
A Natural Flavone Tricin from Grains Can Alleviate Tumor Growth and Lung Metastasis in Colorectal Tumor Mice

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
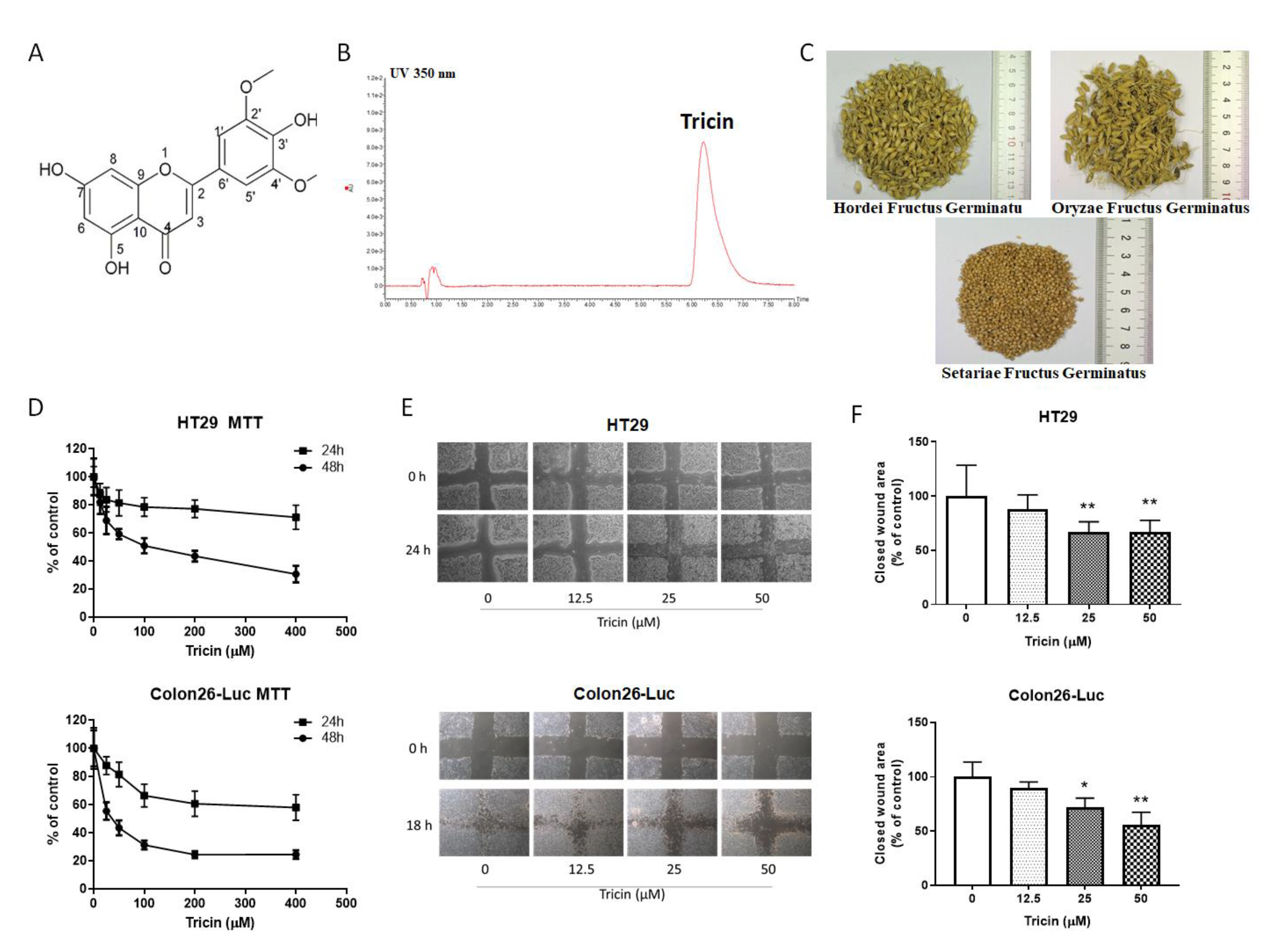
2.1. Quantification of Tricin in Medicinal Grains
2.2. Tricin Reduced Cell Viability and Migration of Human and Mouse Colon Cancer Cells
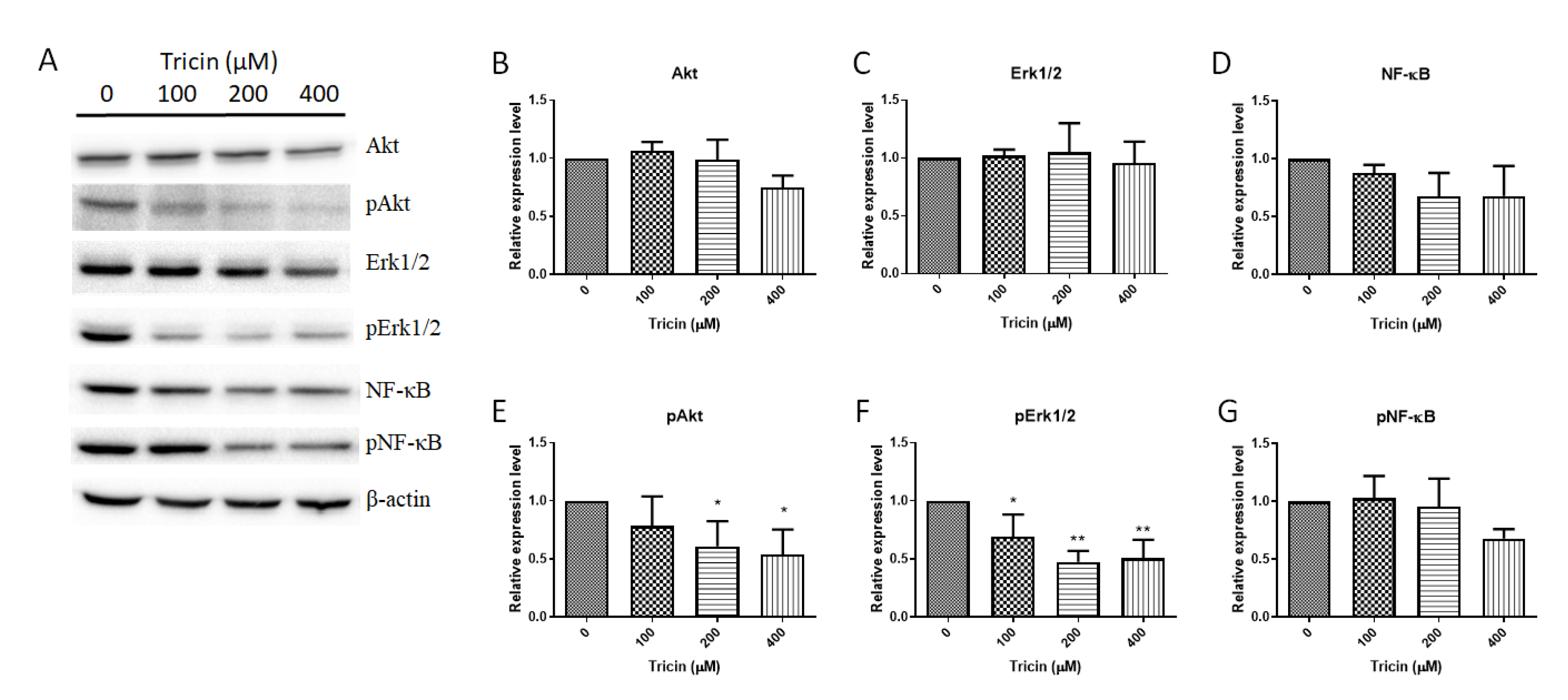
2.3. Tricin Affected Akt, Erk1/2 and NF-κB signaling Pathways in HT-29 Cells
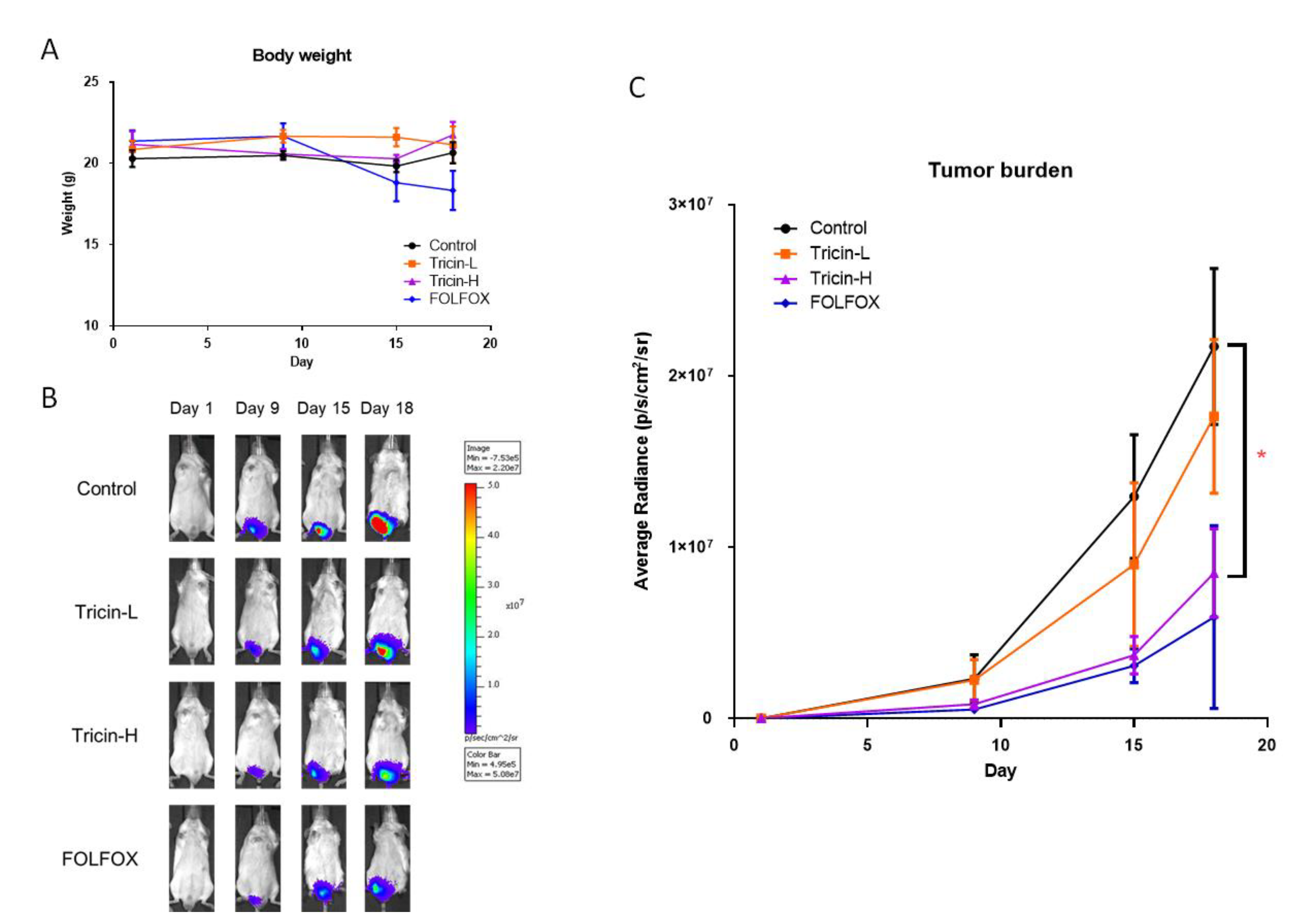
2.4. Tricin Suppressed Mouse Orthotopic Colon Tumor Growth
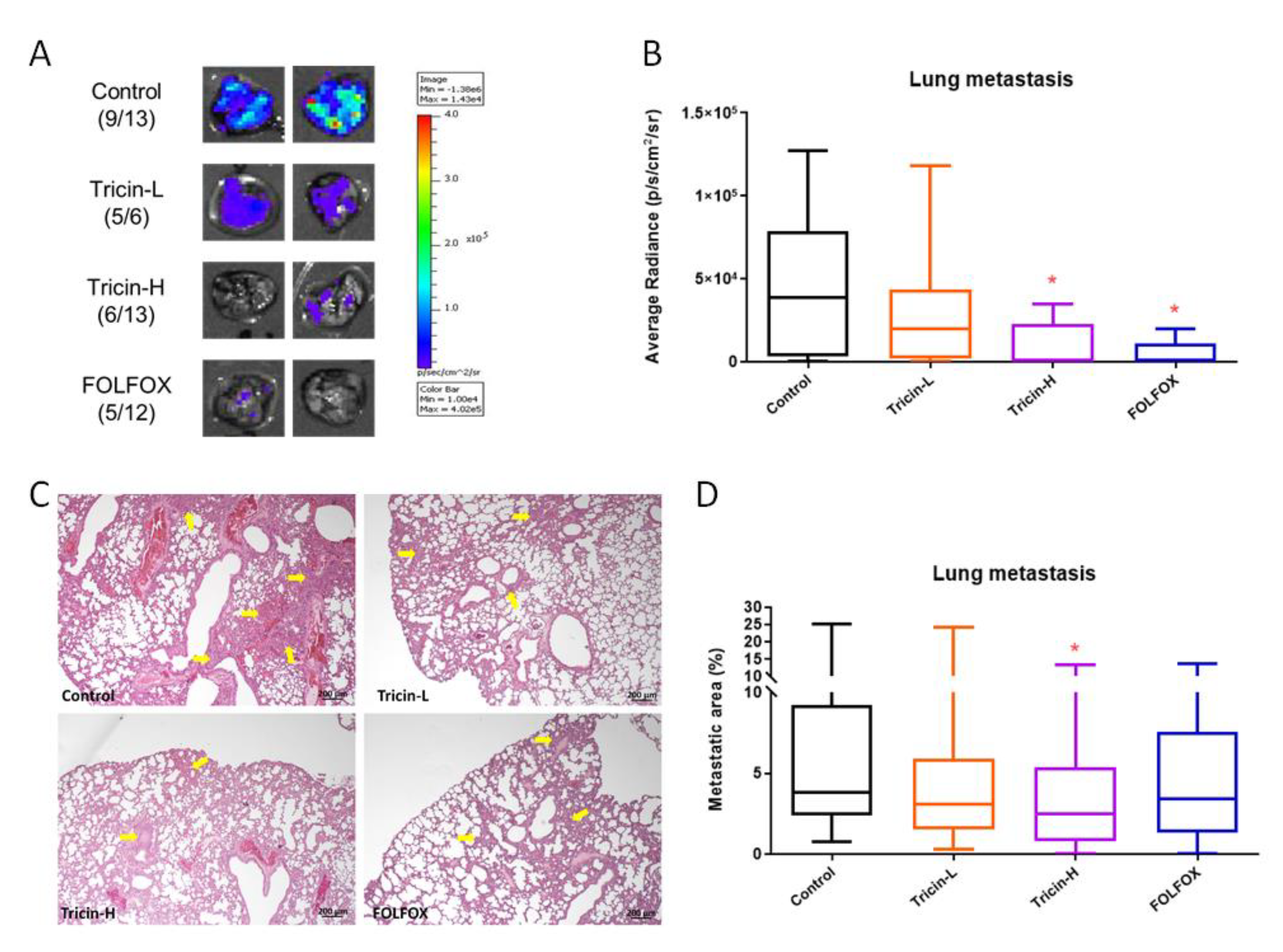
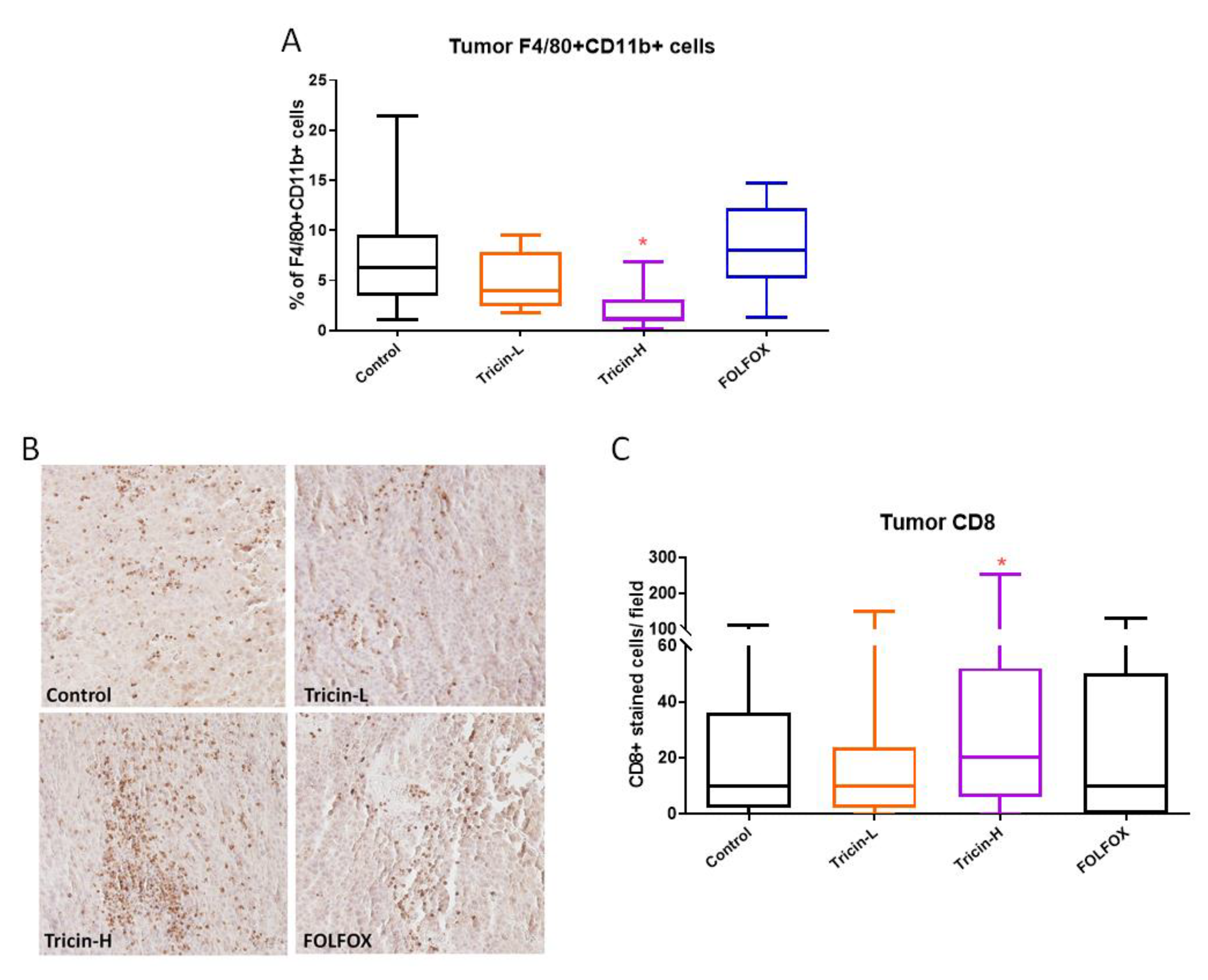
2.5. Tricin Reduced Orthotopic Colon Tumor Metastasis
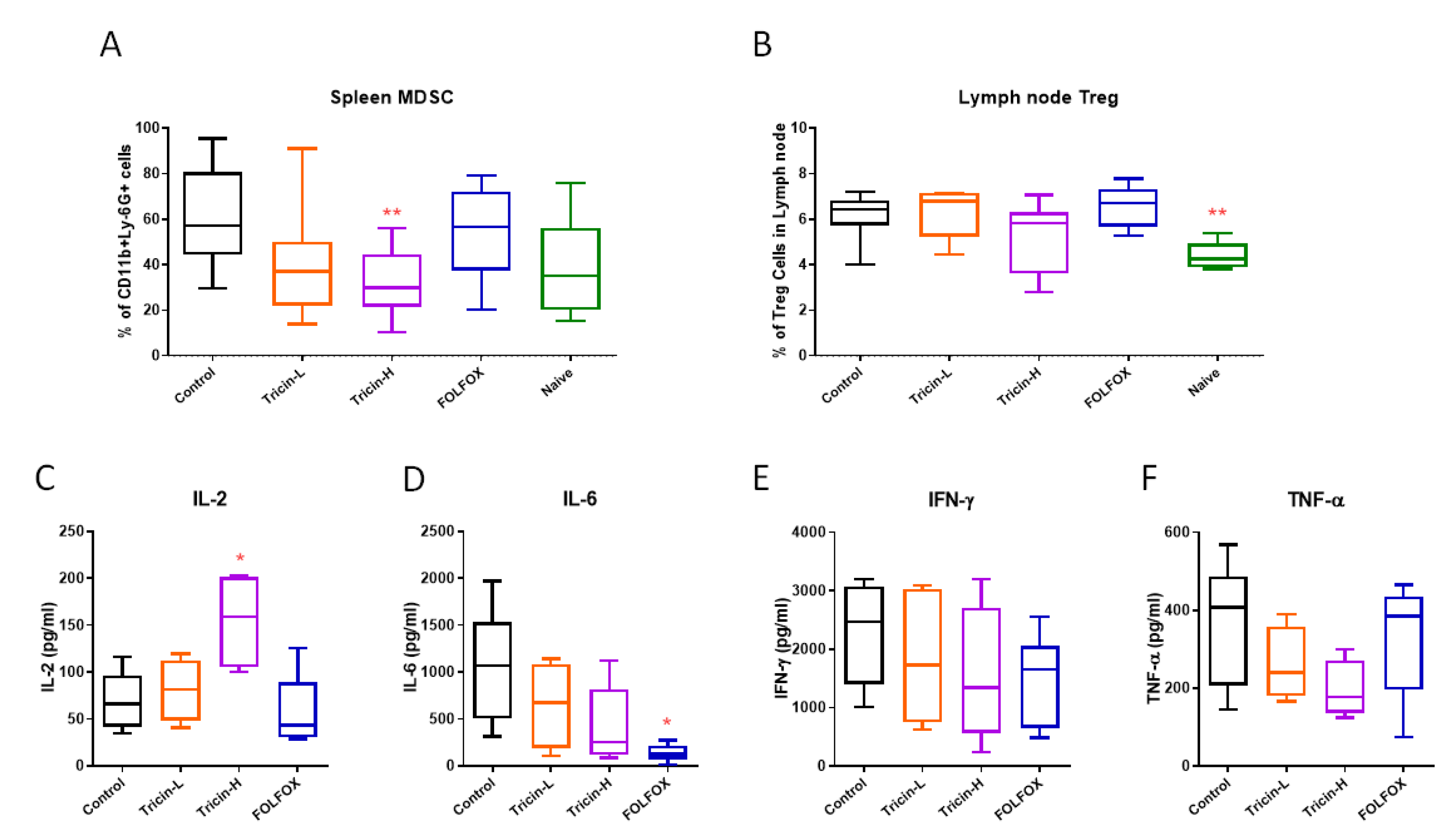
2.6. Immune Cell Populations of Syngeneic Colon Tumor-Bearing Mice Altered by Tricin Treatment
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. UPLC Analysis of Tricin in Medicinal Grains
4.3. Cell Culture and Viability Assay
4.4. Scratch Wound Healing and Cell Migration Assays
4.5. Western Blotting of HT-29 Cells
4.6. Syngeneic Orthotopic Colon Tumor-Bearing Mouse Model
4.7. Flow Cytometry for Immune Cells Characterization and ELISA for Cytokines Determination
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Tabung, F.K.; Brown, L.S.; Fung, T.T. Dietary patterns and colorectal cancer risk: A review of 17 Years of evidence (2000-2016). Curr. Colorect. Canc. Rep. 2017, 13, 440–454. [Google Scholar] [CrossRef]
- Theodoratou, E.; Timofeeva, M.; Li, X.; Meng, X.R.; Ioannidis, J.P.A. Nature, nurture, and cancer risks: Genetic and nutritional contributions to cancer. Annu. Rev. Nutr. 2017, 37, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, L.E.; Taddeo, S.S.; Weeks, B.R.; Carroll, R.J.; Dykes, L.; Rooney, L.W.; Turner, N.D. Impact of novel sorghum bran diets on DSS-induced colitis. Nutrients 2017, 9, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, S.H.; Shi, J.Y.; Li, Z.; Gao, H.X.; Shi, T.L.; Li, Z.W.; Li, Z.Y. Targeted anti-colon cancer activities of a millet bran-derived peroxidase were mediated by elevated ROS generation. Food Funct. 2015, 6, 2331–2338. [Google Scholar] [CrossRef] [PubMed]
- Forster, G.M.; Raina, K.; Kumar, A.; Kumar, S.; Agarwal, R.; Chen, M.H.; Bauer, J.E.; McClung, A.M.; Ryan, E.P. Rice varietal differences in bioactive bran components for inhibition of colorectal cancer cell growth. Food Chem. 2013, 141, 1545–1552. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.V.K.; Maheswaraiah, A.; Naidu, K.A. Rice bran oil and n-3 fatty acid-rich garden cress (Lepidium sativum) seed oil attenuate murine model of ulcerative colitis. Int. J. Colorectal Dis. 2014, 29, 267–269. [Google Scholar] [CrossRef]
- Islam, J.; Koseki, T.; Watanabe, K.; Ardiansyah; Budijanto, S.; Oikawa, A.; Alauddin, M.; Goto, T.; Aso, H.; Komai, M.; et al. Dietary supplementation of fermented rice bran effectively alleviates dextran sodium sulfate-induced colitis in mice. Nutrients 2017, 9, 747. [Google Scholar] [CrossRef]
- Henderson, A.J.; Ollila, C.A.; Kumar, A.; Borresen, E.C.; Raina, K.; Agarwal, R.; Ryan, E.P. Chemopreventive properties of dietary rice bran: Current status and future prospects. Adv. Nutr. 2012, 3, 643–653. [Google Scholar] [CrossRef] [Green Version]
- Hudson, E.A.; Dinh, P.A.; Kokubun, T.; Simmonds, M.S.J.; Gescher, A. Characterization of potentially chemopreventive phenols in extracts of brown rice that inhibit the growth of human breast and colon cancer cells. Cancer Epidem. Biomar. 2000, 9, 1163–1170. [Google Scholar]
- Cai, H.; Al-Fayez, M.; Tunstall, R.G.; Platton, S.; Greaves, P.; Steward, W.P.; Gescher, A.J. The rice bran constituent tricin potently inhibits cyclooxygenase enzymes and interferes with intestinal carcinogenesis in ApcMin mice. Mol. Cancer Ther. 2005, 4, 1287–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, H.; Sale, S.; Britton, R.G.; Brown, K.; Steward, W.P.; Gescher, A.J. Pharmacokinetics in mice and metabolism in murine and human liver fractions of the putative cancer chemopreventive agents 3′,4′,5′,5,7-pentamethoxyflavone and tricin (4′,5,7-trihydroxy-3′,5′-dimethoxyflavone). Cancer Chemoth. Pharm. 2011, 67, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Oyama, T.; Yasui, Y.; Sugie, S.; Koketsu, M.; Watanabe, K.; Tanaka, T. Dietary tricin suppresses inflammation-related colon carcinogenesis in male Crj: CD-1 mice. Cancer Prev. Res. 2009, 2, 1031–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goufo, P.; Trindade, H. Rice antioxidants: Phenolic acids, flavonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, gamma-oryzanol, and phytic acid. Food Sci. Nutr. 2014, 2, 75–104. [Google Scholar] [CrossRef] [PubMed]
- Moheb, A.; Grondin, M.; Ibrahim, R.K.; Roy, R.; Sarhan, F. Winter wheat hull (husk) is a valuable source for tricin, a potential selective cytotoxic agent. Food Chem. 2013, 138, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Poulev, A.; Chen, M.H.; Cherravuru, S.; Raskin, I.; Belanger, F.C. Variation in levels of the flavone tricin in bran from rice genotypes varying in pericarp color. J. Cereal Sci. 2018, 79, 226–232. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2015.
- Chen, W.T.; Ren, J.L.; Jing, L.; Tao, L.J.; Shi, Q.; Hou, F.G. Study on the regularity of traditional Chinese medicine in the treatment of advanced colorectal cancer based on clustering and correlation analysis. Guid. J. Tradit. Chin. Med. Pharm. 2017, 23, 36–40. [Google Scholar]
- Ling, J.H.; Wang, N.; Ren, Y.Z.; Wang, L.H.; Wang, J.H.; Li, X. Determination of tricin in malt by HPLC. Zhong Yao Cai 2005, 36, 1632–1634. [Google Scholar]
- Min, S.J.; Lim, J.Y.; Kim, H.R.; Kim, S.J.; Kim, Y. Sasa quelpaertensis leaf extract inhibits colon cancer by regulating cancer cell stemness in vitro and in vivo. Int. J. Mol. Sci. 2015, 16, 9976–9997. [Google Scholar] [CrossRef] [Green Version]
- Chung, D.J.; Wang, C.J.; Yeh, C.W.; Tseng, T.H. Inhibition of the proliferation and invasion of C6 glioma cells by tricin via the upregulation of focal-adhesion-kinase-targeting microRNA-7. J. Agr. Food Chem. 2018, 66, 6708–6716. [Google Scholar] [CrossRef]
- Shalini, V.; Pushpan, C.K.; Sindhu, G.; Jayalekshmy, A.; Helen, A. Tricin, flavonoid from Njavara reduces inflammatory responses in hPBMCs by modulating the p38MAPK and PI3K/Akt pathways and prevents inflammation associated endothelial dysfunction in HUVECs. Immunobiology 2016, 221, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.; Lu, F.C.; Regner, M.; Zhu, Y.M.; Rencoret, J.; Ralph, S.A.; Zakai, U.I.; Morreel, K.; Boerjan, W.; Ralph, J. Tricin, a flavonoid monomer in monocot lignification. Plant. Physiol. 2015, 167, 1284–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.M.; Ibrahim, R.K. Tricin-a potential multifunctional nutraceutical. Phytochem. Rev. 2010, 9, 413–424. [Google Scholar] [CrossRef]
- Yue, G.G.L.; Lee, J.K.M.; Gao, S.; Chan, Y.Y.; Shaw, P.C.; Simmonds, M.; Lau, C.B.S. A preclinical exploration of the beneficial effect of tricin-containing medicinal grain in colon health. World J. Tradit. Chin. Med. 2019, 5, 277. [Google Scholar]
- Han, J.M.; Kwon, H.J.; Jung, H.J. Tricin, 4′,5,7-trihydroxy-3′,5′-dimethoxyllavone, exhibits potent antiangiogenic activity in vitro. Int. J. Oncol. 2016, 49, 1497–1504. [Google Scholar] [CrossRef]
- Kim, S.; Go, G.W.; Imm, J.Y. Promotion of glucose uptake in C2C12 myotubes by cereal flavone tricin and its underlying molecular mechanism. J. Agr. Food Chem. 2017, 65, 3819–3826. [Google Scholar] [CrossRef]
- Zhang, H.; Li, H.Q. Tricin enhances osteoblastogenesis through the regulation of Wnt/beta-catenin signaling in human mesenchymal stem cells. Mech. Develop. 2018, 152, 38–43. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, S.S.; Bang, M.H.; Jo, S.K.; Jhee, K.H.; Yang, S.A. Protection against UVB-induced damages in human dermal fibroblasts: Efficacy of tricin isolated from enzyme-treated Zizania latifolia extract. Biosci. Biotech. Bioch. 2019, 83, 551–560. [Google Scholar] [CrossRef]
- Testa, J.R.; Tsichlis, P.N. AKT signaling in normal and malignant cells. Oncogene 2005, 24, 7391–7393. [Google Scholar] [CrossRef]
- Burotto, M.; Chiou, V.L.; Lee, J.M.; Kohn, E.C. The MAPK pathway across different malignancies: A new perspective. Cancer 2014, 120, 3446–3456. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Horgan, P.G.; McMillan, D.C.; Edwards, J. NF-kappaB pathways in the development and progression of colorectal cancer. Transl. Res. 2018, 197, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Imm, J.Y. Antiobesity effect of tricin, a methylated cereal flavone, in high-fat-diet-induced obese mice. J. Agric. Food Chem. 2018, 66, 9989–9994. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.M.; Park, S.H.; Jhee, K.H.; Yang, S.A. Protection against UVB-induced wrinkle formation in SKH-1 hairless mice: Efficacy of tricin isolated from enzyme-treated Zizania latifolia extract. Molecules 2018, 23, 2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malvicini, M.; Gutierrez-Moraga, A.; Rodriguez, M.M.; Gomez-Bustillo, S.; Salazar, L.; Sunkel, C.; Nozal, L.; Salgado, A.; Hidalgo, M.; Lopez-Casas, P.P.; et al. A tricin derivative from Deschampsia antarctica Desv. inhibits colorectal carcinoma growth and liver metastasis through the induction of a specific immune response. Mol. Cancer Ther. 2018, 17, 966–976. [Google Scholar] [CrossRef] [Green Version]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, J.; Ma, J.; Xu, H.; Wang, S. G-MDSC promote the stemness of colorectal cancer cells. Eur. J. Immunol. 2019, 49, 655. [Google Scholar]
- Paluskievicz, C.M.; Cao, X.F.; Abdi, R.; Zheng, P.; Liu, Y.; Bromberg, J.S. T regulatory cells and priming the suppressive tumor microenvironment. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Zhang, X.H.; Kelaria, S.; Kerstetter, J.; Wang, J. The functional and prognostic implications of regulatory T cells in colorectal carcinoma. J. Gastrointest. Oncol. 2015, 6, 307–313. [Google Scholar]
- Xia, Y.; Ohno, T.; Nishii, N.; Bhingare, A.; Tachinami, H.; Kashima, Y.; Nagai, S.; Saito, H.; Nakae, S.; Azuma, M. Endogenous IL-33 exerts CD8(+) T cell antitumor responses overcoming pro-tumor effects by regulatory T cells in a colon carcinoma model. Biochem. Biophys. Res. Commun. 2019, 518, 331–336. [Google Scholar] [CrossRef]
- Kang, B.M.; An, B.K.; Jung, W.S.; Jung, H.K.; Cho, J.H.; Cho, H.W.; Jang, S.J.; Yun, Y.B.; Kuk, Y.I. Anti-inflammatory effect of tricin isolated from Alopecurus aequalis Sobol. on the LPS-induced inflammatory response in RAW 264.7 cells. Int. J. Mol. Med. 2016, 38, 1614–1620. [Google Scholar] [CrossRef] [Green Version]
- Verschoyle, R.E.; Greaves, P.; Cai, H.; Arndt, B.; Broggini, M.; D’Incalci, M.; Riccio, E.; Doppalapudi, R.; Kapetanovic, I.M.; Steward, W.P.; et al. Preliminary safety evaluation of the putative cancer chemopreventive agent tricin, a naturally occurring flavone. Cancer Chemoth. Pharm. 2006, 57, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Boocock, D.J.; Steward, W.P.; Gescher, A.J. Tissue distribution in mice and metabolism in murine and human liver of apigenin and tricin, flavones with putative cancer chemopreventive properties. Cancer Chemoth. Pharm. 2007, 60, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, G.G.L.; Kwok, H.F.; Lee, J.K.M.; Jiang, L.; Chan, K.M.; Cheng, L.; Wong, E.C.W.; Leung, P.C.; Fung, K.P.; Lau, C.B.S. Novel anti-angiogenic effects of aromatic-turmerone, essential oil isolated from spice turmeric. J. Funct. Foods 2015, 15, 243–253. [Google Scholar] [CrossRef]
- Li, M.Y.; Song, L.H.; Yue, G.G.L.; Lee, J.K.M.; Zhao, L.M.; Li, L.; Zhou, X.N.; Tsui, S.K.W.; Ng, S.S.M.; Fung, K.P.; et al. Bigelovin triggered apoptosis in colorectal cancer in vitro and in vivo via upregulating death receptor 5 and reactive oxidative species. Sci. Rep. 2017, 7, 42176. [Google Scholar] [CrossRef] [Green Version]
- Leung, H.W.; Wang, Z.; Yue, G.G.L.; Zhao, S.M.; Lee, J.K.; Fung, K.P.; Leung, P.C.; Lau, C.B.S.; Tan, N.H. Cyclopeptide RA-V inhibits cell adhesion and invasion in both estrogen receptor positive and negative breast cancer cells via PI3K/AKT and NF-kappaB signaling pathways. Biochim. Biophys. Acta 2015, 1853, 1827–1840. [Google Scholar] [CrossRef] [Green Version]
- Ko, C.H.; Yue, G.G.L.; Gao, S.; Luo, K.W.; Siu, W.S.; Shum, W.T.; Shiu, H.T.; Lee, J.K.M.; Li, G.; Leung, P.C.; et al. Evaluation of the combined use of metronomic zoledronic acid and Coriolus versicolor in intratibial breast cancer mouse model. J. Ethnopharmacol. 2017, 204, 77–85. [Google Scholar] [CrossRef]
- Li, M.; Yue, G.G.L.; Song, L.H.; Huang, M.B.; Lee, J.K.M.; Tsui, S.K.W.; Fung, K.P.; Tan, N.H.; Lau, C.B.S. Natural small molecule bigelovin suppresses orthotopic colorectal tumor growth and inhibits colorectal cancer metastasis via IL6/STAT3 pathway. Biochem. Pharm. 2018, 150, 189–199. [Google Scholar] [CrossRef]
- Yue, G.G.L.; Wong, L.S.; Leung, H.W.; Gao, S.; Tsang, J.Y.S.; Lin, Z.X.; Tse, G.M.K.; Lau, C.B.S. Evaluation of the safety profiles of estrogenic Chinese herbal medicines in breast cancer. Phytomedicine 2019, 56, 103–117. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compound tricin, and the grains Hordei Fructus Germinatu, Oryzae Fructus Germinatus and Setariae Fructus Germinatus are available from the authors for reference. |






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, G.G.-L.; Gao, S.; Lee, J.K.-M.; Chan, Y.-Y.; Wong, E.C.-W.; Zheng, T.; Li, X.-X.; Shaw, P.-C.; Simmonds, M.S.J.; Lau, C.B.-S. A Natural Flavone Tricin from Grains Can Alleviate Tumor Growth and Lung Metastasis in Colorectal Tumor Mice. Molecules 2020, 25, 3730. https://doi.org/10.3390/molecules25163730
Yue GG-L, Gao S, Lee JK-M, Chan Y-Y, Wong EC-W, Zheng T, Li X-X, Shaw P-C, Simmonds MSJ, Lau CB-S. A Natural Flavone Tricin from Grains Can Alleviate Tumor Growth and Lung Metastasis in Colorectal Tumor Mice. Molecules. 2020; 25(16):3730. https://doi.org/10.3390/molecules25163730
Chicago/Turabian StyleYue, Grace Gar-Lee, Si Gao, Julia Kin-Ming Lee, Yuk-Yu Chan, Eric Chun-Wai Wong, Tao Zheng, Xiao-Xiao Li, Pang-Chui Shaw, Monique S. J. Simmonds, and Clara Bik-San Lau. 2020. "A Natural Flavone Tricin from Grains Can Alleviate Tumor Growth and Lung Metastasis in Colorectal Tumor Mice" Molecules 25, no. 16: 3730. https://doi.org/10.3390/molecules25163730