
Model-Based Optimization of a Fed-Batch Bioreactor for mAb Production Using a Hybridoma Cell Culture


Abstract
:1. Introduction
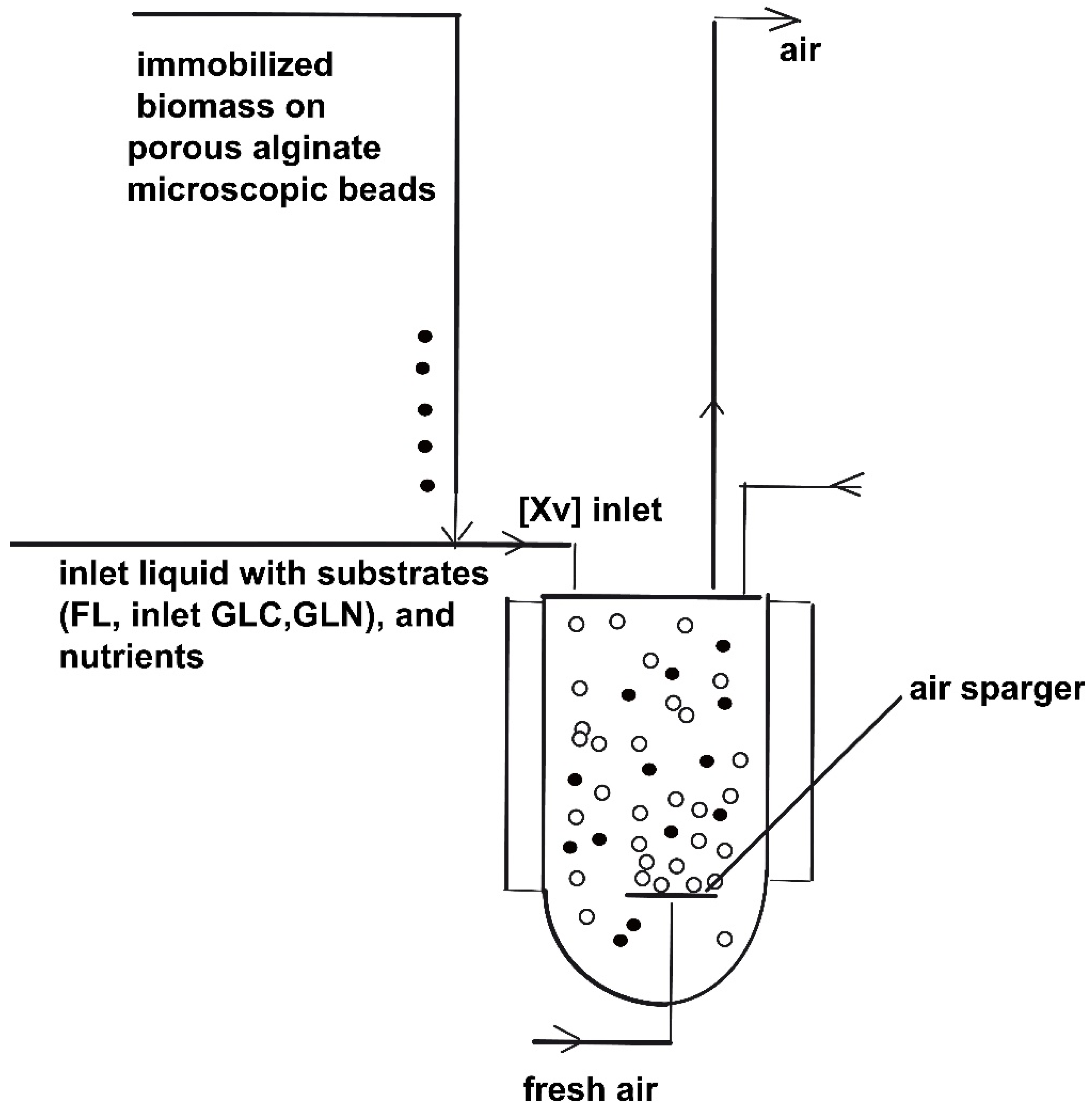
2. SBR Culture and Bioprocess Dynamics
3. Formulation of the Bioreactor Optimization Problem
3.1. Control Variables Selection
- (i).
- the continuously added liquid flow rate FL,j (j = 1, …, Ndiv);
- (ii).
- the time stepwise added [GLC]inlet,j; [GLN]inlet,j; [Xv]inlet,j (j = 1, …, Ndiv);
- (iii).
- the FBR initial condition, that is, the initial liquid flow rate FL,0, and the initial substrates (as shown in Table 2), that is:[GLC]0 = [GLC](t = 0) = [GLC]inlet,1;[GLN]0 = [GLN](t = 0) = [GLN]inlet,1; [Xv]0 = [Xv](t = 0) = [Xv]inlet,1.
3.2. Objective Function (Ω) Choice
3.3. Problem Constraints
- (a)
- The FBR model (1)–(4) including the bioprocess kinetic model (Table 2);
- (b)
- The FBR initial condition, that is: [GLC]0, [GLN]0; FL,0; [Xv]0 = [Xt]0 (adopted);
- (c)
- The initial [mAb]0, [AMM]0, [LAC]0, adopted at values given in Table 3;
- (d)
- To limit the excessive consumption of raw materials, feasible searching ranges are imposed to the control/decision variable (with limits specified in Table 4), that is:
[Xv]inlet,min ≤ [Xv]inlet,j ≤ [Xv]inlet,max
3.4. Searching (Control) Variables and Problem Formulation
3.5. The Problem Solution
3.6. (Ndiv) and Operating Alternative Choice
- (a)
- by choosing unequal time-arcs, of lengths to be determined by the optimization rule;
- (b)
- by considering the whole batch time as an optimization variable;
- (c)
- by increasing the number of equal time-arcs (Ndiv) to obtain a more “refined” and versatile FBR operating policy;
- (d)
- by considering the search min/max limits of the control variables as unknown (to be determined);
- (e)
- by feeding the bioreactor with solutions of uniform concentrations over a small/large number (Ndiv) of time-arcs.
3.7. The Used Solvers
4. In-Silico Optimization Results
- (1)
- FBR-SP1. For the adopted Ndiv = 5, with equal time-arcs, and by using narrow search intervals for the control variables [GLC]inlet,j; [GLN]inlet,j; [Xv]inlet,j; and FL,j specified in Table 4 (in the SP1 row), the obtained optimal operating policy SP1 (for every time-arc) is presented in Table 4, together with the key-species dynamics in Figure 3. Final liquid volume is 1.27 × VL,0
- (2)
- FBR-SP2. For Ndiv = 5 and equal time-arcs, but using wider search intervals for the (above mentioned) control variables, as specified in Table 4 (in the SP2 row), the obtained optimal operating policy SP2 of the control variables is presented for every time-arc in Table 4, together with the key-species dynamics in Figure 4. The final liquid volume is 1.1 × VL,0.
- (3)
- FBR-SP3. For an adopted smaller Ndiv = 2 with equal time-arcs, but using the same wide search intervals for the control variables as for the SP2 case (SP3 row in Table 4), the obtained optimal operating policy SP3 of the control variables is presented for every time-arc in Table 4, together with the key-species dynamics during the batch in Figure 5. The final liquid volume is 1.1 × VL,0.
5. Results and Discussion
- (I).
- In all the simulated alternatives, the FBR performance (in terms of produced mg mAb/L) is better than that realized by the BR (see the results summarized in Table 5), even if the overall batch time is the same (100 h), and a simple operating policy with equal time-arcs, in a small number (2–5) is used. The FBR productivity is up to 6× higher than that of the BR, while FBR is using fewer raw materials (Table 5). In Table 5, the mAb productivity is expressed in the absolute terms of Max [mAb] [mg/L]. Other indices, such as Max [mAb][mg/cells.h] can be used as well, by combining the data of Table 4 and Table 5. Being an intensive index, according to “Max [mAb][mg/cells.h]”, the BR appears more favorable because it uses less total [Xv]. However, due to the large value of the product vs. the used biomass, such a poor advantage of the BR becomes negligible.
- (II).
- The study points out the major influence of the control variable setting ranges (narrower or wider) used by the optimization rule, on the obtained efficiency of the FBR optimal policy. More specifically, according to the results of Table 5, it turns out that:
- (IIa).
- The GLC consumption during FBR is ca. 1/2 for SP1, or 1/3 for SP3 than that of the BR case. Similarly, fewer GLN was also consumed. The biomass (Xv) consumption is roughly the same because of a smaller Ndiv (SP3), or of narrower search ranges of control variables (SP1).
- (IIb).
- (IIc).
- The price paid by the FBR-SP2 to achieve the best performances compared to BR-SPBR, and FBR-SP1, or FBR-SP3 is a higher consumption of raw materials, i.e., (vs. BR) of ca. 1.5× more GLC and GLN, and 10× more biomass.
- (IId).
- The FBR-SP3 (Xv,0, and mAb net productivity) policy appears to be somehow intermediate between the FBR-SP2 and BR-SPBR. Compared to the SPBR, the raw material consumptions are smaller, but the realized FBR performances are better, because an operation with Ndiv = 2 is more versatile than that of the BR with the initial load being the only optimization option.
- (IIe).
- The used biomass is generally higher in the FBR cases compared to those of BR (Table 5). Thus, the used (Xv) is roughly the same for SP3 (Ndiv = 2), but 2× for FBR-SP1, or 10× for FBR-SP2.
- (III).
- The GLC consumption for FBR operating case depends on the inlet GLC policy (the [GLC]inlet,j term in Table 2), and on the used control variable dynamics (Xv, GLN).
- (IV).
- A comparison indicates that the GLC dynamics of the SP1 vs. SP2 of the FBR (with Ndiv = 5) is depending on not only the searching interval chosen for the control variable [GLC]inlet,j, but also on the other species dynamics. Thus, if one compares the (Xv) plots of SP1 in Figure 3 to that of FBR-SP2 in Figure 4, it is easy to observe and explain that when (Xv) is high, GLC consumption is also high, in spite of a larger inlet [GLC]. This clearly shows that the FBR optimization must consider all variables simultaneously.
- (V).
- In the BR-SPBR case of LG17, a less flexible feeding explains its modest mAb productivity. The BR species dynamics (in Figure 2) is comparable to those of FBR-SP3 (Ndiv = 2; in Figure 5), that is, GLC is quickly consumed during the first half of the batch, and the biomass displays a pronounced peak in the first half of the batch.In the end, it is worth mentioning that the present numerical engineering analysis presents multiple elements of novelty as briefly mentioned in Section 6 “Conclusions”.
- (VI).
- Our results prove the multiple advantages obtained when using FBR operated with multiple control variables following very versatile optimal feeding policies consisting in time stepwise variable of: (i) the feeding liquid flow rate, (ii) the added [GLC], (iii) the added [GLN], and (iv) the added [Xv] over the batch.The in-silico analysis of the paper proved how such an optimal FBR operation is leading to quick results, easy to interpret and to implement, being more flexible and effective due to a larger number of degree of freedom (coming from the multiple control variables, and from their variable time stepwise policy), in spite of an economically advantageous small number of employed time-arcs (Ndiv) compared to some optimal policies of a similar FBR reported in the literature. For instance, [18] uses only an exponential trajectory of the feeding liquid flow rate, and only the inlet levels of [GLC] and [GLN] as control variables, all being obtained by using a hybrid deterministic (differential, intrinsic)–empiric (macroscopic) model.
- (VII).
- The present in-silico (model-based) analysis have not been experimentally validated. However, as long as various forms of the used LGM were experimentally validated in a multiple and independent manner by LG17, and by [33,54] (Section 2), the results obtained by our numerical analysis shows sufficient credibility from the engineering point of view, from the following reasons:
- (a)
- Even if an experimental validation of the derived optimal policy FBR-SP2 policy is missing, our paper presents a very strong engineering value by exemplifying, in a relatively simple manner, a numerical procedure (process model-based) that can be used to solve similar complex optimization problems of FBR.
- (b)
- Such an FBR optimal control rule is possible because most kinetic models of moderate complexity are very flexible. Thus, if significant inconsistencies are observed between the model-predicted bioreactor dynamics (e.g., optimal policy SP2 in the present case) and the experimental data, then the optimization stage is applied again by using the same rule, but after performing an intermediate numerical-analysis step (between batches) necessary to improve the model adequacy (the so-called “model updating” based on the online measurements).
- (VIII).
- As displayed in Figure 2, Figure 3, Figure 4 and Figure 5 inhibition given by the increasing LAC, and AMM by-product concentration cannot be diminished by simple manipulations of the chosen control variables, even if the derived operating policy is an optimal one. However, the adopted kinetic model is able to fairly predict the dynamics of these inhibitory species. But the adverse side effects, such as a low pH, or a hyper-osmotic stress (due to the nutrient feeds and base additions to control pH) cannot be avoided by the above-used engineering (model-based) rules. As revealed in the literature, “biological” solutions are used instead to cope with such a problem. For instance, to reduce the LAC production, the use of adapted CHO (Chinese Hamster Ovary) cells can be an alternative [43].
- (IX).
- The used time-arcs of constant control variables are of 20 h (for SP1, SP2), and of 50 h (for SP3). Such an operation cannot raise special operating problems for a FBR, with also including PAT (Process analytical technology) tools. This is an additional argument not using a larger number of small time-arcs (Ndiv, see Section 3.6).
- (X).
- The comparison in Table 5 of the obtained optimal policies FBR-SP2 and FBR-SP3 with those from the literature for a BR [44,54], or a FBR [33] operated with the same cell culture, indicates better performances despite a longer batch time, and a larger number of substrate feeding solutions of those in the literature.
6. Conclusions
Funding
Conflicts of Interest
Abbreviations
| Ci | Species i concentration |
| Complex functions in Equation (2) accounting for the biomass growth inhibition or death inhibition respectively | |
| FL | Liquid feed flow rate |
| Model rate constant vector | |
| , , , , , etc. | Rate constants given in Table 2, the index “j” relates to the all forms of these constants. |
| Ndiv | Number of equal divisions (“time-arcs”) of the batch time tf |
| ri | Species ”i” reaction rate |
| t | Time |
| tf | Batch time |
| VL | Liquid volume in the bioreactor |
| XV | Viable cell density |
| Xt | Total cell density |
| Index | |
| f | Final |
| inlet | Value in the bioreactor inlet |
| max | Maximum |
| min | Minimum |
| 0,o | Initial |
| Greeks | |
| Discrete interval | |
| , γ, λ, μmax, μd,max | Rate constants given in Table 2 |
| μ | Specific growth rate of viable biomass in Table 2 |
| μd | Specific cell death rate in Table 2 |
| Ω | The optimization objective function in eqn.(1) |
| AMM | Ammonia |
| BR | Batch reactor |
| BRP | Batch reactor with intermittent addition of biomass/raw materials |
| CCM | Central carbon metabolism |
| DO | Dissolved oxygen |
| EKM | Extended kinetic model of [33,54] |
| FBR | Fed-batch bioreactor |
| GLC | Glucose |
| GLN | Glutamine |
| LAC | Lactate |
| LDH | Lactate dehydrogenase |
| LGM | The kinetic model of [44] |
| LG17 | Notation for the reference [44] |
| MA(S)CR | Mechanically agitated (semi-)continuous reactor |
| mAb | Monoclonal antibody |
| Max | Maximum |
| MINLP | Mixed-integer NLP |
| MMA | The multi-modal optimization solver of [48,61] |
| NADH | Nicotinamide adenine dinucleotide (reduced form) |
| NLP | Nonlinear programming (numerical rules for solving optimization problems) |
| PAT | Process Analytical Technology tools. |
| SP | Bioreactor setpoint |
| SPBR | BR nominal setpoint of [44] (Table 3) |
| SBR | Semi-batch reactor |
| SeqBR | Sequential batch reactor |
| TPFB | Three-phase fluidized-bed bioreactor |
| [x] | x species concentration |
References
- Moser, A. Bioprocess Technology—Kinetics and Reactors; Springer: Berlin, Germany, 1988. [Google Scholar]
- Heinemann, M.; Panke, S. Synthetic Biology—Putting engineering into biology. Bioinformatics 2006, 22, 2790–2799. [Google Scholar] [CrossRef] [Green Version]
- Maria, G. In-Silico Design of Genetic Modified Micro-Organisms (GMO) of Industrial Use, by Using Systems Biology and (Bio)chemical Engineering Tools, Simi Valley; Juniper Publ.: Irvine, CA, USA, 2018; ISBN 978-1-946628-12-1. Available online: https://juniperpublishers.com/ebook-info.php (accessed on 28 November 2020).
- Lübbert, A.; Jørgensen, S.B. Bioreactor performance: A more scientific approach for practice. J. Biotechnol. 2001, 85, 187–212. [Google Scholar] [CrossRef]
- Engasser, J.-M. Bioreactor engineering: The design and optimization of reactors with living cells. Chem. Eng. Sci. 1988, 43, 1739–1748. [Google Scholar] [CrossRef]
- Gavrilescu, M.; Chisti, Y. Biotechnology—A sustainable alternative for chemical industry. Biotechnol. Adv. 2005, 23, 471–499. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, J. Bioreactors for Tissue Engineering Principles, Design and Operation; Al-Rubeai, M., Ed.; Springer: Berlin, Germany, 2005. [Google Scholar]
- Nedovic, V.; Willaert, R. Applications of Cell Immobilisation Technology; Springer: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Abel, O.; Marquardt, W. Scenario-integrated online optimisation of batch reactors. J. Process Control 2003, 13, 703–715. [Google Scholar] [CrossRef]
- Bonvin, D. Optimal operation of batch reactors—A personal view. J. Process Control 1998, 8, 355–368. [Google Scholar] [CrossRef]
- Martinez, E. Batch-to-batch optimization of batch processes using the STATSIMPLEX search method. In Proceedings of the 2nd Mercosur Congress on Chemical Engineering, Rio de Janeiro, Costa Verde, Brasil, 31 March 2005. [Google Scholar]
- Dewasme, L.; Côte, F.; Filee, P.; Hantson, A.-L.; Wouwer, A.V. Macroscopic Dynamic Modeling of Sequential Batch Cultures of Hybridoma Cells: An Experimental Validation. Bioengineering 2017, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- DiBiasio, D. Introduction to the control of biological reactors. In Chemical Engineering Problems in Biotechnology; Shuler, M.L., Ed.; American Institute of Chemical Engineers: New York, NY, USA, 1989; pp. 351–391. ISBN 9780816904693. [Google Scholar]
- Maria, G.; Crişan, M. Operation of a mechanically agitated semi-continuous multi-enzymatic reactor by using the Pareto-optimal multiple front method. J. Process Control 2017, 53, 95–105. [Google Scholar] [CrossRef]
- Smets, I.Y.; Claes, J.E.; November, E.J.; Bastin, G.P.; Van Impe, J.F. Optimal adaptive control of (bio)chemical reactors: Past, present and future. J. Process Control 2004, 14, 795–805. [Google Scholar] [CrossRef]
- Şcoban, A.G.; Maria, G. Model-based optimization of the feeding policy of a fluidized bed bioreactor for mercury uptake by immobilizedPseudomonas putidacells. Asia Pac. J. Chem. Eng. 2016, 11, 721–734. [Google Scholar] [CrossRef]
- Dewasme, L.L.; Amribt, Z.; Santos, L.O.; Hantson, A.-L.; Bogaerts, P.; Wouwer, A.V. Hybridoma cell culture optimization using nonlinear model predictive control. In Proceedings of the 12th IFAC Symposium on Computer Applications in Biotechnology, Mumbai, India, 16–18 December 2013; International Federation of Automatic Control: Laxenburg, Austria, 2013; Volume 46, pp. 60–65. [Google Scholar]
- Amribt, Z.; Dewasme, L.; Wouwer, A.V.; Bogaerts, P. Optimization and robustness analysis of hybridoma cell fed-batch cultures using the overflow metabolism model. Bioprocess Biosyst. Eng. 2014, 37, 1637–1652. [Google Scholar] [CrossRef] [PubMed]
- Binette, J.-C.; Srinivasan, B. On the Use of Nonlinear Model Predictive Control without Parameter Adaptation for Batch Processes. Processes 2016, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Franco-Lara, E.; Weuster-Botz, D. Estimation of optimal feeding strategies for fed-batch bioprocesses. Bioprocess Biosyst. Eng. 2005, 28, 71. [Google Scholar] [CrossRef] [Green Version]
- Mendes, R.; Rocha, I.; Pinto, J.P.; Ferreira, E.C.; Rocha, M. Differential Evolution for the Offline and Online Optimization of Fed-Batch Fermentation Processes. In Advances in Differential Evolution. Studies in Computational Intelligence, Chakraborty; Chakraborty, U.K., Ed.; Springer: Berlin, Germany, 2008; pp. 299–317. [Google Scholar]
- Crisan, M.; Maria, G. Modular Simulation to Determine the Optimal Operating Policy of a Batch Reactor for the Enzymatic Fructose Reduction to Mannitol with the in situ Continuous Enzymatic Regeneration of the NAD Cofactor. Rev. Chim. 2017, 68, 2196–2203. [Google Scholar] [CrossRef]
- Maria, G. Enzymatic reactor selection and derivation of the optimal operation policy, by using a model-based modular simulation platform. Comput. Chem. Eng. 2012, 36, 325–341. [Google Scholar] [CrossRef]
- Maria, G.; Crisan, M. Evaluation of optimal operation alternatives of reactors used ford-glucose oxidation in a bi-enzymatic system with a complex deactivation kinetics. Asia Pac. J. Chem. Eng. 2014, 10, 22–44. [Google Scholar] [CrossRef]
- Maria, G. Model-based heuristic optimized operating policies for d-glucose oxidation in a batch reactor with pulsate addition of enzyme. Comput. Chem. Eng. 2007, 31, 1231–1241. [Google Scholar] [CrossRef]
- Lee, J.; Lee, K.S.; Lee, J.H.; Park, S. An online batch span minimization and quality control strategy for batch and semi-batch processes. Control Eng. Pract. 2001, 9, 901–909. [Google Scholar] [CrossRef]
- Ruppen, D.; Bonvin, D.; Rippin, D. Implementation of adaptive optimal operation for a semi-batch reaction system. Comput. Chem. Eng. 1998, 22, 185–199. [Google Scholar] [CrossRef]
- Srinivasan, B.; Bonvin, D.; Visser, E.; Palanki, S. Dynamic optimization of batch processes. Comput. Chem. Eng. 2003, 27, 27–44. [Google Scholar] [CrossRef]
- Rao, M.; Qiu, H. Process Control Engineering: A Textbook for Chemical, Mechanical and Electrical Engineers; Gordon and Breach Science Publ.: Amsterdam, The Netherlands, 1993. [Google Scholar]
- Hartig, F.; Keil, F.J.; Luus, R. Comparison of Optimization Methods for a Fed-Batch Reactor. Hung. J. Ind. Chem. 1995, 23, 81–160. [Google Scholar]
- Koutinas, M.; Kiparissides, A.; Pistikopoulos, E.N.; Mantalaris, A. Bioprocess systems engineering: Transferring traditional process engineering principles to industrial biotechnology. Comput. Struct. Biotechnol. J. 2012, 3, e201210022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsangaris, D.; Baltzis, B. Evaluation of batch and semi-batch reactor operation for enzymatic reactions with inhibitory kinetics. Chem. Eng. Sci. 1996, 51, 2757–2762. [Google Scholar] [CrossRef]
- Kontoravdi, C.; Pistikopoulos, E.N.; Mantalaris, A. Systematic development of predictive mathematical models for animal cell cultures. Comput. Chem. Eng. 2010, 34, 1192–1198. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, S.S.; Palsson, B.Ø. Effect of initial cell density on hybridoma growth, metabolism, and monoclonal antibody production. J. Biotechnol. 1990, 16, 259–278. [Google Scholar] [CrossRef] [Green Version]
- Koller, M. A Review on Established and Emerging Fermentation Schemes for Microbial Production of Polyhydroxyalkanoate (PHA) Biopolyesters. Fermentation 2018, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Sawatzki, A.; Hans, S.; Narayanan, H.; Haby, B.; Krausch, N.; Sokolov, M.; Glauche, F.; Riedel, S.L.; Neubauer, P.; Bournazou, M.N.C. Accelerated Bioprocess Development of Endopolygalacturonase-Production with Saccharomyces cerevisiae Using Multivariate Prediction in a 48 Mini-Bioreactor Automated Platform. Bioengineering 2018, 5, 101. [Google Scholar] [CrossRef] [Green Version]
- Maria, G.; Dan, A. Derivation of optimal operating policies under safety and technological constraints for the acetoacetylation of pyrrole in a semi-batch catalytic reactor. Comput. Chem. Eng. 2011, 35, 177–189. [Google Scholar] [CrossRef]
- Avili, M.G.; Fazaelipoor, M.H.; Jafari, S.A.; Ataei, S.A. Comparison between Batch and Fed-Batch Production of Rhamnolipid by Pseudomonas aeruginosa. Iran. J. Biotechnol. 2012, 10, 263–269. Available online: https://www.researchgate.net/publication/288966207 (accessed on 3 June 2004).
- Lee, G.M.; Varma, A.H.; Palsson, B.O. Production of monoclonal antibody using free-suspended and immobilized hybridoma cells: Effect of serum. Biotechnol. Bioeng. 1991, 38, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Hauser, H.; Wagner, R. Mammalian Cell Biotechnology in Protein Production; Walter de Gruyter Publ.: New York, NY, USA, 1997. [Google Scholar]
- Dorka, P. Modelling Batch and Fed-Batch Mammalian Cell Cultures for Optimizing MAb Productivity. Master’s Thesis, University of Waterloo, Waterloo, ON, Canada, 2007. [Google Scholar]
- Li, F.; Vijayasankaran, N.; Shen, A.; Kiss, R.; Amanullah, A. Cell culture processes for monoclonal antibody production. mAbs 2010, 2, 466–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freund, N.W.; Croughan, M.S. A Simple Method to Reduce both Lactic Acid and Ammonium Production in Industrial Animal Cell Culture. Int. J. Mol. Sci. 2018, 19, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Gunawan, R. Bioprocess optimization under uncertainty using ensemble modeling. J. Biotechnol. 2017, 244, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Kadam, J.; Schlegel, M.; Srinivasan, B.; Bonvin, D.; Marquardt, W. Dynamic optimization in the presence of uncertainty: From offline nominal solution to measurement-based implementation. J. Process Control 2007, 17, 389–398. [Google Scholar] [CrossRef]
- Terwiesch, P.; Agarwal, M.; Rippin, D.W. Batch unit optimization with imperfect modelling: A survey. J. Process Control 1994, 4, 238–258. [Google Scholar] [CrossRef]
- Watanabe, N.; Nishimura, Y.; Matsubara, M. Optimal design of chemical processes involving parameter uncertainty. Chem. Eng. Sci. 1973, 28, 905–913. [Google Scholar] [CrossRef]
- Maria, G. A review of algorithms and trends in kinetic model identification for Chemical and Biochemical Systems. Chem. Biochem. Eng. Q. 2004, 18, 195–222. [Google Scholar]
- Maria, G. Adaptive random search and shortcut techniques for process model identification and monitoring. In Foundations of Computer Aided Process Operations; Pekny, J.F., Blau, G.E., Carnahan, B., Eds.; AIChE symposium series; American Institute of Chemical Engineers: New York, NY, USA, 1998; Volume 94, pp. 351–359. [Google Scholar]
- Selimoglu, S.M.; Ayyildiz-Tamis, D.; Gurhan, I.D.; Elibol, M. Purification of alginate and feasible production of monoclonal antibodies by the alginate-immobilized hybridoma cells. J. Biosci. Bioeng. 2012, 113, 233–238. [Google Scholar] [CrossRef]
- Selimoglu, S.M.; Elibol, M. Alginate as an immobilization material for MAb production via encapsulated hybridoma cells. Crit. Rev. Biotechnol. 2010, 30, 145–159. [Google Scholar] [CrossRef]
- Lee, G.M.; Kim, S.J.; Palsson, B.O. Enhanced specific antibody productivity of calcium alginate-entrapped hybridoma is cell line-specific. Cytotechnology 1994, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Quiroga-Campano, A.L.; Panoskaltsis, N.; Mantalaris, A. Energy-based culture medium design for biomanufacturing optimization: A case study in monoclonal antibody production by GS-NS0 cells. Metab. Eng. 2018, 47, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Kiparissides, A.; Koutinas, M.; Kontoravdi, C.; Mantalaris, A.; Pistikopoulos, E.N. ‘Closing the loop’ in biological systems modeling—From the in silico to the in vitro. Automatica 2011, 47, 1147–1155. [Google Scholar] [CrossRef]
- Venables, D. Two Stage Chemostat Studies of Hybridoma Growth, Nutrient Utilization, and Monoclonal Antibody Production. Ph.D. Thesis, University of Surrey (MI), Stag Hill, Guildford, UK, 1994. [Google Scholar]
- Bree, M.A.; Dhurjati, P.; Geoghegan, R.F.; Robnett, B. Kinetic modelling of hybridoma cell growth and immunoglobulin production in a large-scale suspension culture. Biotechnol. Bioeng. 1988, 32, 1067–1072. [Google Scholar] [CrossRef]
- Zeng, A.P.; Deckwer, W.D.; Hu, W.S. Determinants and rate laws of growth and death of hybridoma cells in continuous culture. Biotechnol. Bioeng. 1998, 57, 642–654. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Yap, P.-K.; Teoh, A.-P. Correlation between steady-state cell concentration and cell death of hybridoma cultures in chemostat. Biotechnol. Bioeng. 1995, 45, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Bharat, A. Process Analytical Technology (PAT), Msc-Diss., P.D.V.V.P.F’S. College of Pharmacy, AhmedNagar (India). 2013. Available online: https://www.slideshare.net/anjalibharat19/process-analytical-tchnology (accessed on 28 November 2020).
- Loeblein, C.; Perkins, J.; Srinivasan, B.; Bonvin, D. Performance analysis of online batch optimization systems. Comput. Chem. Eng. 1997, 21, S867–S872. [Google Scholar] [CrossRef] [Green Version]
- Maria, G. ARS Combination with an Evolutionary Algorithm for Solving MINLP Optimization Problems. In Modelling, Identification and Control; Hamza, M.H., Ed.; IASTED/ACTA Press: Anaheim, CA, USA, 2003; pp. 112–118. Available online: http://www.sigmod.org/sigmod/dblp/db/conf/mic/mic2003.html (accessed on 3 June 2004).
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reactor Type | Notation [Examples] | Operation; Modeling Hypotheses |
|---|---|---|
 | (i) simple Batch Reactor (BR) Examples: [9,10] | isothermal, iso-pH, and iso-DO (air sparger); perfectly mixed liquid phase (with no concentration gradients, by using mechanical agitation). Reactants/biomass added at the beginning of the batch only. |
 | (ii) SeqBR (Sequential Batch-to-batch Reactor) Examples: [11,12] | Ibidem. Reactants and/or biomass added at the beginning of each batch, in optimized amounts (to be determined) |
 | (iii) Semi-Batch (fed-batch) Reactor (SBR or FBR). Examples: [13,14,15,16,17,18,19,20,21] | Ibidem. Substrates/biomass/supplements added during the batch by following a certain (optimal) policy (to be determined) |
 | (iv) BRP (Batch Reactor with intermittent/Pulse-like additions of biocatalyst/substrates). Examples: [22,23,24,25] | Ibidem. Reactants and/or biomass added during the batch in a Pulse-like additions of equal/uneven solution volumes, with a certain frequency (to be determined) |
 | (v) continuously operated packed-bed columns, FXBR (FiXed-Bed continuous bioReactor) Examples: [23] | immobilized enzyme on a porous support packed in columns; continuous fed of the substrate/nutrient solution; continuous solution output; various aeration alternatives. Model hypotheses: isothermal, ideal plug-flow reactor of constant volume, with model dynamic terms allowing simulating transient operating conditions and the continuous enzyme/biomass deactivation |
 | (vi) MA(S)CR (Mechanically Agitated (Semi-)Continuous Reactor) (three-phases). Examples: [14] | immobilized enzyme on porous support suspended in the mechanically agitated bioreactor, with sparged gas (air); continuous fed of the substrate/nutrient solution, with/without continuous evacuation; Model hypotheses: isothermal, ideal perfectly mixed liquid phase (with no concentration gradients, by using mechanical agitation, aeration), with model dynamic terms allowing simulating transient operating conditions and the continuous enzyme/biomass deactivation. Substrates/biomass can be added with a constant/variable feed flow rate (to be determined). |
| Species | Parameters | Remarks |
|---|---|---|
| Biomass balance: Viable biomass balance: = control variable; j = 1, …, time stepwise values to be optimized; = ; to be optimized; Total biomass balance: (adopted); = where: | = 0.0551 h−1 = 0.058 h−1 = 0.75 mM = 0.075 mM = 172 mM = 28.5 mM = 0.03 h−1 = 1.76 mM | LGM |
| Balance of other species: , = control variable; j = 1, …, time stepwise values to be optimized; = ; to be optimized where: | = 1.06 × 108 cell/mmol = 4.85 × 10−14 mmol/cell.h | LGM |
| , = control variable; j = 1, …, time stepwise values to be optimized; = ; to be optimized, where: = 0.0096 h−1; = 5.57 × 108 cell/mmol = = −0.00067 mmol/cell.h | LGM | |
| Liquid volume dynamics: , (a) For the adopted = 5, the j = 1, …, time-arcs switching points are: T1 = 20 h.; T2 = 40 h.; T3 = 60 h.; T4 = 80 h.; = 100 h., where time stepwise values are to be determined together with the other control variables to ensure an optimal FBR operation; (b) For the adopted = 2, the j = 1, …, time-arcs switching points are: T1 = 50 h.; = 100 h., with where time stepwise values are to be determined together with the other control variables to ensure an optimal FBR operation; | This paper | |
| , = 0 where: | = 1.4 L | LGM |
= 0; = 0.31 mM where: = 0.0096 h−1; = 0.427 L | LGM | |
= 0; = 80.6 mg/L | γ = 0.1 h λ = 7.21 × 10−9 mg/(cell·h) | LGM |
| Parameter | Nominal Value | Remarks (*) |
|---|---|---|
| Total cell initial density (Xt,0) | 2 × 108 Cell/L | Ref. to reactor-lq. |
| Viable cell initial density (XV,0) | 2 × 108 Cell/L | Ref. to reactor-lq. |
| Glucose initial concentration, [GLC]0 | 29.1, mM | |
| Glutamine initial concentration, [GLN]0 | 4.9, mM | |
| Lactate initial concentration, [LAC]0 | 0, mM | |
| Ammonia initial concentration, [AMM]0 | 0.31, mM | |
| Monoclonal antibody initial concentration, [mAb]0 | 80.6, mg/L | Ref. to reactor-lq. |
| Temperature | 35–37 °C | [42] |
| pH (buffer, using CO2 injection) | 7 | See an optimal policy given by [42] |
| Aeration in excess, nutrients in sufficient amounts | [42,55] | |
| Initial volume of the liquid in the bioreactor (VL,0) | 1 L | LGM |
| Batch time (tf) | approx. 100 h. | LGM |
| Reactor SP | Searching Policy | Control Variables | Obs. | ||||
|---|---|---|---|---|---|---|---|
| SPBR (BR) Optimal Values | Sensitivity Analysis (Exhaustive) | Initial Values of the BR Content | |||||
| [GLC], mM | [GLN], mM | Xv,0 = Xt,0, Cell/L | Max [mAb](t), (mg/L) | ||||
| 29.1 | 4.9 | 2 × 108 | 1254.6 | LGM | |||
| SP1(a) (FBR) Optimal values (d) | Searching variables | FL, L/h. | [GLC]inlet, mM | [GLN]inlet, mM | Xv,inlet Cell/L | This paper | |
| Searching ranges | (10−4–10−2) | (25–100) | (5–25) | (2 × 108–2 × 109) | |||
| Multi-dimensional optimization | Inlet optimal values of the FBR control variables | ||||||
| FL, (b,c), L/h. | [GLC]inlet mM | [GLN]inlet mM | Xv,inlet Cell/L | Max [mAb](t), (mg/L) | |||
| Time interval (0, 20) h. | 10−3 | 96.62 | 17.75 | 2 × 108 | This paper | ||
| Time interval (20, 40) h. | 9.55 × 10−3 | 45.13 | 9.52 | 1.7 × 109 | |||
| Time interval (40, 60) h. | 10−3 | 26.99 | 16.58 | 1.62 × 109 | |||
| Time interval (60, 80) h. | 10−3 | 87.80 | 21.77 | 1.33 × 109 | |||
| Time interval (80, 100) h | 10−3 | 68.42 | 14.43 | 5.74 × 108 | |||
| Optimal value of Max [mAb](t), (mg/L) | 1351.3 | This paper | |||||
| SP2(a)(FBR) Optimal values (d) | Searching variables | FL, L/h. | [GLC]inlet, mM | [GLN]inlet, mM | Xv,inlet, Cell/L | This paper | |
| Searching ranges | (10−4–5 × 10−2) | (25–150) | (5–25) | (2 × 108–5 × 109) | |||
| Multi-dimensional optimization | Inlet optimal values of the FBR control variables | ||||||
| FL (b,c), L/h. | [GLC]inlet mM | [GLN]inlet mM | Xv,inlet, Cell/L | Max [mAb](t), (mg/L) | |||
| Time interval (0, 20) h. | 10−3 | 141.63 | 17.76 | 4.38 × 109 | This paper | ||
| Time interval (20, 40) h. | 10−3 | 55.81 | 9.52 | 4.20 × 109 | |||
| Time interval (40, 60) h. | 10−3 | 25.60 | 16.58 | 3.98 × 109 | |||
| Time interval (60, 80) h. | 10−3 | 126.92 | 21.77 | 3.21 × 109 | |||
| Time interval (80, 100) h | 10−3 | 94.62 | 14.43 | 1.20 × 109 | |||
| Optimal value of Max [mAb](t), (mg/L) | 6098.4 | This paper | |||||
| SP3(a) (FBR) Optimal values (d) | Searching variables | FL, L/h. | [GLC]inlet, mM | [GLN]inlet, mM | Xv,inlet, Cell/L | This paper | |
| Searching ranges | (10−4–5 × 10−2) | (25–150) | (5–25) | (2 × 108–5 × 109) | |||
| Multi-dimensional optimization | Inlet optimal values of the FBR control variables | ||||||
| FL (b,c) L/h. | [GLC]inlet mM | [GLN]inlet mM | Xv,inlet, Cell/L | Max [mAb](t), (mg/L) | |||
| Time interval (0, 50) h. | 10−3 | 88.65 | 21.58 | 3.21 × 109 | This paper | ||
| Time interval (50, 100) h. | 10−3 | 137.97 | 20.65 | 1.2 × 109 | |||
| Optimal value of Max [mAb](t), (mg/L) | 5700.1 | This paper | |||||
| Bioreactor Operation | Raw Material Consumption (b) | Reactor Performance Max [mAb](t), (b) | FBR Dilution | |||||
|---|---|---|---|---|---|---|---|---|
| Type | Ndiv | Set-Point | Consumed GLC (mmoles) | Consumed GLN (mmoles) | Xv,0 (cells) (c) | (mg/L) | (mg/cells/h) | (%) (a) |
| BR | 1 | Nominal [44] SPBR (d) | 29.1 | 4.9 | 2 × 108 | 1254 | 6.3 × 10−8 | 0 |
| FBR | 5 | Optimal SP1 | 14.22 | 3.23 | ca.4 × 108 | 1351 | 3.4 × 10−8 | 27 |
| FBR | 5 | Optimal SP2 | 44.46 | 8.00 | 1.7 × 109 | 6098 | 3.6 × 10−8 | 10 |
| FBR | 2 | Optimal SP3 | 11.33 | 2.11 | 2.2 × 108 | 5700 | 2.6 × 10−7 | 10 |
| BR | 1 | [54] (e) | 1–5 × 108 | ~1100 | 0 | |||
| FBR | 7–13 | [33] (f) | 2 × (108–109) | ~2400 | ? | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maria, G. Model-Based Optimization of a Fed-Batch Bioreactor for mAb Production Using a Hybridoma Cell Culture. Molecules 2020, 25, 5648. https://doi.org/10.3390/molecules25235648
Maria G. Model-Based Optimization of a Fed-Batch Bioreactor for mAb Production Using a Hybridoma Cell Culture. Molecules. 2020; 25(23):5648. https://doi.org/10.3390/molecules25235648
Chicago/Turabian StyleMaria, Gheorghe. 2020. "Model-Based Optimization of a Fed-Batch Bioreactor for mAb Production Using a Hybridoma Cell Culture" Molecules 25, no. 23: 5648. https://doi.org/10.3390/molecules25235648
APA StyleMaria, G. (2020). Model-Based Optimization of a Fed-Batch Bioreactor for mAb Production Using a Hybridoma Cell Culture. Molecules, 25(23), 5648. https://doi.org/10.3390/molecules25235648