in vivo toxicokinetics studies. The isolation workflow proposed by our group may serve useful for the forthcoming research of uncommon CYN alkaloids.
4. Materials and Methods
4.1. Cylindrospermopsis raciborskii Strain Maintenance
The
Cylindrospermopsis raciborskii strain 11K is an Australian strain provided by Professor Marli Fatima Fiori at the Centro de Energía Nuclear na Agricultura (CENA), Universidade de São Paulo (Campus Luiz de Queiroz, Piracicaba). This strain produces CYN, 7D-CYN and 7D-deoxy-CYN. A Brazilian non-CYN-producing strain (
C. raciborskii ITEP 18) was grown and used as a matrix for CYN spiked-in extractions during the validation of the LC-DAD method. These strains were maintained in our laboratory using the ASM-1 culture medium proposed by Gorham et al. (1964), with some modifications [
38,
39].
Both cyanobacterial strains were cultured inside an incubator at the desired final volume, with a working temperature of 22 °C, a 12-h light/dark interval and a light intensity of 30 to 50 μmol. photons.m−2.s−1;calibrated by a LI-COR® 250A (LI-COR, Lincoln, NE, USA) light meter.
4.2. Tests on Sample Size and Cartridge Handling for Liquid Samples
Extraction experiments to assess the adsorption of the analytes in two different SPE sorbents required liquid samples obtained from the Cylindrospermopsis raciborskii strain 11K cultures with a density of 1 × 108 cells/mL.
Each lot of approximately 20 L of dense culture broth was centrifuged (10,000 rpm, 10 min, 4 °C), and the supernatant (liquid medium) was collected. The remaining precipitate (filamentous cells or cellular debris) was frozen at −20 °C before lyophilization. Next, we filtered the culture medium through a 0.44-µm membrane to avoid clogging of the SPE cartridges.
Small-scale experiments using the liquid medium were performed initially to assess the adsorption of the analytes (mostly CYN and 7D-CYN) on two SPE columns: C18 Sep-Pak 1g (Waters
®, Milford, MA, USA) and graphitized nonporous carbon (GNPC) Envi-carb 1g (Supelco
®, Bellefonte, PA, USA). On the first experiment, SPE columns were pretreated with 10 mL of MeOH, followed by 10 mL of water. After the SPE column conditioning, we applied 300 mL of liquid medium to each cartridge through a PTFE (polytetrafluoroethylene) tubing system connected by a SPE tube adapter. The adsorbed analytes were eluted using 5 mL of each of the following aqueous solutions: 10%, 50%, 75% and 100% MeOH (
Figures S1–S5). The GNPC cartridges were eluted in backflush mode (or reverse mode) due to the high affinity of the analytes to the stationary phase. The recovered volumes were dried and concentrated in 200-µL 50% MeOH before LC-DAD analysis.
Another small-scale experiment using the liquid medium was aimed to evaluate the percent recovery of CYNs using two elution techniques for the GNPC cartridges. GNPC columns were pretreated initially with 10 mL of MeOH. After this step, we applied 10 mL of water to each SPE column. Once the SPE columns were conditioned, we applied 300 mL of the liquid medium to each cartridge through the PTFE tubing system. The adsorbed analytes were eluted using two elution techniques: MeOH:CH
2Cl
2 (4:1) 5% FA (3 fractions of 5 mL) using the cartridge in the usual forward mode or 10%, 50% and 100% MeOH (3 fractions of 5 mL) using the cartridge in backflush mode (
Figure S6). The recovered volumes were dried and concentrated in 200 µL of 50% MeOH. We calculated the 100% recovery of every CYN alkaloid by quantitating the analytes on a lyophilized sample of the filtrated culture medium. LC-DAD-MS
2 was used to carry out the analysis in order to improve the confirmation of CYN variants at low recoveries.
Large-scale experiments using the liquid medium were performed to test the affinity of the analytes (mostly CYN and 7D-CYN) to the GNPC column when using larger sample volumes (1.5–4 L). GNPC cartridges were pretreated using 10 mL of MeOH, followed by 10 mL of water. After this conditioning step, we applied the desired volume of the liquid medium to the cartridge through the PTFE tubing system. Arbitrary volumes of 1.5, 3 and 4 L were applied once to each GNPC column. We eluted the adsorbed analytes once with 10 mL of MeOH:CH
2Cl
2 (4:1) 5% FA. The recovered eluates and effluents (
Figure S6) were dried and concentrated in 200-µL 50% MeOH before LC-DAD-MS
2 analysis.
4.3. Tests on Sample Size and Cartridge Handling for Solid Samples
Biomass experiments were intended to optimize the 7D-desulfo-CYN isolation from a complex sample. The biomass samples used in the present work consisted of lyophilized filamentous cells or cellular debris. We obtained these cyanobacterial remnants from the frozen precipitates derived from the centrifugation of dense culture broths of the Cylindrospermopsis raciborskii strain 11K, as stated earlier. After weighing the lyophilized sample, cells were suspended in 50% MeOH. In this suspension, cell membranes were disrupted using an ultrasonic probe sonicator. The lysate was centrifuged (10,000 rpm, 10 min, 4 °C), and the supernatant was transferred to a clean tube. The pellet was resuspended with 50% MeOH, sonicated, centrifuged and the supernatant was collected and mixed with the previous extract. The final extract was filtered through a 0.44 µm membrane and diluted with water to approximately 1% MeOH. These aqueous extracts were later permeated through the desired SPE columns through the PTFE tubing system.
The biomass experiments tested the adsorption of the analytes to the sorbents of Oasis HLB 0.5 g and C18 Sep-Pak 0.5 g cartridges (Waters®, Milford, MA, USA). The affinity of the analytes to the SPE sorbents while adjusting the sample’s pH (pH 4, 7 and 10) was also studied. The SPE columns were pretreated with 6-mL 100% MeOH, followed by 6 mL of water. While permeating the aqueous samples through the columns, the resulting effluent was collected and later lyophilized for analysis. We eluted the analytes from the sorbents with 6-mL 10% MeOH, followed by 6 mL pure MeOH. Both fractions were also collected for every sample. A similar experiment was performed with C18 cartridges only, in which the cartridges were eluted using 10-mL 15% MeOH, followed by 10 mL of pure MeOH (all fractions were collected for every sample).
4.4. Semipreparative Purification
The isolation of high-purity CYNs from the biomass or broth using SPE columns usually required this last purification step. We resuspended the GNPC extracts in Milli-Q® (MilliporeSigma, Burlington, MA, USA) grade H2O and C18 extracts in 20% MeOH (to improve 7D-desulfo-CYN’s solubility) before the chromatographic separation using a Luna 5 µm C18 100A, 250 × 10 mm (Phenomenex®, Torrance, CA, USA) HPLC column. The mobile phase consisted of Milli-Q® grade H2O, and elution of the CYN variants was achieved with a gradual increase of MeOH from 5% to 80% for 35 min. The semipreparative purification was carried out using an HPLC (LC20AD, Shimadzu Corporation, Kyoto, Japan) with a diode-array detector (SPD-M20A) coupled to a fraction collector (FRC-10A, Shimadzu Corporation, Kyoto, Japan). Detection of the CYN structural analogs was set between 200 and 600 nm using the diode-array detector.
4.5. Sample Preparation for LC-DAD Method Validation
A cyanobacterial strain confirmed by our lab as a nonproducer of CYN (
C. raciborskii ITEP 18) was grown according to
Section 4.1 and used as a matrix for CYN spiked-in extractions. Cultures with a density of 1 × 10
8 cells/mL were centrifuged, and the supernatant was collected. The resulting liquid medium was divided into aliquots, each with a volume of 300 mL. Acyclovir-certified standard (Thermo Fisher Scientific, Waltham, MA, USA) was used as the IS. Calibrators and control samples were prepared using different lots of CYN-certified standards (Eurofins Abraxis, Warminster, PA, USA).
4.6. LC-DAD Method Validation
Validation of the LC-DAD method was performed following regulatory guidelines established by the National Agency for Sanitary Vigilance, document RDC No. 166 (Brazil 2017) [
40,
41]. The assessed validation parameters were linearity, limits of detection (LOD), limits of quantification (LOQ), selectivity, carryover, within-day bias and within-day imprecision.
Linearity (R2 ≥ 0.99) was estimated by the analysis of calibration curves (n = 5). Calibrators were prepared at the following concentrations: 0.5, 5, 25, 50, 100 and 150 µg/mL. These calibrators were considered acceptable at ≤15% coefficient of variation (CV) (≤20% CV at the lower LOQ).
Estimation of the LOD and LOQ was performed using decreasing concentrations of CYN-spiked samples. The lower LOD was the lowest concentration, with a signal/noise ratio ≥3, and the lower LOQ was the lowest concentration, with a signal/noise ratio ≥10.
Within-day imprecision and within-day bias were calculated at five concentrations in five replicates. Imprecision was expressed as the coefficient of variation (% CV). Acceptable imprecision was ≤15% CV (≤20% CV at the lower LOQ was accepted). Bias was estimated for each concentration as the relative standard error (% RSE). Relative standard errors of 85–115% were considered acceptable, and 80–120% RSE were accepted at the lower LOQ.
The matrix effect was evaluated using blank samples. Interference in the DAD detector’s response was tested at the retention times of CYN and IS (acyclovir). Interference peaks of ≤20% of the analyte response at the lower LOQ were considered acceptable. Interference peaks of ≤5% of the IS response were considered acceptable.
Chromatographic selectivity was evaluated by calculating the peak resolution (Rs) of a nonextracted mix containing the following analytes: CYN (certified standard), acyclovir (certified standard), 7D-CYN (purified fraction), 7D-desulfo-CYN (purified fraction) and desulfo-CYN. The desulfo-CYN degradation product was synthesized by diluting a purified fraction of cylindrospermopsin in an aqueous solution of HCl 5% and maintaining it in constant shaking at 70 °C during a 48-h interval.
4.7. LC-DAD-MS2 Instrumentation
The extracts were separated through a Synergi 4 µm Hydro-RP 80A, 150 × 4.6 mm (Phenomenex
®, Torrance, CA, USA) HPLC column, protected with a guard column of the same material. The mobile phase consisted of 5-mmol/L ammonium acetate buffer with 0.05% FA. Several gradient elutions using MeOH were tested, according to Eaglesham et al. (1999) [
42] and Gallo et al. (2009) [
1]. The gradient elution of the final method started at a 100% aqueous mobile phase, with a gradual increase up to 80% MeOH at 15 min (flow 0.2 mL/min). Samples were resuspended in 50% MeOH, and 5 uL was injected into an HPLC (LC20AD, Shimadzu Corporation, Kyoto, Japan) with a diode-array detector (SPD-M20A, Shimadzu Corporation, Kyoto, Japan), coupled to a quadrupole time-of-flight mass spectrometer (Micro TOF-QII; Bruker Daltonics, Billerica, MA, USA) with an ESI (electrospray ionization) interphase.
The detection range for CYNs’ quantitation was set between 200 and 600 nm (λmax = 262 nm) using the diode-array detector. Since the diode-array detector was coupled to the Micro TOF-QII analyzer, quantitation and structure confirmation of CYN and structural analogs were carried out simultaneously. The ionization source conditions were as follows: capillary potential of 3500 V, temperature and flow of drying gas (nitrogen) of 300 °C and 5 mL/min, respectively, and a nebulizer pressure of 35 psi. Mass spectra were acquired using electrospray ionization in the positive mode, m/z range 50 to 3000. The Micro TOF-QII instrument was operated in MS1 scan mode or MS2 MRM (multiple reaction monitoring) mode as needed. In the MRM mode, CID (collision-induced dissociation) experiments were performed using the following collision energies for fragmentation: 20 eV, 35 eV and 40 eV.
4.8. Toxicity of the CYN Variants
HepG2 cells (Code 0103) purchased from Rio de Janeiro Cell Bank (BCRJ, Rio de Janeiro, Brazil) were grown in DMEM (Dulbecco’s Modified Eagle’s medium) high glucose (Gibco®, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin and 0.1 µg/mL streptomycin at 37 °C in a 5% CO2 atmosphere. HepG2 cells were seeded in 6-well plates with a density of 2.5 × 105 cells/well, incubated overnight and treated with the chosen CYN variant at the desired time interval. After the treatment intervals, cells were rinsed with phosphate-buffered saline (PBS). The cells were detached from the flask using 300 µL of 2.5 g/L trypsin solution (0.25% trypsin diluted in PBS) for 5 min. DMEM with 10% FBS (3 mL) was added to inactivate the trypsin, transferred to the collection tubes and centrifuged at 250× g for 5 min at 4 °C. The supernatant was discarded, and the pellet was washed using PBS with 4% FBS, followed by 200 µL of annexin V-binding buffer (10 mM Hepes pH 7.4, 140 mM NaCl and 2.5 mM CaCl2). We stained the cells using 5 µL of annexin V conjugated with allophycocyanin (annexin V-APC) (BDTM, East Rutherford, NJ, USA) and 2 µL of propidium iodide (PI) (Sigma-Aldrich, St. Louis, MO, USA). The samples were incubated for 15 min in the dark at room temperature. Cells treated with 10% dimethyl sulfoxide and with 40% methanol were used as positive controls for apoptosis and necrosis, respectively, and cells without any treatment were used as the negative control. We analyzed the samples on a flow cytometer FACS Canto II (BDTM, East Rutherford, NJ, USA), acquiring 10,000 events using the FACS Diva™ software, version 8.0 (BDTM, East Rutherford, NJ, USA). The excitation wavelength was λEX 633 nm and 480 nm and emission at λEM 660 nm and 578 nm for annexin V-APC and PI, respectively. The samples were analyzed using FlowJo software version 8.0 (BDTM, Ashland, OR, USA). The analysis was conducted in quadruplicate, and the results were expressed as mean ± the standard deviation. Statistical comparisons between groups (control vs. treatment) were performed using Mann-Whitney U tests; p < 0.05 was considered statistically significant.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}