
Post-Translational Modifications of Extracellular Proteasome

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
3. Material and Methods
3.1. Cell Culture Conditions and Cell Viability Assays
3.2. Affinity Purification of Proteasome
3.3. Assay of Proteasome Proteolytic Activity
3.4. MALDI FT-ICR Mass Spectrometry
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Konstantinova, I.M.; Tsimokha, A.S.; Mittenberg, A.G. Role of proteasomes in cellular regulation. Int. Rev. Cell Mol. Biol. 2008, 267, 59–124. [Google Scholar]
- Glickman, M.H.; Rubin, D.M.; Coux, O.; Wefes, I.; Pfeifer, G.; Cjeka, Z.; Baumeister, W.; Fried, V.A.; Finley, D. A subcomplex of the proteasome regulatory particle required for ubiquitin-conjugate degradation and related to the COP9-signalosome and eIF3. Cell 1998, 94, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef]
- Groll, M.; Ditzel, L.; Lowe, J.; Stock, D.; Bochtler, M.; Bartunik, H.D.; Huber, R. Structure of 20S proteasome from yeast at 2.4 A resolution. Nature 1997, 386, 463–471. [Google Scholar] [CrossRef]
- Wada, M.; Kosaka, M.; Saito, S.; Sano, T.; Tanaka, K.; Ichihara, A. Serum concentration and localization in tumor cells of proteasomes in patients with hematologic malignancy and their pathophysiologic significance. J. Lab. Clin. Med. 1993, 121, 215–223. [Google Scholar]
- Mueller, O.; Anlasik, T.; Wiedemann, J.; Thomassen, J.; Wohlschlaeger, J.; Hagel, V.; Keyvani, K.; Schwieger, I.; Dahlmann, B.; Sure, U.; et al. Circulating extracellular proteasome in the cerebrospinal fluid: A study on concentration and proteolytic activity. J. Mol. Neurosci. MN 2012, 46, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Sixt, S.U.; Beiderlinden, M.; Jennissen, H.P.; Peters, J. Extracellular proteasome in the human alveolar space: A new housekeeping enzyme? Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L1280–L1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diakonov, E.E.; Selenina, A.V.; Tomilin, A.N.; Tsimokha, A.S. Evidences against vesicle-dependent trafficking and involvement of extracellular proteasomes into cell-to-cell communications. Biochem. Biophys. Res. Commun. 2019, 508, 368–373. [Google Scholar] [CrossRef]
- Kulichkova, V.A.; Artamonova, T.O.; Lyublinskaya, O.G.; Khodorkovskii, M.A.; Tomilin, A.N.; Tsimokha, A.S. Proteomic analysis of affinity-purified extracellular proteasomes reveals exclusively 20S complexes. Oncotarget 2017, 8, 102134–102149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsimokha, A.S.; Zaykova, J.J.; Bottrill, A.; Barlev, N.A. Extracellular Proteasomes Are Deficient in 19S Subunits as Revealed by iTRAQ Quantitative Proteomics. J. Cell. Physiol. 2017, 232, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Zoeger, A.; Blau, M.; Egerer, K.; Feist, E.; Dahlmann, B. Circulating proteasomes are functional and have a subtype pattern distinct from 20S proteasomes in major blood cells. Clin. Chem. 2006, 52, 2079–2086. [Google Scholar] [CrossRef]
- Dutaud, D.; Aubry, L.; Henry, L.; Levieux, D.; Hendil, K.B.; Kuehn, L.; Bureau, J.P.; Ouali, A. Development and evaluation of a sandwich ELISA for quantification of the 20S proteasome in human plasma. J. Immunol. Methods 2002, 260, 183–193. [Google Scholar] [CrossRef]
- Lavabre-Bertrand, T.; Henry, L.; Carillo, S.; Guiraud, I.; Ouali, A.; Dutaud, D.; Aubry, L.; Rossi, J.F.; Bureau, J.P. Plasma proteasome level is a potential marker in patients with solid tumors and hemopoietic malignancies. Cancer 2001, 92, 2493–2500. [Google Scholar] [CrossRef]
- Stoebner, P.E.; Lavabre-Bertrand, T.; Henry, L.; Guiraud, I.; Carillo, S.; Dandurand, M.; Joujoux, J.M.; Bureau, J.P.; Meunier, L. High plasma proteasome levels are detected in patients with metastatic malignant melanoma. Br. J. Dermatol. 2005, 152, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Egerer, K.; Kuckelkorn, U.; Rudolph, P.E.; Ruckert, J.C.; Dorner, T.; Burmester, G.R.; Kloetzel, P.M.; Feist, E. Circulating proteasomes are markers of cell damage and immunologic activity in autoimmune diseases. J. Rheumatol. 2002, 29, 2045–2052. [Google Scholar] [PubMed]
- Heubner, M.; Wimberger, P.; Dahlmann, B.; Kasimir-Bauer, S.; Kimmig, R.; Peters, J.; Wohlschlaeger, J.; Sixt, S.U. The prognostic impact of circulating proteasome concentrations in patients with epithelial ovarian cancer. Gynecol. Oncol. 2011, 120, 233–238. [Google Scholar] [CrossRef]
- Jakob, C.; Egerer, K.; Liebisch, P.; Turkmen, S.; Zavrski, I.; Kuckelkorn, U.; Heider, U.; Kaiser, M.; Fleissner, C.; Sterz, J.; et al. Circulating proteasome levels are an independent prognostic factor for survival in multiple myeloma. Blood 2007, 109, 2100–2105. [Google Scholar] [CrossRef] [Green Version]
- Sixt, S.U.; Dahlmann, B. Extracellular, circulating proteasomes and ubiquitin - incidence and relevance. Biochim. Biophys. Acta 2008, 1782, 817–823. [Google Scholar] [CrossRef] [Green Version]
- Bochmann, I.; Ebstein, F.; Lehmann, A.; Wohlschlaeger, J.; Sixt, S.U.; Kloetzel, P.M.; Dahlmann, B. T lymphocytes export proteasomes by way of microparticles: A possible mechanism for generation of extracellular proteasomes. J. Cell. Mol. Med. 2014, 18, 59–68. [Google Scholar] [CrossRef]
- Dieude, M.; Bell, C.; Turgeon, J.; Beillevaire, D.; Pomerleau, L.; Yang, B.; Hamelin, K.; Qi, S.; Pallet, N.; Beland, C.; et al. The 20S proteasome core, active within apoptotic exosome-like vesicles, induces autoantibody production and accelerates rejection. Sci. Transl. Med. 2015, 7, 318ra200. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Chen, X.; Pan, Q.; Wang, Y.; Su, S.; Jiang, C.; Li, Y.; Xu, N.; Wu, L.; Lou, X.; et al. A Comprehensive Proteomics Analysis Reveals a Secretory Path- and Status-Dependent Signature of Exosomes Released from Tumor-Associated Macrophages. J. Proteome Res. 2015, 14, 4319–4331. [Google Scholar] [CrossRef] [PubMed]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Thery, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drews, O.; Wildgruber, R.; Zong, C.; Sukop, U.; Nissum, M.; Weber, G.; Gomes, A.V.; Ping, P. Mammalian proteasome subpopulations with distinct molecular compositions and proteolytic activities. Mol. Cell. Proteom. 2007, 6, 2021–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhakrishnan, S.K.; Lee, C.S.; Young, P.; Beskow, A.; Chan, J.Y.; Deshaies, R.J. Transcription factor Nrf1 mediates the proteasome recovery pathway after proteasome inhibition in mammalian cells. Mol. Cell 2010, 38, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Nissan, G.; Sharon, M. Regulating the 20S proteasome ubiquitin-independent degradation pathway. Biomolecules 2014, 4, 862–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, H.; Kimura, Y.; Kimura, A. Biological significance of co- and post-translational modifications of the yeast 26S proteasome. J. Proteom. 2016, 134, 37–46. [Google Scholar] [CrossRef]
- Kors, S.; Geijtenbeek, K.; Reits, E.; Schipper-Krom, S.J.F. Regulation of proteasome activity by (post-) transcriptional mechanisms. Front. Mol. Biosci. 2019, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.M.; Netto, L.E.; Simoes, V.; Santos, L.F.; Gozzo, F.C.; Demasi, M.A.; Oliveira, C.L.; Bicev, R.N.; Klitzke, C.F.; Sogayar, M.C.; et al. Redox control of 20S proteasome gating. Antioxid. Redox Signal. 2012, 16, 1183–1194. [Google Scholar] [CrossRef] [Green Version]
- Bose, S.; Stratford, F.L.; Broadfoot, K.I.; Mason, G.G.; Rivett, A.J. Phosphorylation of 20S proteasome alpha subunit C8 (alpha7) stabilizes the 26S proteasome and plays a role in the regulation of proteasome complexes by gamma-interferon. Biochem. J. 2004, 378 Pt 1, 177–184. [Google Scholar] [CrossRef]
- Satoh, K.; Sasajima, H.; Nyoumura, K.I.; Yokosawa, H.; Sawada, H. Assembly of the 26S proteasome is regulated by phosphorylation of the p45/Rpt6 ATPase subunit. Biochemistry 2001, 40, 314–319. [Google Scholar]
- Yokota, N.; Kataoka, Y.; Hashii, N.; Kawasaki, N.; Sawada, H. Sperm-specific C-terminal processing of the proteasome PSMA1/alpha6 subunit. Biochem. Biophys. Res. Commun. 2011, 410, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Bingol, B.; Wang, C.F.; Arnott, D.; Cheng, D.; Peng, J.; Sheng, M. Autophosphorylated CaMKIIalpha acts as a scaffold to recruit proteasomes to dendritic spines. Cell 2010, 140, 567–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, S.; Mason, G.G.; Rivett, A.J. Phosphorylation of proteasomes in mammalian cells. Mol. Biol. Rep. 1999, 26, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Kato, Y.; Hirano, H. N-myristoylation of the Rpt2 subunit regulates intracellular localization of the yeast 26S proteasome. Biochemistry 2012, 51, 8856–8866. [Google Scholar] [PubMed]
- Moreno-Gonzalo, O.; Fernandez-Delgado, I.; Sanchez-Madrid, F. Post-translational add-ons mark the path in exosomal protein sorting. Cell. Mol. Life Sci. 2018, 75, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artamonova, T.O.; Khodorkovskii, M.A.; Tsimokha, A.S. Mass spectrometric analysis of affinity-purified proteasomes from the human myelogenous leukemia K562 cell line. Russ. J. Bioorganic Chem. 2014, 40, 666–680. [Google Scholar] [CrossRef]
- Moiseeva, T.N.; Fedorova, O.A.; Tsimokha, A.S.; Mittenberg, A.G.; Barlev, N.A. Effect of ubiquitination on peptidase activities of proteasomes in genotoxic stress. Dokl. Biochem. Biophys. 2010, 435, 307–311. [Google Scholar] [CrossRef]
- Guo, X.; Huang, X.; Chen, M.J. Reversible phosphorylation of the 26S proteasome. Protein Cell 2017, 8, 255–272. [Google Scholar] [CrossRef] [Green Version]
- Zong, N.; Ping, P.; Lau, E.; Choi, H.J.; Ng, D.C.; Meyer, D.; Fang, C.; Li, H.; Wang, D.; Zelaya, I.M.; et al. Lysine ubiquitination and acetylation of human cardiac 20S proteasomes. Proteom. Clin. Appl. 2014, 8, 590–594. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, J.; Iwafune, Y.; Akiyama, T.; Okayama, A.; Nakamura, H.; Arakawa, N.; Kimura, Y.; Hirano, H. Co- and post-translational modifications of the 26S proteasome in yeast. Proteomics 2010, 10, 2769–2779. [Google Scholar] [CrossRef]
- Iwafune, Y.; Kawasaki, H.; Hirano, H. Electrophoretic analysis of phosphorylation of the yeast 20S proteasome. Electrophoresis 2002, 23, 329–338. [Google Scholar] [CrossRef]
- Jarome, T.J.; Helmstetter, F.J. The ubiquitin-proteasome system as a critical regulator of synaptic plasticity and long-term memory formation. Neurobiol. Learn. Mem. 2013, 105, 107–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Zong, C.; Wang, Y.; Young, G.W.; Deng, N.; Souda, P.; Li, X.; Whitelegge, J.; Drews, O.; Yang, P.Y.; et al. Revealing the dynamics of the 20 S proteasome phosphoproteome: A combined CID and electron transfer dissociation approach. Mol. Cell. Proteom. MCP 2008, 7, 2073–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Hu, Y.; Huang, P.; Toleman, C.A.; Paterson, A.J.; Kudlow, J.E. Proteasome function is regulated by cyclic AMP-dependent protein kinase through phosphorylation of Rpt6. J. Biol. Chem. 2007, 282, 22460–22471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsimokha, A.S.; Mittenberg, A.G.; Kulichkova, V.A.; Kozhukharova, I.V.; Gause, L.N.; Konstantinova, I.M. Changes in composition and activities of 26S proteasomes under the action of doxorubicin--apoptosis inductor of erythroleukemic K562 cells. Cell Biol. Int. 2007, 31, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Kulichkova, V.A.; Tsimokha, A.S.; Fedorova, O.A.; Moiseeva, T.N.; Bottril, A.; Lezina, L.; Gauze, L.N.; Konstantinova, I.M.; Mittenberg, A.G.; Barlev, N.A. 26S proteasome exhibits endoribonuclease activity controlled by extra-cellular stimuli. Cell Cycle 2010, 9, 840–849. [Google Scholar] [CrossRef]
- Liu, X.; Huang, W.; Li, C.; Li, P.; Yuan, J.; Li, X.; Qiu, X.B.; Ma, Q.; Cao, C. Interaction between c-Abl and Arg tyrosine kinases and proteasome subunit PSMA7 regulates proteasome degradation. Mol. Cell 2006, 22, 317–327. [Google Scholar] [CrossRef]
- Li, D.; Dong, Q.; Tao, Q.; Gu, J.; Cui, Y.; Jiang, X.; Yuan, J.; Li, W.; Xu, R.; Jin, Y. c-Abl regulates proteasome abundance by controlling the ubiquitin-proteasomal degradation of PSMA7 subunit. Cell Rep. 2015, 10, 484–496. [Google Scholar] [CrossRef] [Green Version]
- Rape, M. Ubiquitylation at the crossroads of development and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 59–70. [Google Scholar] [CrossRef]
- Isasa, M.; Katz, E.J.; Kim, W.; Yugo, V.; Gonzalez, S.; Kirkpatrick, D.S.; Thomson, T.M.; Finley, D.; Gygi, S.P.; Crosas, B. Monoubiquitination of RPN10 regulates substrate recruitment to the proteasome. Mol. Cell 2010, 38, 733–745. [Google Scholar] [CrossRef] [Green Version]
- Besche, H.C.; Sha, Z.; Kukushkin, N.V.; Peth, A.; Hock, E.M.; Kim, W.; Gygi, S.; Gutierrez, J.A.; Liao, H.; Dick, L.; et al. Autoubiquitination of the 26S proteasome on Rpn13 regulates breakdown of ubiquitin conjugates. EMBO J. 2014, 33, 1159–1176. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, S.M.; Neslund-Dudas, C.; Shen, M.; Cui, C.; Mitra, B.; Dou, Q.P. Involvement of ALAD-20S Proteasome Complexes in Ubiquitination and Acetylation of Proteasomal α2 Subunits. J. Cell. Biochem. 2016, 117, 144–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinert, B.T.; Scholz, C.; Wagner, S.A.; Iesmantavicius, V.; Su, D.; Daniel, J.A.; Choudhary, C. Lysine succinylation is a frequently occurring modification in prokaryotes and eukaryotes and extensively overlaps with acetylation. Cell Rep. 2013, 4, 842–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, Y.; Takaoka, M.; Tanaka, S.; Sassa, H.; Tanaka, K.; Polevoda, B.; Sherman, F.; Hirano, H. N(alpha)-acetylation and proteolytic activity of the yeast 20 S proteasome. J. Biol. Chem. 2000, 275, 4635–4639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groll, M.; Bajorek, M.; Köhler, A.; Moroder, L.; Rubin, D.M.; Huber, R.; Glickman, M.H.; Finley, D. A gated channel into the proteasome core particle. Nat. Struct. Biol. 2000, 7, 1062–1067. [Google Scholar] [CrossRef]
- Kimura, Y.; Saeki, Y.; Yokosawa, H.; Polevoda, B.; Sherman, F.; Hirano, H. N-Terminal modifications of the 19S regulatory particle subunits of the yeast proteasome. Arch. Biochem. Biophys. 2003, 409, 341–348. [Google Scholar]
- Wang, D.; Fang, C.; Zong, N.C.; Liem, D.A.; Cadeiras, M.; Scruggs, S.B.; Yu, H.; Kim, A.K.; Yang, P.; Deng, M.; et al. Regulation of acetylation restores proteolytic function of diseased myocardium in mouse and human. Mol. Cell. Proteom. MCP 2013, 12, 3793–3802. [Google Scholar] [CrossRef] [Green Version]
- Beltrao, P.; Albanèse, V.; Kenner, L.R.; Swaney, D.L.; Burlingame, A.; Villén, J.; Lim, W.A.; Fraser, J.S.; Frydman, J.; Krogan, N.J. Systematic functional prioritization of protein posttranslational modifications. Cell 2012, 150, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Landry, C.R.; Levy, E.D.; Michnick, S.W. Weak functional constraints on phosphoproteomes. Trends Genet. 2009, 25, 193–197. [Google Scholar] [CrossRef]
- Nguyen Ba, A.N.; Moses, A.M. Evolution of characterized phosphorylation sites in budding yeast. Mol. Biol. Evol. 2010, 27, 2027–2037. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, C.F.; Baker, P.R.; Chen, P.L.; Kaiser, P.; Huang, L. Mass spectrometric characterization of the affinity-purified human 26S proteasome complex. Biochemistry 2007, 46, 3553–3565. [Google Scholar] [PubMed]
- Kulichkova, V.A.; Artamonova, T.O.; Zaykova, J.J.; Ermolaeva, J.B.; Khodorkovskii, M.A.; Barlev, N.A.; Tomilin, A.N.; Tsimokha, A.S. Simultaneous EGFP and Tag Labeling of the beta7 Subunit for Live Imaging and Affinity Purification of Functional Human Proteasomes. Mol. Biotechnol. 2015, 57, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Tsimokha, A.S.; Kulichkova, V.A.; Karpova, E.V.; Zaykova, J.J.; Aksenov, N.D.; Vasilishina, A.A.; Kropotov, A.V.; Antonov, A.; Barlev, N.A. DNA damage modulates interactions between microRNAs and the 26S proteasome. Oncotarget 2014, 5, 3555–3567. [Google Scholar] [CrossRef] [Green Version]
- Jerebtsova, M.; Klotchenko, S.A.; Artamonova, T.O.; Ammosova, T.; Washington, K.; Egorov, V.V.; Shaldzhyan, A.A.; Sergeeva, M.V.; Zatulovskiy, E.A.; Temkina, O.A.; et al. Mass spectrometry and biochemical analysis of RNA polymerase II: Targeting by protein phosphatase-1. Mol. Cell. Biochem. 2011, 347, 79–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
{kind=link}
| Proteasome | Peptide | Start | End | |
|---|---|---|---|---|
| Cellular | Extracellular | |||
| α1 (PSMA6) | ||||
| K45-ac | (R)GKDCAVIVTQK(K) | 44 | 54 | |
| T52-p | (R)GKDCAVIVTQK(K) | 44 | 54 | |
| (R)GKDCAVIVTQKK(V) | 44 | 55 | ||
| K54-ac | (R)GKDCAVIVTQK(K) | 44 | 54 | |
| T231-p | (R)ILTEAEIDAHLVALAERD(-) | 229 | 246 | |
| α2 (PSMA2) | ||||
| T48-p | (K)AANGVVLATEKK(Q) | 40 | 51 | |
| K53-ac | (K)QKSILYDER(S) | 52 | 60 | |
| K70-ub | (K)VEPITKHIGLVYSGMGPDYR(V) | 65 | 84 | |
| K227-ub | (R)RLTPTEVKDYLAAIA(-) | 220 | 234 | |
| K227-ac | (R)LTPTEVKDYLAAIA(-) | 221 | 234 | |
| α3 (PSMA4) | ||||
| S188-p | (K)SALALAIK(V) | 188 | 195 | |
| K195-ac | (K)SALALAIK(V) | 188 | 195 | |
| K205-ac * | (K)TMDVSKLSAEK(V) | 200 | 210 | |
| K210-ac | (K)TMDVSKLSAEK(V) | 200 | 210 | |
| (K)LSAEK(V) | 206 | 210 | ||
| S207-p | (K)LSAEK(V) | 206 | 210 | |
| K222-ac | (K)VEIATLTRENGK(T) | 211 | 222 | |
| (K)VEIATLTRENGKTVIR(V) | 211 | 226 | ||
| K222-ub | (K)VEIATLTRENGKTVIR(V) | 211 | 226 | |
| K222-sc | (R)ENGKTVIR(V) | 219 | 226 | |
| α4 (PSMA7) | ||||
| S49-p | (K)KSVAKLQDER(T) | 48 | 57 | |
| K52-ub | (K)SVAKLQDER(T) | 49 | 57 | |
| K52-sc | (K)SVAKLQDERTVR(K) | 49 | 60 | |
| S167-p | (K)ANAIGRGAKSVREFLEK(N) | 158 | 174 | |
| S201-p | (K)ALLEVVQSGGK(N) | 194 | 204 | |
| Y228-p | (K)YVAEIEKEKEENEK(K) | 228 | 241 | |
| α5 (PSMA5) | ||||
| S79-p | (K)IVEIDAHIGCAMSGLIADAK(T) | 67 | 86 | |
| S197-p | (K)EAIKSSLIILKQVMEEK(L) | 193 | 209 | |
| (K)SSLIILKQVMEEK(L) | 197 | 209 | ||
| S198-p * | (K)EAIKSSLIILKQVMEEK(L) | 193 | 209 | |
| (K)SSLIILKQVMEEK(L) | 197 | 209 | ||
| K239-ac * | (K)EELEEVIKDI(-) | 232 | 241 | |
| α6 (PSMA1) | ||||
| S40-p | (K)SKTHAVLVALKR(A) | 40 | 51 | |
| K41-ac | (K)SKTHAVLVALKR(A) | 40 | 51 | |
| K41-sc | (K)SKTHAVLVALKR(A) | 40 | 51 | |
| T42-p | (K)SKTHAVLVALKR(A) | 40 | 51 | |
| K50-ac | (K)SKTHAVLVALKR(A) | 40 | 51 | |
| K50-sc | (K)SKTHAVLVALKR(A) | 40 | 51 | |
| S54-p | (K)RAQSELAAHQKK(I) | 51 | 62 | |
| K62-sc | (K)KILHVDNHIGISIAGLTADAR(L) | 62 | 82 | |
| α7 (PSMA3) | ||||
| K29-ac | (R)VFQVEYAMK(A) | 21 | 29 | |
| K43-ac | (K)AVENSSTAIGIRCK(D) | 30 | 43 | |
| S56-p | (R)CKDGVVFGVEKLVLSK(L) | 42 | 57 | |
| (K)LVLSK(L) | 53 | 57 | ||
| K57-ac | (K)LVLSK(L) | 53 | 57 | |
| K65-ac | (K)LYEEGSNKR(L) | 58 | 66 | |
| S87-p | (R)SLADIAR(E) | 87 | 93 | |
| T175-p | (R)QAAKTEIEKLQMK(E) | 171 | 183 | |
| T186-p | (R)QAAKTEIEKLQMK(E) | 171 | 183 | |
| (K)EMTCRDIVKEVAK(I) | 184 | 196 | ||
| K192-ac * | (K)EMTCRDIVK(E) | 184 | 192 | |
| K208-sc | (K)DKAFELELSWVGELTNGR(H) | 207 | 224 | |
| K230-ac | (R)HEIVPK(D) | 225 | 230 | |
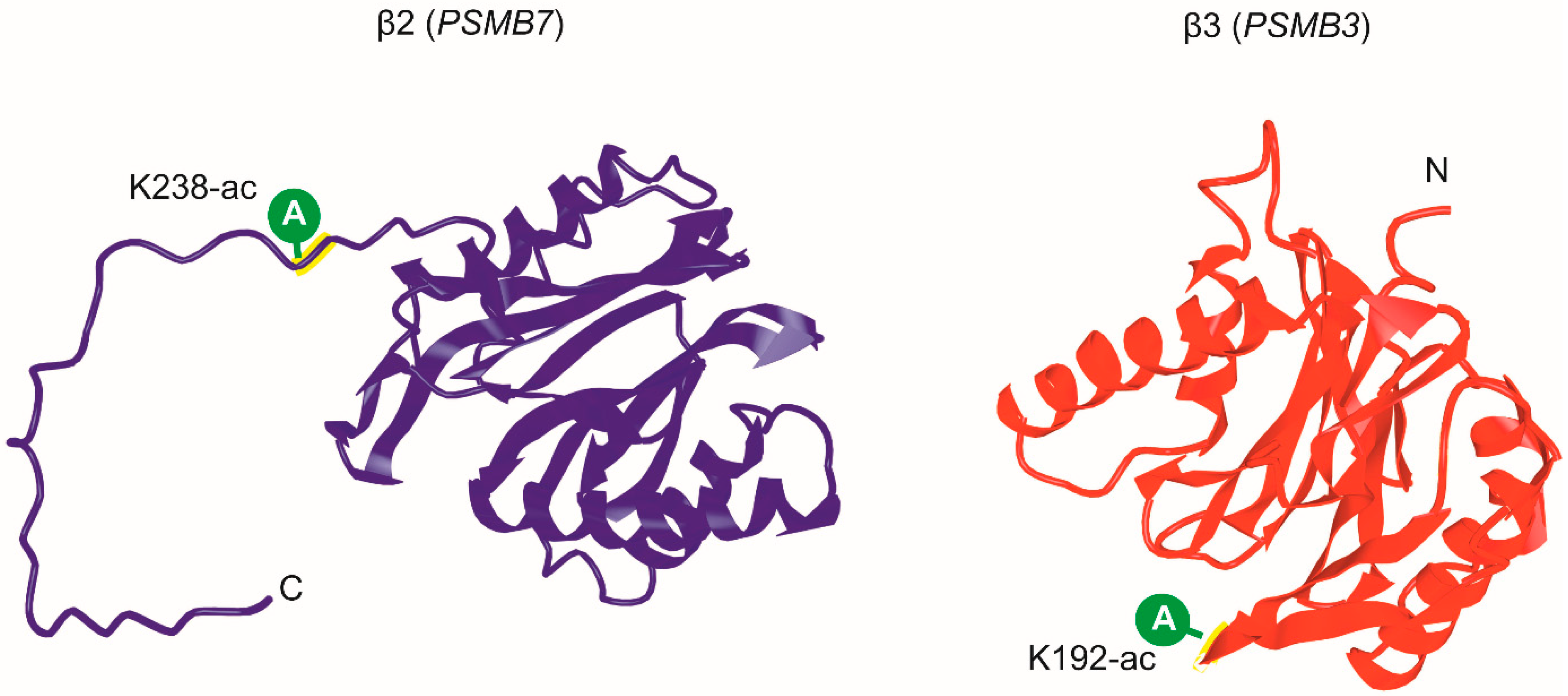
| β2 (PSMB7) | ||||
| K31-ub | (R)NAVLEADFAKR(G) | 22 | 32 | |
| Y34-p | (K)RGYKLPK(V) | 32 | 38 | |
| (R)GYKLPKVR(K) | 33 | 40 | ||
| K35-ac | (R)GYKLPKVR(K) | 33 | 40 | |
| K38-ac | (R)GYKLPKVR(K) | 33 | 40 | |
| T61-p | (K)DGIVLGADTRATEGMVVADK(N) | 53 | 72 | |
| T64-p | (K)DGIVLGADTRATEGMVVADK(N) | 53 | 72 | |
| K72-ac | (K)DGIVLGADTRATEGMVVADK(N) | 53 | 72 | |
| T121-p * | (R)VVTANRMLK(Q) | 119 | 127 | |
| K225-ac | (K)NKLDFLRPYTVPNKK(G) | 224 | 238 | |
| Y232-p | (K)NKLDFLRPYTVPNKK(G) | 224 | 238 | |
| T233-p * | (K)NKLDFLRPYTVPNKK(G) | 224 | 238 | |
| K237-ac * | (K)NKLDFLRPYTVPNKK(G) | 224 | 238 | |
| (K)LDFLRPYTVPNK(K) | 226 | 237 | ||
| (K)LDFLRPYTVPNKKGTR(L) | 226 | 241 | ||
| K238-ac | (K)NKLDFLRPYTVPNKK(G) | 224 | 238 | |
| (K)LDFLRPYTVPNKKGTR(L) | 226 | 241 | ||
| T240-p | (K)GTRLGR(Y) | 239 | 244 | |
| β3 (PSMB3) | ||||
| K41-ac | (R)RFGIQAQMVTTDFQK(I) | 27 | 41 | |
| K115-ac | (K)RFGPYYTEPVIAGLDPK(T) | 99 | 115 | |
| K192-ac | (R)DAVSGMGVIVHIIEK(D) | 178 | 192 | |
| S181-p * | (R)DAVSGMGVIVHIIEKDK(I) | 178 | 194 | |
| K201-ub | (R)TLKARMD(-) | 199 | 205 | |
| β4 (PSMB2) | ||||
| S23-p | (R)VAASNIVQMKDDHDKMFK(M) | 20 | 37 | |
| K29-ac * | (R)VAASNIVQMKDDHDKMFK(M) | 20 | 37 | |
| K34-ac | (R)VAASNIVQMKDDHDKMFK(M) | 20 | 37 | |
| (K)DDHDKMFK(M) | 30 | 37 | ||
| K37-ac | (R)VAASNIVQMKDDHDKMFK(M) | 20 | 37 | |
| (K)DDHDKMFK(M) | 30 | 37 | ||
| K198-ub | (K)NGIHDLDNISFPKQGS(-) | 186 | 201 | |
| β5 (PSMB5) | ||||
| K130-ub | (R)IYELRNKER(I) | 124 | 132 | |
| K130-ac | (R)NKERISVAAASK(L) | 129 | 140 | |
| S134-p | (R)NKERISVAAASK(L) | 129 | 140 | |
| S139-p | (R)NKERISVAAASK(L) | 129 | 140 | |
| K140-ac | (R)NKERISVAAASK(L) | 129 | 140 | |
| β6 (PSMB1) | ||||
| K70-ac | (R)LSEGFSIHTRDSPK(C) | 57 | 70 | |
| Y103-p | (K)IIEARLKMYK(H) | 95 | 104 | |
| S209-p | (R)AMRLVKDVFISAAER(D) | 199 | 213 | |
| (R)LVKDVFISAAER(D) | 202 | 213 | ||
| K204-ac | (R)LVKDVFISAAER(D) | 202 | 213 | |
| K228-sc | (R)DVYTGDALRICIVTKEGIR(E) | 214 | 232 | |
| K228-ac | (R)ICIVTKEGIREETVSLR(K) | 223 | 239 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsimokha, A.S.; Artamonova, T.O.; Diakonov, E.E.; Khodorkovskii, M.A.; Tomilin, A.N. Post-Translational Modifications of Extracellular Proteasome. Molecules 2020, 25, 3504. https://doi.org/10.3390/molecules25153504
Tsimokha AS, Artamonova TO, Diakonov EE, Khodorkovskii MA, Tomilin AN. Post-Translational Modifications of Extracellular Proteasome. Molecules. 2020; 25(15):3504. https://doi.org/10.3390/molecules25153504
Chicago/Turabian StyleTsimokha, Anna S., Tatiana O. Artamonova, Egor E. Diakonov, Mikhail A. Khodorkovskii, and Alexey N. Tomilin. 2020. "Post-Translational Modifications of Extracellular Proteasome" Molecules 25, no. 15: 3504. https://doi.org/10.3390/molecules25153504