
Coixol Suppresses NF-κB, MAPK Pathways and NLRP3 Inflammasome Activation in Lipopolysaccharide-Induced RAW 264.7 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
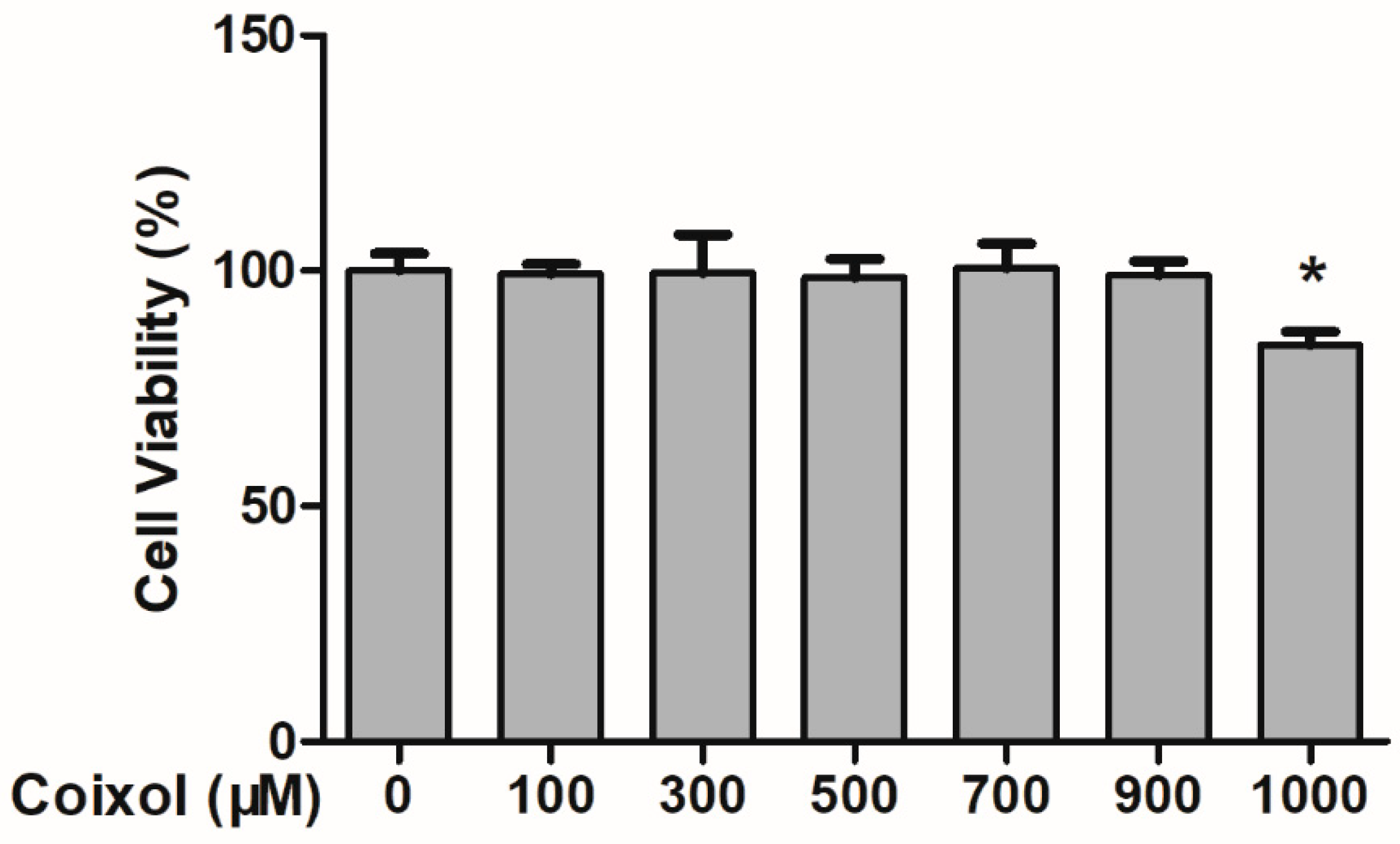
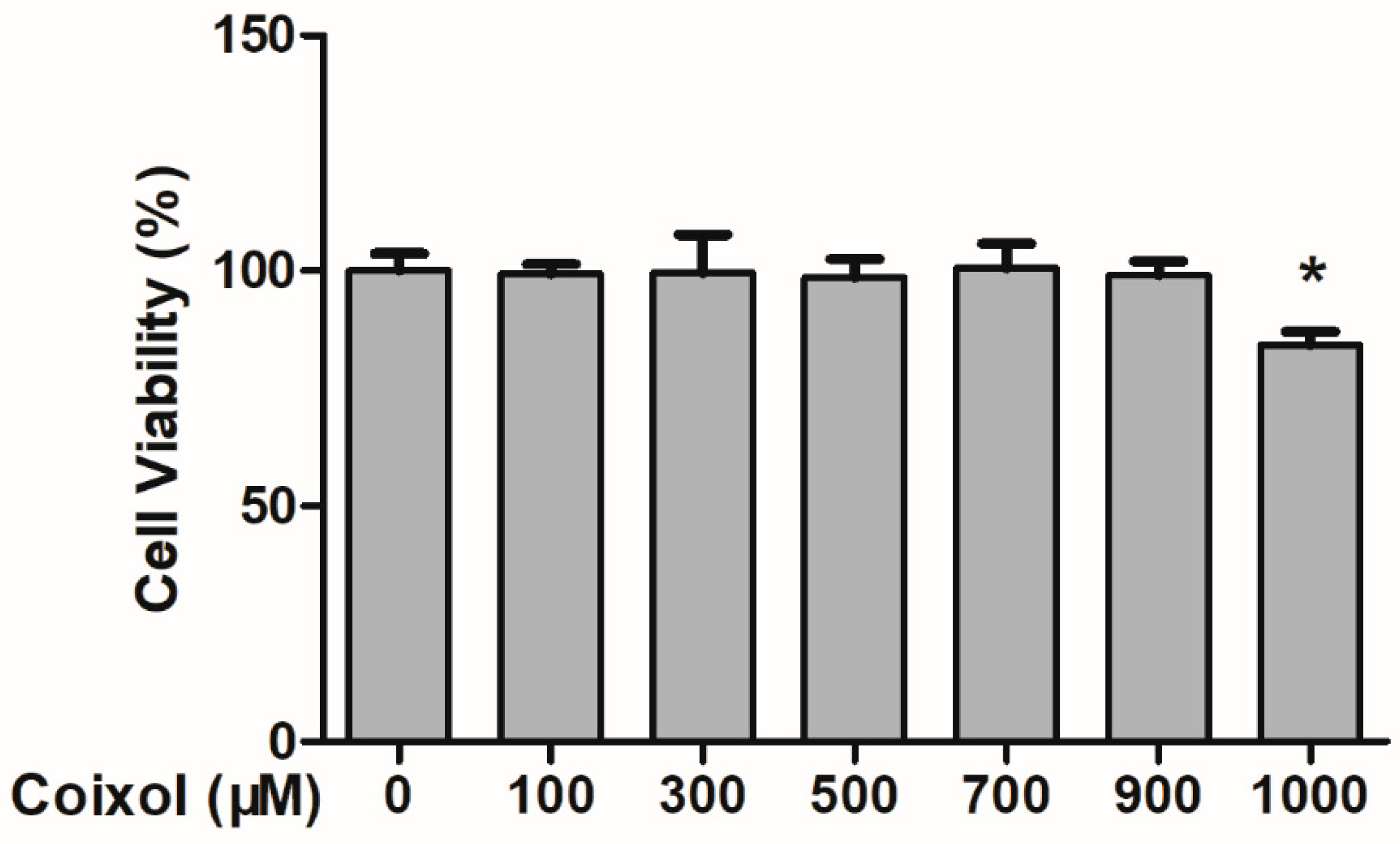
2.1. Cytotoxic Effects of Coixol on RAW 264.7 Cells
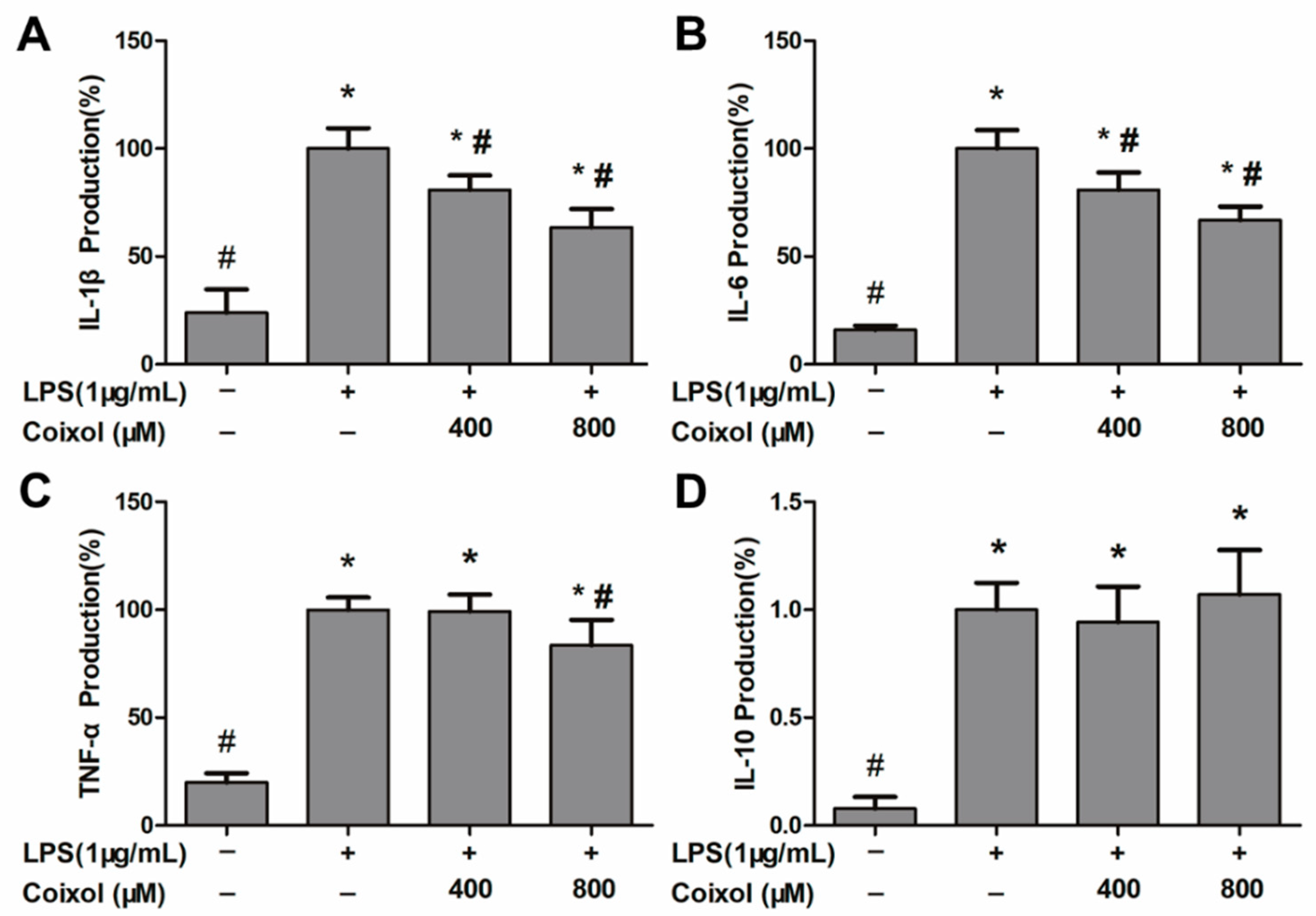
2.2. Production of IL-1β, IL-6, TNF-α, and IL-10
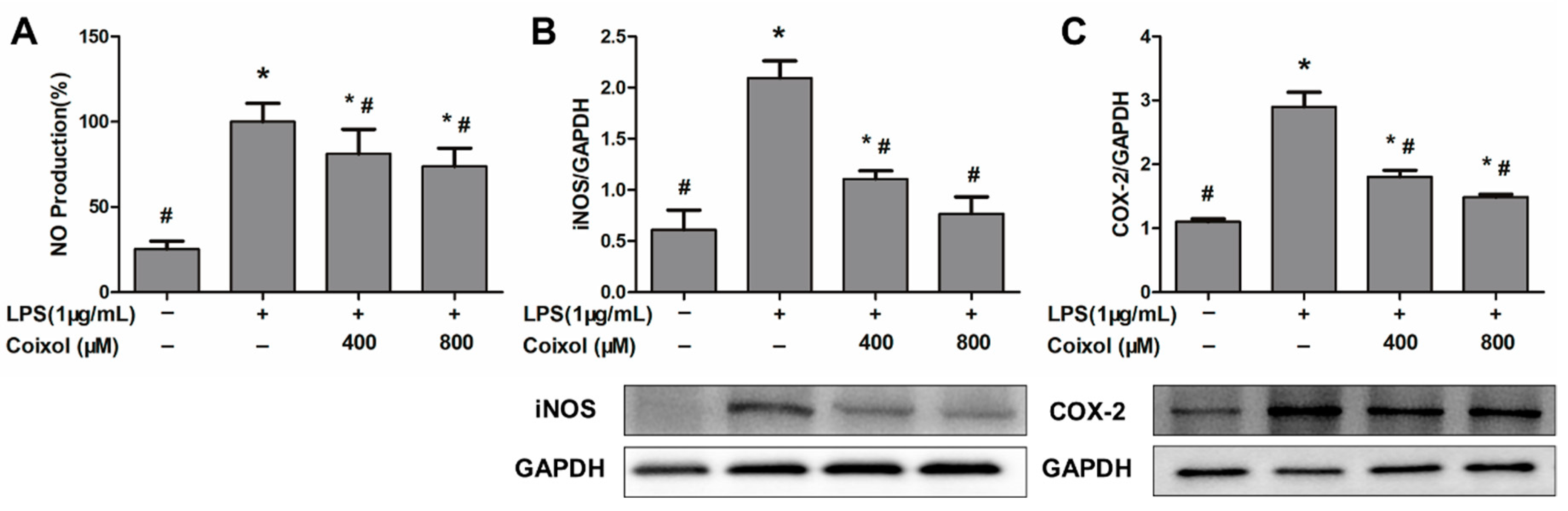
2.3. Expression of NO, iNOS, and COX-2
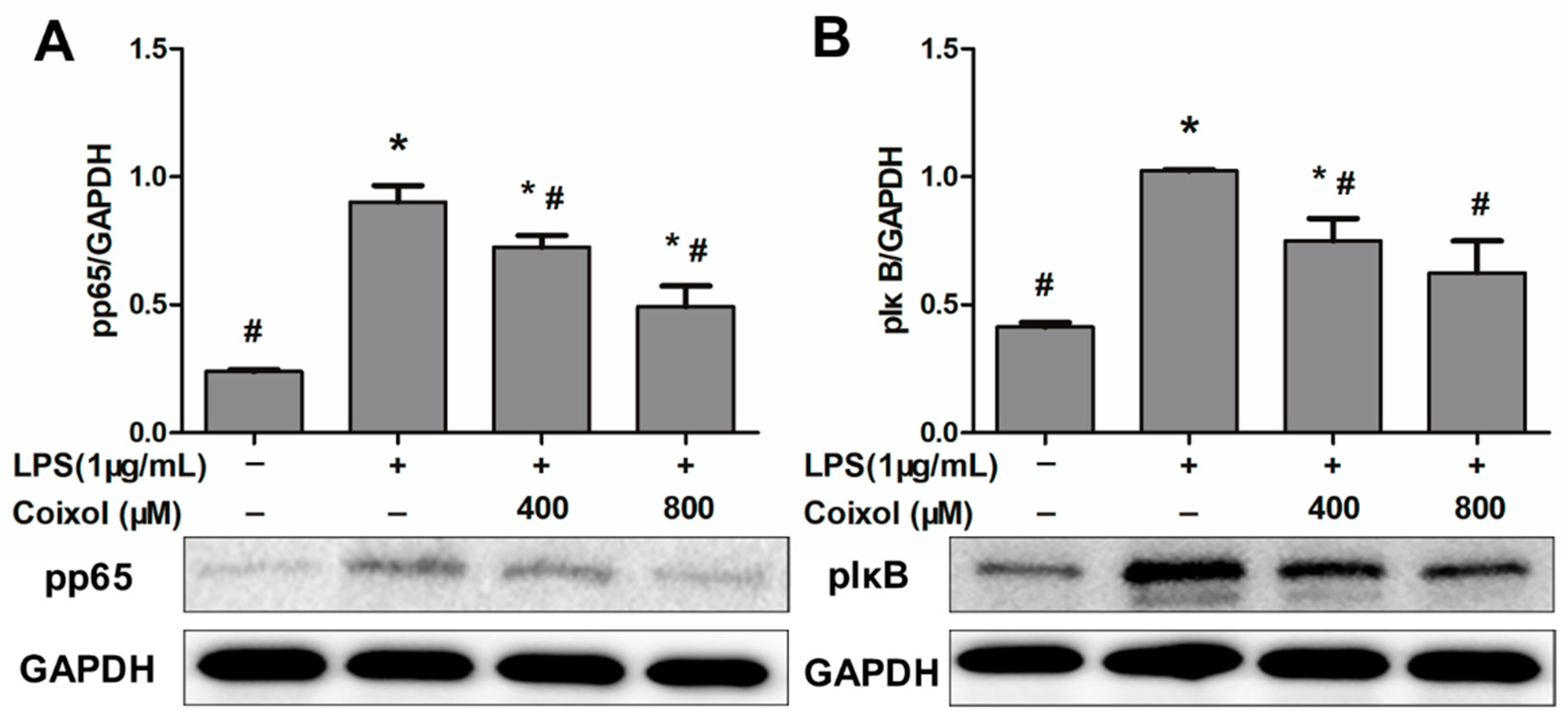
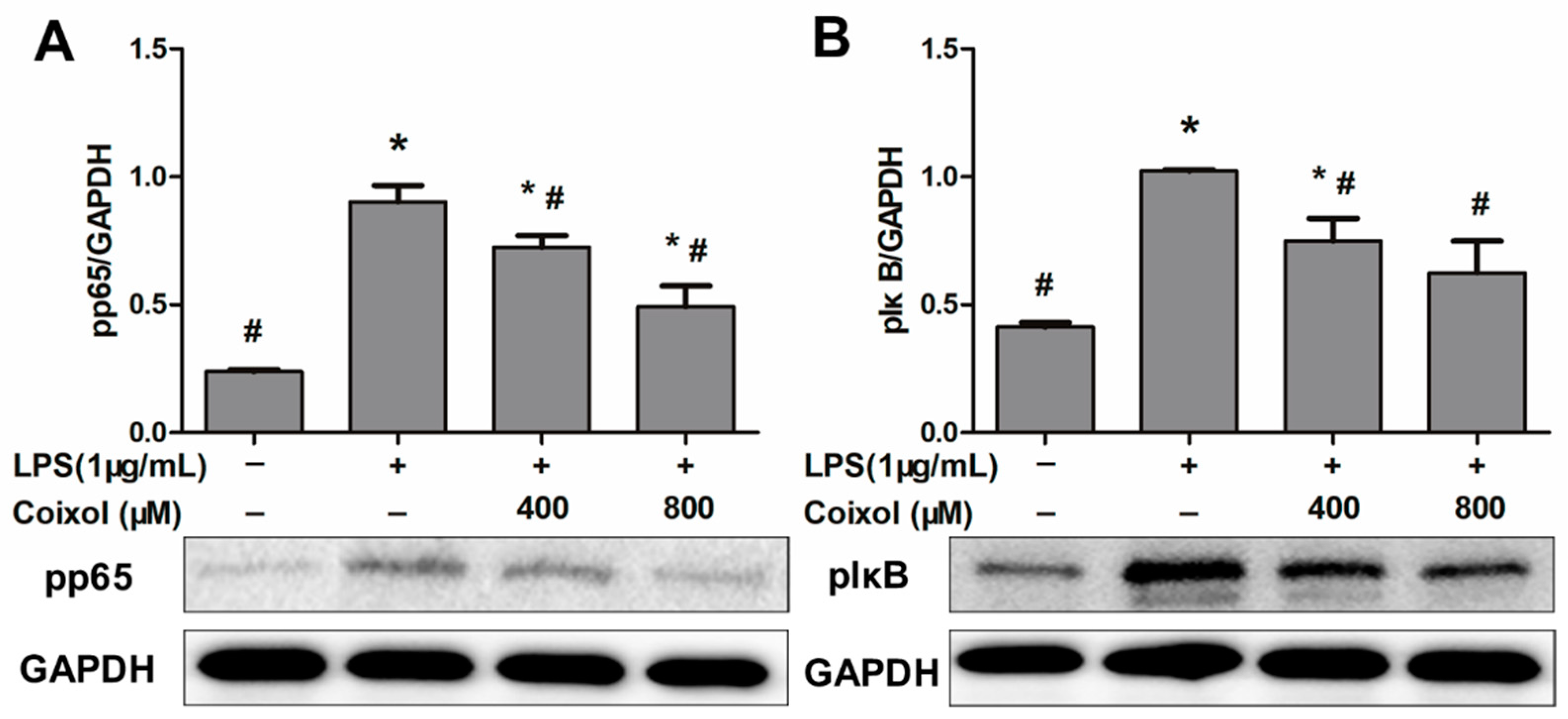
2.4. Expression of NF-κB Pathways
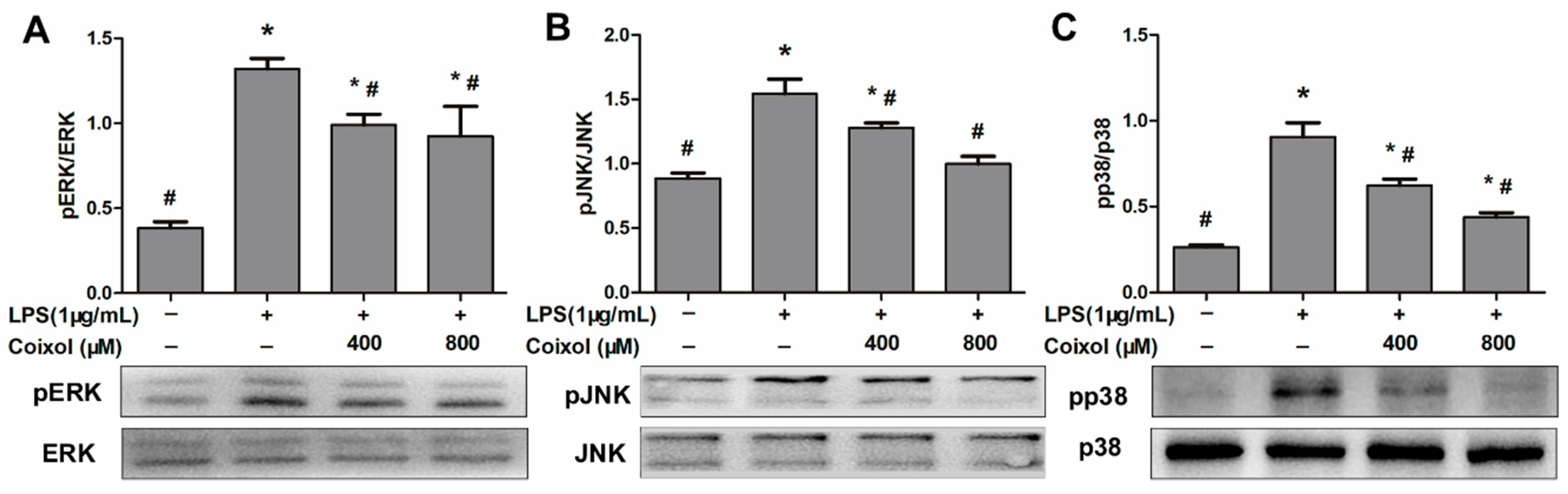
2.5. Expression of MAPK Pathways
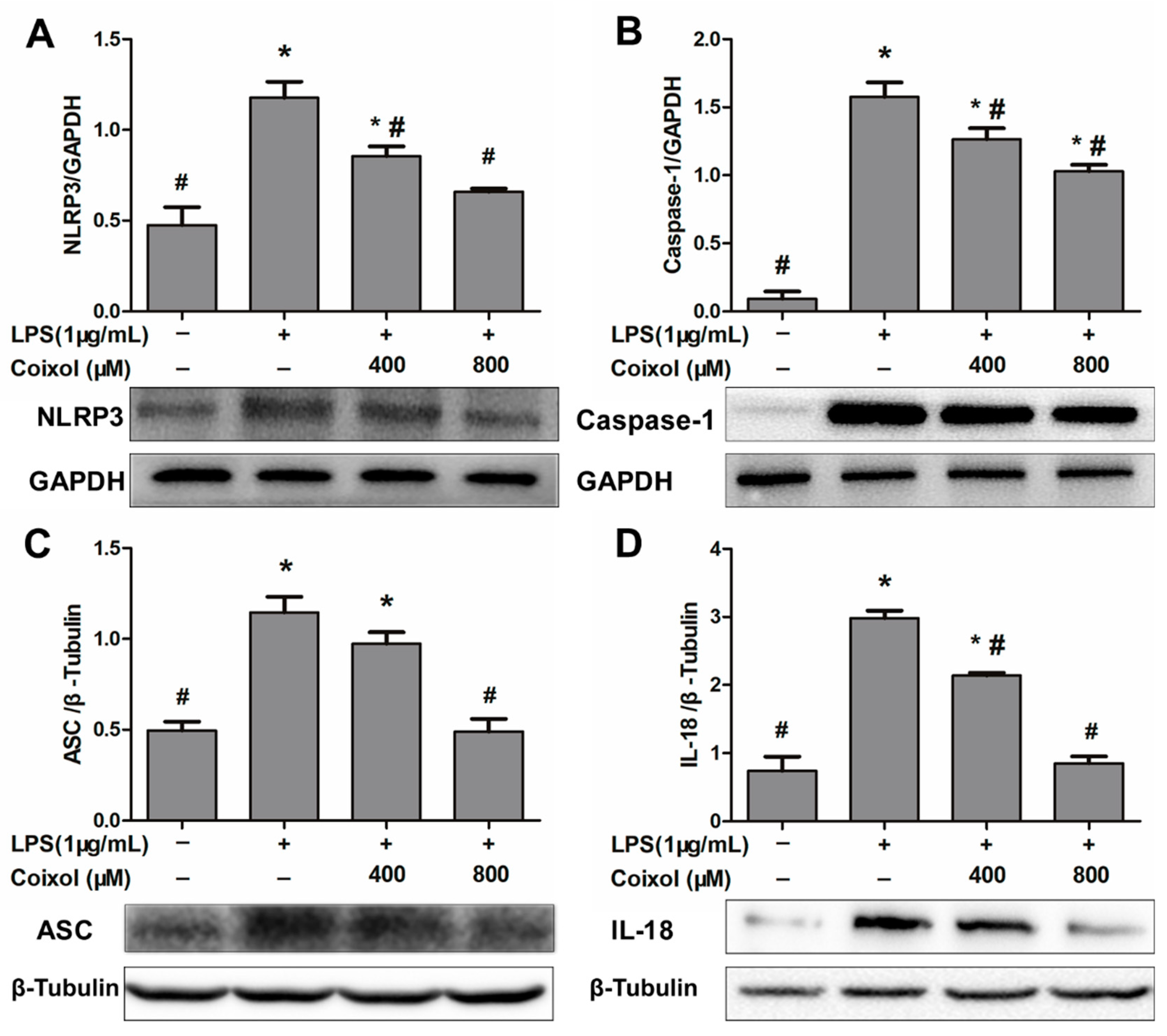
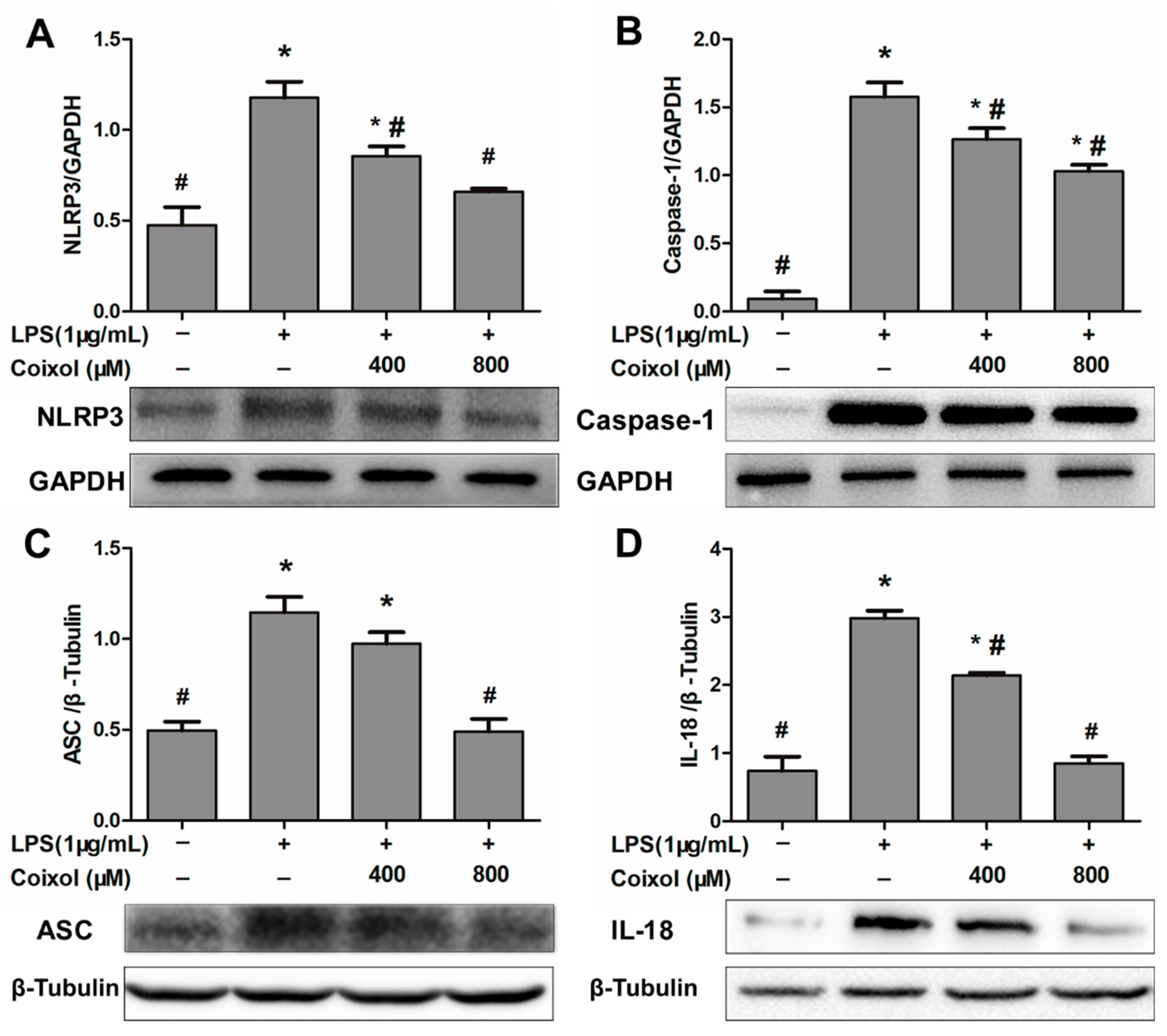
2.6. Expression of NLRP3 Inflammasome
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Viability Assay
4.3. Determination of NO Production
4.4. Determination of IL-1β, IL-6, TNF-α and IL-10 Production
4.5. Western Blotting
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. Mechanisms of disease the pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.G.; Guzik, T.J.; Lob, H.E.; Madhur, M.S.; Marvar, P.J.; Thabet, S.R.; Vinh, A.; Weyand, C.M. Inflammation, Immunity, and Hypertension. Hypertension 2011, 57, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.H.; Raison, C.L. The role of inflammation in depression: From evolutionary imperative to modern treatment target. Nat. Rev. Immunol. 2016, 16, 22–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olefsky, J.M.; Glass, C.K. Macrophages, Inflammation, and Insulin Resistance. Annu. Rev. Physiol. 2010, 72, 219–246. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, Inflammation, and Cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Lawrence, T. The Nuclear Factor NF-kappa B Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta-Mol. Basis Dis. 2010, 1802, 396–405. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Hara, H.; Nunez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Arora, R.K. Job’s-tears (coix lacryma-jobi)—A minor food and fodder crop of northeastern India. Econ. Bot. 1977, 31, 358–366. [Google Scholar] [CrossRef]
- Wrigley, C.W.; Corke, H.; Seetharaman, K.; Faubion, J. Coix: Overview; Elsevier: Oxford, UK, 2016; pp. 184–189. [Google Scholar]
- Huang, C.J.; Hou, M.F.; Kan, J.Y.; Juan, C.H.; Yuan, S.S.F.; Luo, K.H.; Chuang, H.Y.; Hu, S.C.S. Prophylactic Treatment with Adlay Bran Extract Reduces the Risk of Severe Acute Radiation Dermatitis: A Prospective, Randomized, Double-Blind Study. Evid. Based Complement. Altern. Med. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Y.P.; Huang, X.E.; Cao, J.; Lu, Y.Y.; Wu, X.Y.; Liu, J.; Xu, X.; Xiang, J.; Ye, L.H. Clinical Safety and Efficacy of Kanglaite (R) (Coix Seed Oil) Injection Combined with Chemotherapy in Treating Patients with Gastric Cancer. Asian Pac. J. Cancer Prev. 2012, 13, 5319–5321. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F. Coix: Chemical composition and health effects. Trends Food Sci. Technol. 2017, 61, 160–175. [Google Scholar] [CrossRef]
- Choi, G.; Han, A.R.; Lee, J.H.; Park, J.Y.; Kang, U.; Hong, J.; Kim, Y.S.; Seo, E.K. A Comparative Study on Hulled Adlay and Unhulled Adlay through Evaluation of Their LPS-Induced Anti-Inflammatory Effects, and Isolation of Pure Compounds. Chem. Biodivers. 2015, 12, 380–387. [Google Scholar] [CrossRef]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef] [PubMed]
- Sanders, E.H.; Gardner, P.D.; Berger, P.J.; Negus, N.C. 6-methoxybenzoxazolinone—A plant derivative that stimulates reproduction in microtus-montanus. Science 1981, 214, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Yuwiler, A.; Winters, W.D. Effects of 6-methoxy-2-benzoxazolinone on the pineal melatonin generating-system. J. Pharm. Exp. 1985, 233, 45–50. [Google Scholar]
- Chung, C.P.; Hsu, C.Y.; Lin, J.H.; Kuo, Y.H.; Chiang, W.C.; Lin, Y.L. Antiproliferative Lactams and Spiroenone from Adlay Bran in Human Breast Cancer Cell Lines. J. Agric. Food Chem. 2011, 59, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Liu, F.; Ng, T.B. Examination of pineal indoles and 6-methoxy-2-benzoxazolinone for antioxidant and antimicrobial effects. Comp. Biochem. Physiol. C-Toxicol. Pharm. 2001, 130, 379–388. [Google Scholar] [CrossRef]
- Lee, H.J.; Ryu, J.; Park, S.H.; Seo, E.K.; Han, A.R.; Lee, S.K.; Kim, Y.S.; Hong, J.H.; Seok, J.H.; Lee, C.J. Suppressive effects of coixol, glyceryl trilinoleate and natural products derived from Coix Lachryma-Jobi var. ma-yuen on gene expression, production and secretion of airway MUC5AC mucinn. Arch. Pharm. Res. 2015, 38, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Hafizur, R.M.; Khan, M.I.; Jawed, A.; Wang, H.; Zhao, M.M.; Matsunaga, K.; Izumi, T.; Siddiqui, S.; Khan, F.; et al. Coixol amplifies glucose-stimulated insulin secretion via cAMP mediated signaling pathway. Eur. J. Pharm. 2019, 858, 11. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Wu, C.H.; Shih, C.K.; Liu, C.Y.; Shih, P.H.; Shieh, T.M.; Lin, C.I.; Chiang, W.C.; Hsia, S.M. Application of the solvent extraction technique to investigation of the anti-inflammatory activity of adlay bran. Food Chem. 2014, 145, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, M.L.; Jin, S.L.C.; Milla, M.E.; Burkhart, W.; Carter, H.L.; Chen, W.J.; Clay, W.C.; Didsbury, J.R.; Hassler, D.; Hoffman, C.R.; et al. Cloning of a disintegrin metalloproteinase that processes precursor tumour-necrosis factor-alpha. Nature 1997, 385, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; DuBois, R.N. The role of COX-2 in intestinal inflammation and colorectal cancer. Oncogene 2010, 29, 781–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawabata, A. Prostaglandin E-2 and Pain-An Update. Biol. Pharm. Bull. 2011, 34, 1170–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrijvers, E.M.C.; Witteman, J.C.M.; Sijbrands, E.J.G.; Hofman, A.; Koudstaal, P.J.; Breteler, M.M.B. Insulin metabolism and the risk of Alzheimer disease The Rotterdam Study. Neurology 2010, 75, 1982–1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reaven, G.M. Insulin Resistance: The Link Between Obesity and Cardiovascular Disease. Med. Clin. N. Am. 2011, 95, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Vandanmagsar, B.; Youm, Y.H.; Ravussin, A.; Galgani, J.E.; Stadler, K.; Mynatt, R.L.; Ravussin, E.; Stephens, J.M.; Dixit, V.D. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat. Med. 2011, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Zhou, Q.; Liu, T.; Liu, Z. Coixol Suppresses NF-κB, MAPK Pathways and NLRP3 Inflammasome Activation in Lipopolysaccharide-Induced RAW 264.7 Cells. Molecules 2020, 25, 894. https://doi.org/10.3390/molecules25040894
Hu Y, Zhou Q, Liu T, Liu Z. Coixol Suppresses NF-κB, MAPK Pathways and NLRP3 Inflammasome Activation in Lipopolysaccharide-Induced RAW 264.7 Cells. Molecules. 2020; 25(4):894. https://doi.org/10.3390/molecules25040894
Chicago/Turabian StyleHu, Yusheng, Qilyu Zhou, Tianlong Liu, and Zhongjie Liu. 2020. "Coixol Suppresses NF-κB, MAPK Pathways and NLRP3 Inflammasome Activation in Lipopolysaccharide-Induced RAW 264.7 Cells" Molecules 25, no. 4: 894. https://doi.org/10.3390/molecules25040894
APA StyleHu, Y., Zhou, Q., Liu, T., & Liu, Z. (2020). Coixol Suppresses NF-κB, MAPK Pathways and NLRP3 Inflammasome Activation in Lipopolysaccharide-Induced RAW 264.7 Cells. Molecules, 25(4), 894. https://doi.org/10.3390/molecules25040894