
Fermentation with Edible Rhizopus Strains to Enhance the Bioactive Potential of Hull-Less Pumpkin Oil Cake



Abstract
1. Introduction
2. Results and Discussion
2.1. Antioxidant Activity
2.1.1. Effect of the Processing
2.1.2. The Comparison of the Effect of Individual Strains
2.2. Inositol Phosphates
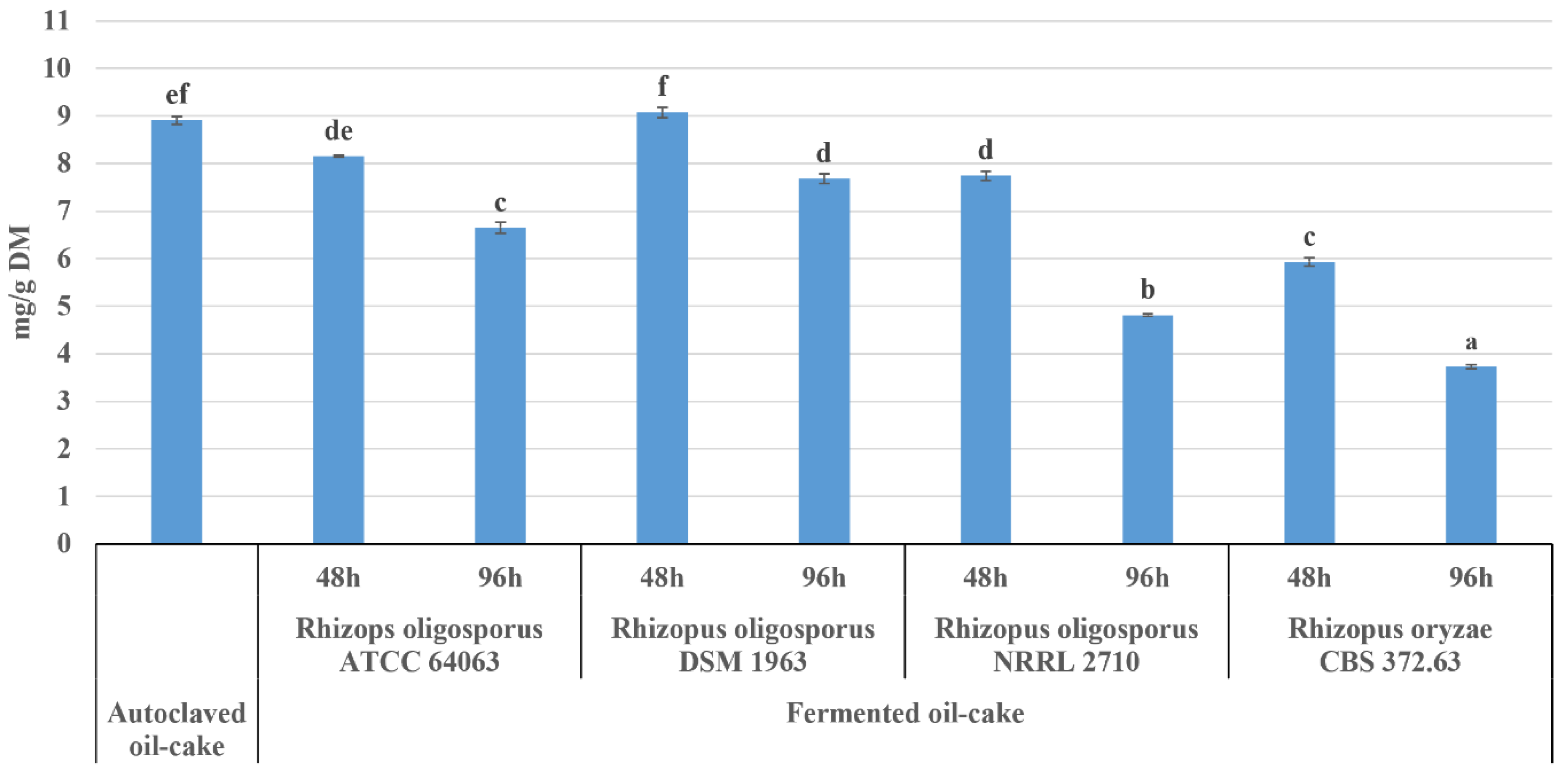
2.2.1. Phytate
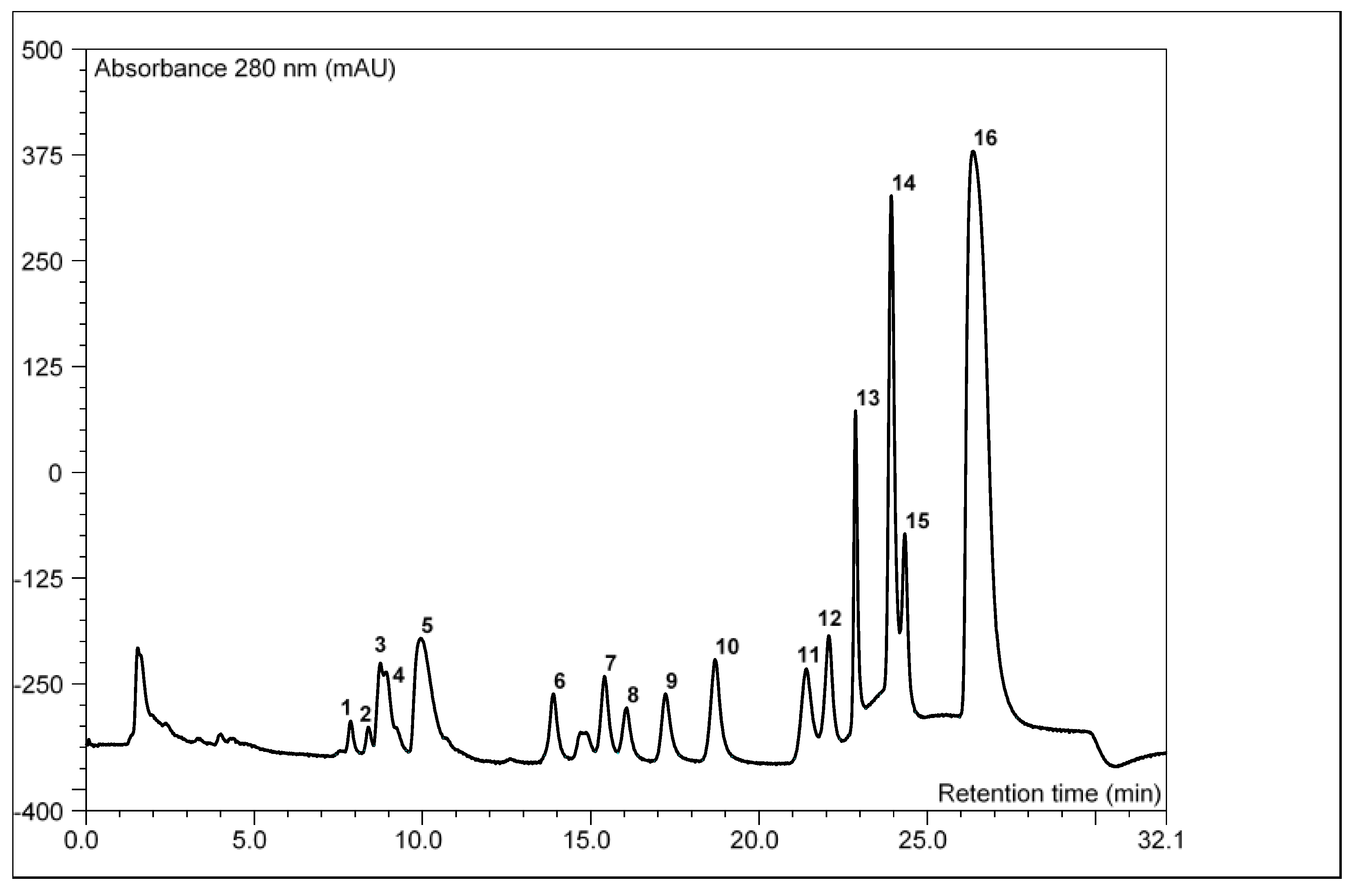
2.2.2. Inositol Phosphate Profile
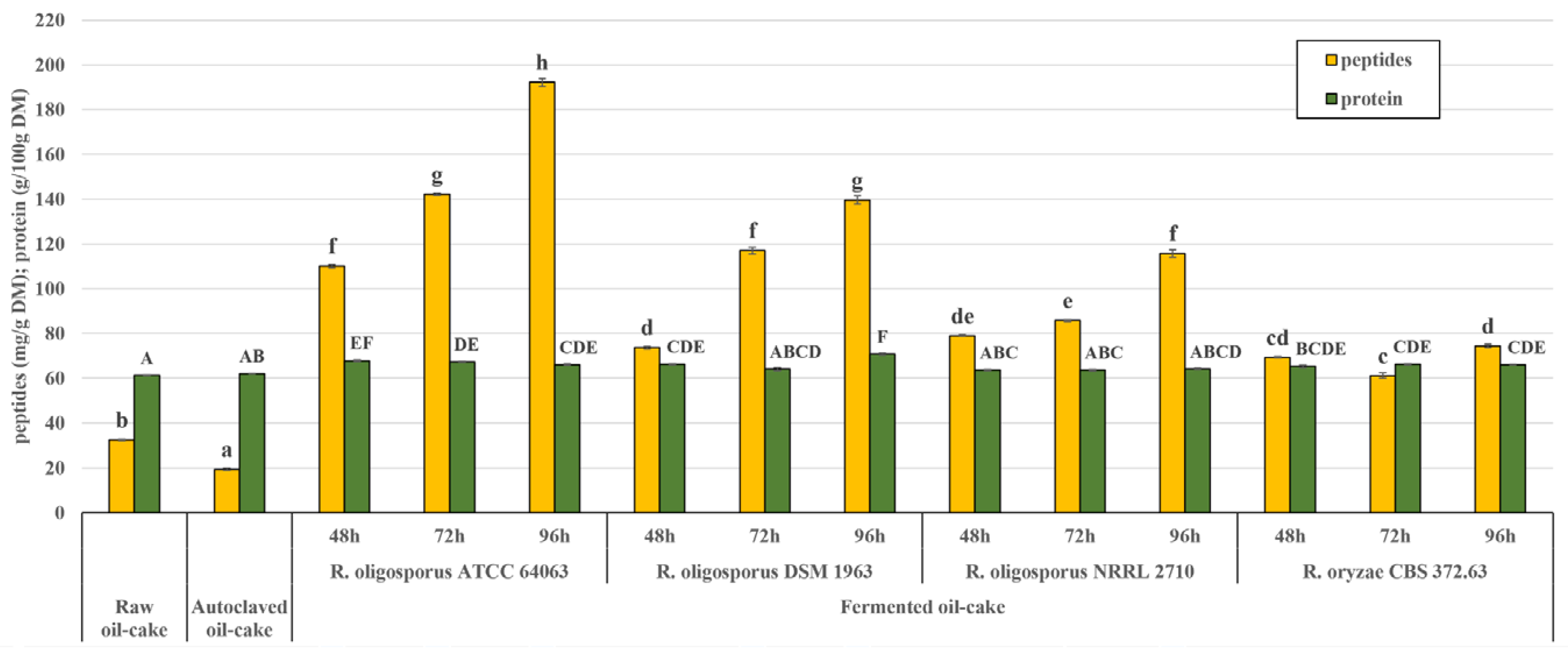
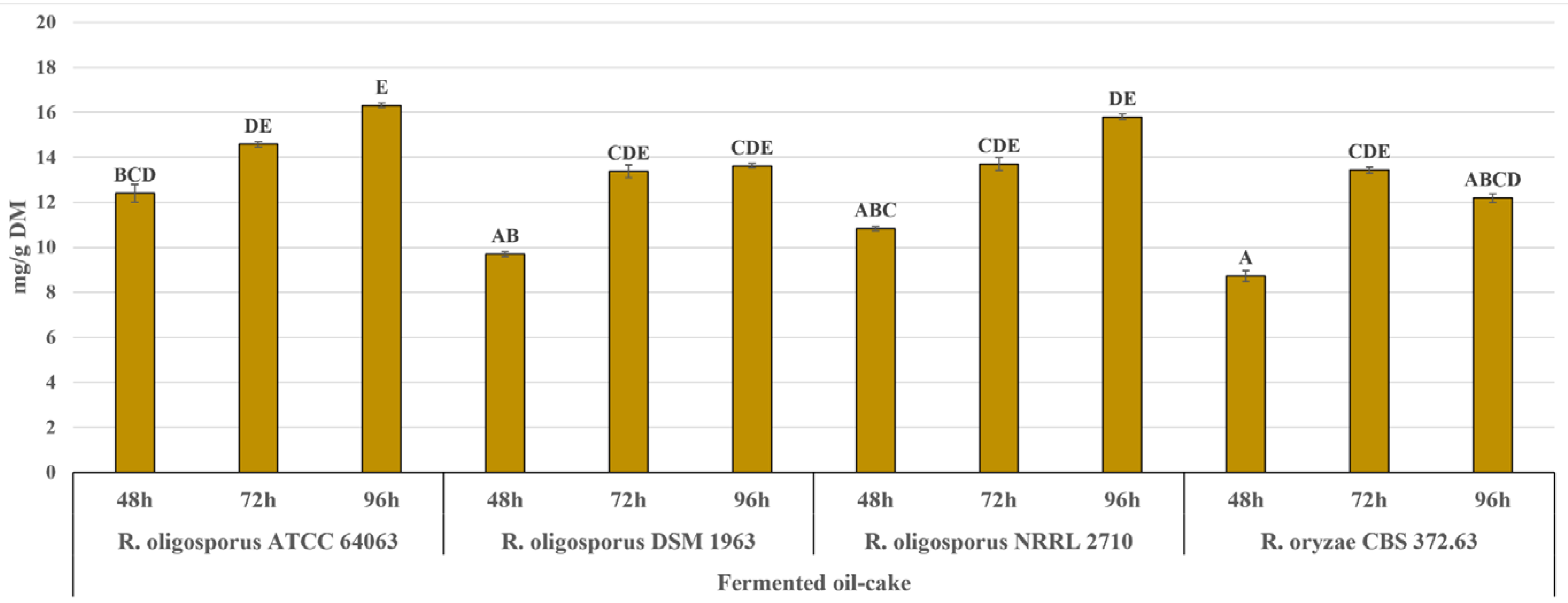
2.3. Protein, Peptide and Glucosamine Levels
3. Materials and Methods
3.1. Materials
3.2. Microorganisms
3.3. Fermentation of Pumpkin Oil Cake
3.4. Analytical Methods
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Janarny, G.; Gunathilake, K.D.P.P. Changes in rice bran bioactives, their bioactivity, bioaccessiblity and bioavailability with solid-state fermentation by Rhizopus oryzae. Biocatal. Agric. Biotechnol. 2020, 23, 1878–1881. [Google Scholar] [CrossRef]
- Owens, J.D.; Astuti, M.; Kuswanto, K.R. Tempe and related products. In Indigenous fermented foods of Southeast Asia; Owens, J.D., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 1–108. [Google Scholar]
- Starzyńska-Janiszewska, A.; Stodolak, B.; Jamróz, M. Antioxidant properties of extracts from fermented and cooked seeds of Polish cultivars of Lathyrus sativus. Food Chem. 2008, 109, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Starzyńska-Janiszewska, A.; Stodolak, B.; Gómez-Caravaca, A.M.; Mickowska, B.; Martin-Garcia, B.; Byczyński, Ł. Mould starter selection for extended solid-state fermentation of quinoa. Lwt-Food Sci. Technol. 2019, 99, 231–237. [Google Scholar] [CrossRef]
- Stodolak, B.; Starzyńska-Janiszewska, A.; Bączkowicz, M. Aspergillus oryzae (Koji Mold) and Neurospora intermedia (Oncom Mold) application for flaxseed oil cake processing. Lwt-Food Sci. Technol. 2020, 131, 109651. [Google Scholar] [CrossRef]
- Popovič, L.; Peričin, D.; Vaštag, Ž.; Popović, S.; Krimer, V.; Torbica, A. Antioxidative and functional properties of pumpkin oil cake globulin hydrolysates. J. Am. Oil Chem. Soc. 2013, 90, 1157–1165. [Google Scholar] [CrossRef]
- Peričin, D.; Krimer, V.; Trivić, S.; Radulović, L. The distribution of phenolic acids in pumpkin’s hull-less seed, skin, oil cake meal, dehulled kernel and hull. Food Chem. 2009, 113, 450–456. [Google Scholar] [CrossRef]
- Zdunczyk, Z.; Minakowski, D.; Frejnagel, S.; Flis, M. Comparative study of the chemical composition and nutritional value of pumpkin seed cake, soybean meal and casein. Nahr.-Food 1999, 43, 392–395. [Google Scholar] [CrossRef]
- Omoruyi, F.O.; Stennett, D.; Foster, S.; Dilworth, L. New frontiers for the use of IP6 and inositol combination in treating diabetes mellitus: A review. Molecules 2020, 25, 1720. [Google Scholar] [CrossRef]
- Atuonwu, A.C.; Akobundu, E.N.T. Nutritional and sensory quality of cookies supplemented with defatted pumpkin (Cucurbita pepo) seed flour. Pak. J. Nutr. 2010, 9, 672–677. [Google Scholar] [CrossRef]
- Popovič, S.; Peričin, D.; Vaštag, Ž.; Popović, L.; Lazić, V. Evaluation of edible film-forming ability of oil cake; effect of pH and temperaturę. Food Hydrocoll. 2011, 25, 470–476. [Google Scholar] [CrossRef]
- Cristiane, R.; Santos, A.; Baretto de Araújo, K.; Soares, C.M.; Lins de Aquino, L.C. Evaluation of temperatue and moisture response surface on the lipase from pumpkin seeds fermentation using Aspergillus niger. Acta Sci. Technol. 2012, 34, 255–260. [Google Scholar] [CrossRef][Green Version]
- Treml, J.; Šmejkal, K. Flavonoids as potent scavengers of hydroxyl radicals. Compr. Rev. Food Sci. Food Saf. 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Parsons, B.J. Antioxidants in food: The significance of characterisation, identification, chemical and biological assays in determining the role of antioxidants in food. Foods 2017, 6, 68. [Google Scholar] [CrossRef]
- Ratz-Łyko, A.; Arct, J. Evaluation of antioxidant and antimicrobial properties of enzymatically hydrolysed Cucurbita pepo and Linum usitatissimum seedcakes. Food Sci. Biotechnol. 2015, 24, 1789–1796. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Li, X. Solvent effects and improvements in the deoxyribose degradation assay for hydroxyl radical-scavenging. Food Chem. 2013, 141, 2083–2088. [Google Scholar] [CrossRef]
- Lipinski, B. Hydroxyl Radical and Its Scavengers in Health and Disease. Oxid. Med. Cell. Longev. 2011, 5. [Google Scholar] [CrossRef]
- Stodolak, B.; Starzyńska-Janiszewska, A.; Wywrocka-Gurgul, A.; Wikiera, A. Solid-state fermented flaxseed oil cake of improved antioxidant capacity as potential. J. Food Process. Preserv. 2016, 41, e12855. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Duliński, R.; Stodolak, B.; Mickowska, B.; Wikiera, A. Prolonged tempe-type fermentation in order to improve bioactive potential and nutritional parameters of quinoa seeds. J. Cereal Sci. 2016, 71, 116–121. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Stodolak, B.; Dulińnski, R.; Bączkowicz, M.; Mickowska, B.; Wikiera, A.; Byczyński, Ł. Effect of solid-state fermentation tempe type on antioxidant and nutritional parameters of buckwheat groats as compared with hydrothermal processing. J. Food Process. Preserv. 2016, 40, 298–305. [Google Scholar] [CrossRef]
- Jayabalan, R.; Subathradevi, P.; Marimuthu, S.; Sathishkumar, M.; Swaminathan, K. Changes in free-radical scavenging ability of kombucha tea during fermentation. Food Chem. 2008, 109, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Berg, S.; Olsson, J.; Swanberg, J.; Schnurer, M.; Eriksson, A. Method for the production of fermented whole grain barley with Rhizopus, and products thereof. European Patent 1,363,505 B1, 1 January 2007. [Google Scholar]
- Londoño-Hernández, L.; Ramírez-Toro, C.; Ruiz, H.A.; Ascacio-Valdés, J.A.; Aguilar-Gonzalez, M.A.; Rodríguez-Herrera, R.; Aguilar, C.N. Rhizopus oryzae—Ancient microbial resource with importance in modern food industry. Int. J. Food Microbiol. 2017, 257, 110–127. [Google Scholar] [CrossRef] [PubMed]
- Simonet, B.M.; Ríos, A.; Grases, F.; Valcárcel Cases, M. Determination of myo-inositol phosphates in food samples by flow injection-capillary zone electrophoresis. Electrophoresis 2003, 24, 2092–2098. [Google Scholar] [CrossRef] [PubMed]
- El-Adawy, T.A.; Taha, K.M. Characteristics and composition of watermelon, pumpkin, and paprika seed oils and flours. J. Agric. Food Chem. 2001, 49, 1253–1259. [Google Scholar] [CrossRef]
- Mansour, E.H.; Dworschák, E.; Lugasi, A.; Barna, É.; Gergely, A. Nutritive value of pumpkin (Cucurbita pepo Kakai 35) seed products. J. Sci. Food Agric. 1993, 61, 73–78. [Google Scholar] [CrossRef]
- Duliński, R.; Stodolak, B.; Byczyński, Ł.; Poreda, A.; Starzyńska-Janiszewska, A.; Żyła, K. Solid-state fermentation reduces phytic acid level, improves the profile of myo-inositol phosphates and enhances the availability of selected minerals in flaxseed oil cake. Food Technol. Biotechnol. 2017, 55, 413–419. [Google Scholar] [CrossRef]
- Ramachandran, S.; Krishnankutty, R.; Nampoothiri, K.M.; Szakacs, G.; Pandey, A. Mixed substrate fermentation for the production of phytase by Rhizopus spp. using oilcakes as substrates. Process. Biochem. 2005, 40, 1749–1754. [Google Scholar] [CrossRef]
- Rani, R.; Ghosh, S. Production of phytase under solid-state fermentation using Rhizopus oryzae: Novel strain improvement approach and studies on purification and characterization. Bioresour. Technol. 2011, 102, 10641–10649. [Google Scholar] [CrossRef]
- Vucenik, I.; Shamsuddin, A.M. Cancer inhibition by inositol hexaphosphate (IP6) and inositol: From laboratory to clinic. J. Nutr. 2003, 133, 3778S–3784S. [Google Scholar] [CrossRef]
- Vig, A.P.; Walia, A. Beneficial effects of Rhizopus oligosporus fermentation on reduction of glucosinolates, fibre and phytic acid in rapeseed (Brassica napus) meal. Bioresour. Technol. 2001, 78, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Kadam, D.; Lele, S.S. Value addition of oilseed meal: A focus on bioactive peptides. J. Food Meas. Charact. 2018, 12, 449–458. [Google Scholar] [CrossRef]
- Vaštag, Ž.; Popović, L.; Popović, S.; Krimer, V.; Peričin, D. Hydrolysis of pumpkin oil cake protein isolate and free radical scavenging activity of hydrolysates: Influence of temperature, enzyme/substrate ratio and time. Food Bioprod. Process. 2010, 88, 277–282. [Google Scholar] [CrossRef]
- Zheng, Z.; Shetty, K. Cranberry processing waste for solid state fungal inoculants production. Process. Biochem. 1998, 33, 323–329. [Google Scholar] [CrossRef]
- Gharieb, M.M.; El-Sabbagh, S.; Shalaby, M.A.; Darwesh, O.M. Production of chitosan from different species of Zygomycetes and its antimicrobial activity. Int. J. Sci. Eng. Res. 2015, 6, 123–130. [Google Scholar]
- Gutiérrez, T.J. Chitosan Applications for the Food Industry. In Chitosan: Derivatives, Composites and Applications; John Wiley and Sons: Hoboken, NJ, USA, 2017; pp. 183–232. [Google Scholar] [CrossRef]
- Swain, T.; Hillis, W.E. The phenolic constituents of Prunus domestica L. The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 1959, 10, 63–68. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. Hydrophilic and lipophilic antioxidant activity changes during on-vine ripening of tomatoes (Lycopersicon esculentum Mill.). Postharvest Biol. Technol 2007, 28, 59–65. [Google Scholar] [CrossRef]
- Gökmen, V.; Serpen, A.; Fogliano, V. Direct measurement of the total antioxidant capacity of foods: The “QUENCHER” approach. Trends Food Sci. Technol. 2009, 20, 278–288. [Google Scholar] [CrossRef]
- Marambe, P.W.M.L.H.K.; Shand, P.J.; Wanasundara, J.P.D. An in-vitro investigation of selected biological activities of hydrolysed flaxseed (Linum usitatissimum L.) proteins. J. Am. Oil Chem. Soc. 2008, 85, 1155–1164. [Google Scholar] [CrossRef]
- Ardestani, A.; Yazdanparast, R. Antioxidant and free radical scavenging potential of Achillea santolina extracts. Food Chem. 2007, 104, 21–29. [Google Scholar] [CrossRef]
- Duliński, R.; Cielecka, E.K.; Pierzchalska, M.; Żyła, K. Phytases improve myo-inositol bioaccessibility in rye bread: A study using an in vitro method of digestion and a caco-2 cell culture model. Food Technol. Biotechnol. 2015, 53, 66–72. [Google Scholar] [CrossRef]
- Blaabjerg, K.; Hansen-Møller, J.; Poulsen, H.D. High-performance ion chromatography method for separation and quantification of inositol phosphates in diets and digesta. J. Chromatogr. B 2010, 878, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Dionex—Thermo Fisher Scientific Inc. Application Note 65: Analysis of Inositol Phosphates. 2003. Available online: http://tools.thermofisher.com/content/sfs/brochures/4663-AN65_LC_insP3_2003_LPN034407-01.pdf (accessed on 1 September 2019).
- Marczenko, Z.; Balcerzak, M. Nitrogen. In Spectrophotometric Methods in Inorganic Analysis; Galus, M., Ed.; PWN: Warsaw, Poland, 1998; pp. 102–112. [Google Scholar]
- Zhu, Y.P.; Cheng, Y.Q.; Wang, L.J.; Fan, J.F.; Li, L.T. Enhanced antioxidative activity of Chinese traditionally fermented okara (meitauza) prepared with various microorganism. Int. J. Food Prop. 2008, 11, 519–529. [Google Scholar] [CrossRef]
- Desgranges, C.; Vergoignan, C.; Georges, M.; Durand, A. Biomass estimation in solid state fermentation I. Manual biochemical methods. Appl. Microbiol. Biotechnol. 1991, 35, 200–205. [Google Scholar] [CrossRef]
- Tsuji, A.; Kinishita, T.; Hoshino, M. Analytical chemical studies on amino sugars. II. Determination of hexosamines using 3-methyl-2-benzothizolone hydrazone hydrochloride. Chem. Pharm. Bull. (Tokyo) 1969, 17, 1505–1510. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FCRS (mg/g DM) | SA-ABTS˙+ (µmol Trolox/g DM) | QA-ABTS˙+ (µmol Trolox/g DM) | SA-OH˙ (EC50) | Reducing Power (RP0.5) | |||
|---|---|---|---|---|---|---|---|
| Raw oil-cake | 3.33 ± 0.09 a | 18.25 ± 0.42 a | 44.48 ± 0.77 a | 1.52 ± 0.02 g | 23.62 ± 0.51 f | ||
| Autoclaved oil-cake | 4.79 ± 0.08 b | 32.07 ± 0.22 b | 43.94 ± 1.22 a | 0.42 ± 0.02 a | 25.72 ± 0.60 g | ||
| Fermented oil-cake | Strain | Time | |||||
| Rhizopus oligosporus ATCC 64063 | 48 h | 9.74 ± 0.05 f | 67.16 ± 0.67 i | 61.27 ± 1.20 de | 0.51 ± 0.06 ab | 5.74 ± 0.11 a | |
| 72 h | 10.14 ± 0.16 f | 66.19 ± 0.80 i | 64.61 ± 1.13 e | 0.77 ± 0.05 cd | 10.41 ± 0.16 e | ||
| 96 h | 11.22 ± 0.13 g | 68.34 ± 2.55 i | 64.80 ± 0.33 e | 1.01 ± 0.04 ef | 9.90 ± 0.13 de | ||
| Rhizopus oligosporus DSM 1964 | 48 h | 8.14 ± 0.13 cd | 55.22 ± 1.28 ef | 57.43 ± 0.98 bcd | 0.67 ± 0.02 bc | 7.42 ± 0.09 b | |
| 72 h | 8.55 ± 0.07 d | 60.65 ± 0.50 h | 57.81 ± 0.71 bcd | 0.94 ± 0.02 def | 9.57 ± 0.15 d | ||
| 96 h | 9.62 ± 0.10 ef | 59.66 ± 0.85 gh | 64.15 ± 1.61 e | 0.91 ± 0.05 de | 10.14 ± 0.15 de | ||
| Rhizopus oligosporus NRRL 2710 | 48 h | 8.25 ± 0.01 cd | 56.81 ± 1.81 fg | 60.84 ± 0.38 cde | 0.78 ± 0.03 cd | 8.45 ± 0.12 c | |
| 72 h | 8.24 ± 0.12 cd | 52.74 ± 0.70 de | 58.22 ± 1.14 bcd | 1.13 ± 0.01 f | 8.27 ± 0.15 c | ||
| 96 h | 9.14 ± 0.11 e | 59.44 ± 0.25 gh | 61.53 ± 1.85 de | 1.04 ± 0.07 ef | 8.46 ± 0.22 c | ||
| Rhizopus oryzae CBS 372.63 | 48 h | 7.81 ± 0.11 c | 50.40 ± 0.93 d | 55.37 ± 0.84 bc | 0.90 ± 0.02 de | 8.46 ± 0.22 c | |
| 72 h | 8.06 ± 0.16 cd | 43.05 ± 1.58 c | 54.65 ± 1.03 b | 1.05 ± 0.07 ef | 8.79 ± 0.11 c | ||
| 96 h | 7.96 ± 0.14 c | 45.24 ± 0.19 c | 56.72 ± 0.43 bcd | 0.66 ± 0.05 bc | 8.43 ± 0.22 c | ||
| InsP1–2 | InsP3 | InsP4 | InsP5 | InsP6 | |||
|---|---|---|---|---|---|---|---|
| Autoclaved oil-cake | 1.32 | 4.74 | 6.11 | 18.98 | 68.84 | ||
| Fermented oil-cake | Strain | time | |||||
| Rhizopus oligosporus ATCC 64063 | 48 h | 2.17 | 8.95 | 9.67 | 17.89 | 61.32 | |
| 96 h | 0.83 | 6.02 | 11.27 | 23.56 | 58.34 | ||
| Rhizopus oligosporus DSM 1964 | 48 h | 0.43 | 13.81 | 16.89 | 24.06 | 44.79 | |
| 96 h | 1.04 | 5.45 | 7.1 | 19.61 | 66.78 | ||
| Rhizopus oligosporus NRRL 2710 | 48 h | 0.98 | 27.25 | 8.8 | 6.24 | 56.72 | |
| 96 h | n.i. | 4.23 | 2.15 | 14.87 | 78.75 | ||
| Rhizopus oryzae CBS 372.63 | 48 h | 3.33 | 17.87 | 40.01 | 18.63 | 20.17 | |
| 96 h | 1.97 | 12.92 | 37.08 | 21.54 | 26.47 | ||
Sample Availability: Samples of the compounds are not available from the authors. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starzyńska-Janiszewska, A.; Duliński, R.; Stodolak, B. Fermentation with Edible Rhizopus Strains to Enhance the Bioactive Potential of Hull-Less Pumpkin Oil Cake. Molecules 2020, 25, 5782. https://doi.org/10.3390/molecules25245782
Starzyńska-Janiszewska A, Duliński R, Stodolak B. Fermentation with Edible Rhizopus Strains to Enhance the Bioactive Potential of Hull-Less Pumpkin Oil Cake. Molecules. 2020; 25(24):5782. https://doi.org/10.3390/molecules25245782
Chicago/Turabian StyleStarzyńska-Janiszewska, Anna, Robert Duliński, and Bożena Stodolak. 2020. "Fermentation with Edible Rhizopus Strains to Enhance the Bioactive Potential of Hull-Less Pumpkin Oil Cake" Molecules 25, no. 24: 5782. https://doi.org/10.3390/molecules25245782
APA StyleStarzyńska-Janiszewska, A., Duliński, R., & Stodolak, B. (2020). Fermentation with Edible Rhizopus Strains to Enhance the Bioactive Potential of Hull-Less Pumpkin Oil Cake. Molecules, 25(24), 5782. https://doi.org/10.3390/molecules25245782