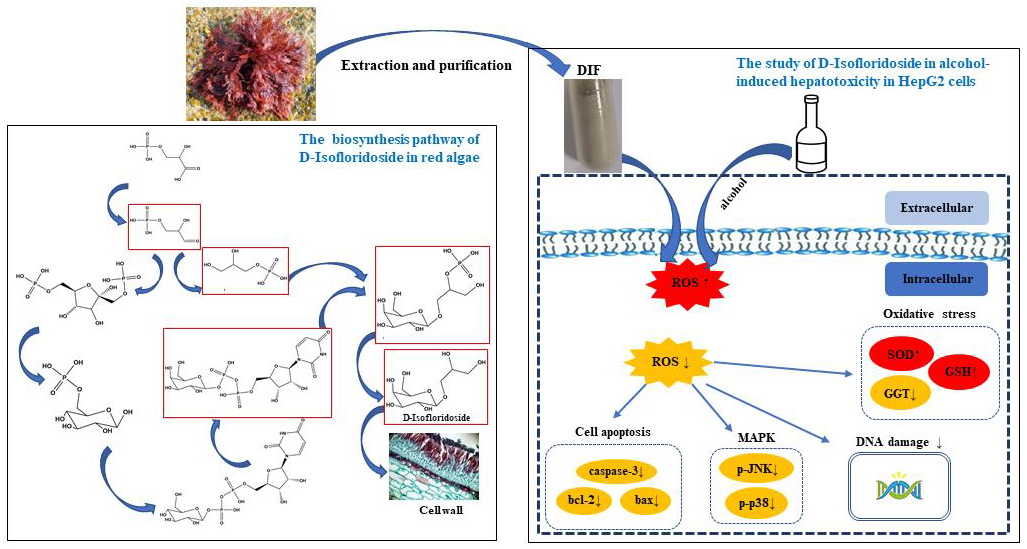
The Protective Effect of the Polysaccharide Precursor, D-Isofloridoside, from Laurencia undulata on Alcohol-Induced Hepatotoxicity in HepG2 Cells

 ,
,  and
and
Abstract
:
1. Introduction
2. Results
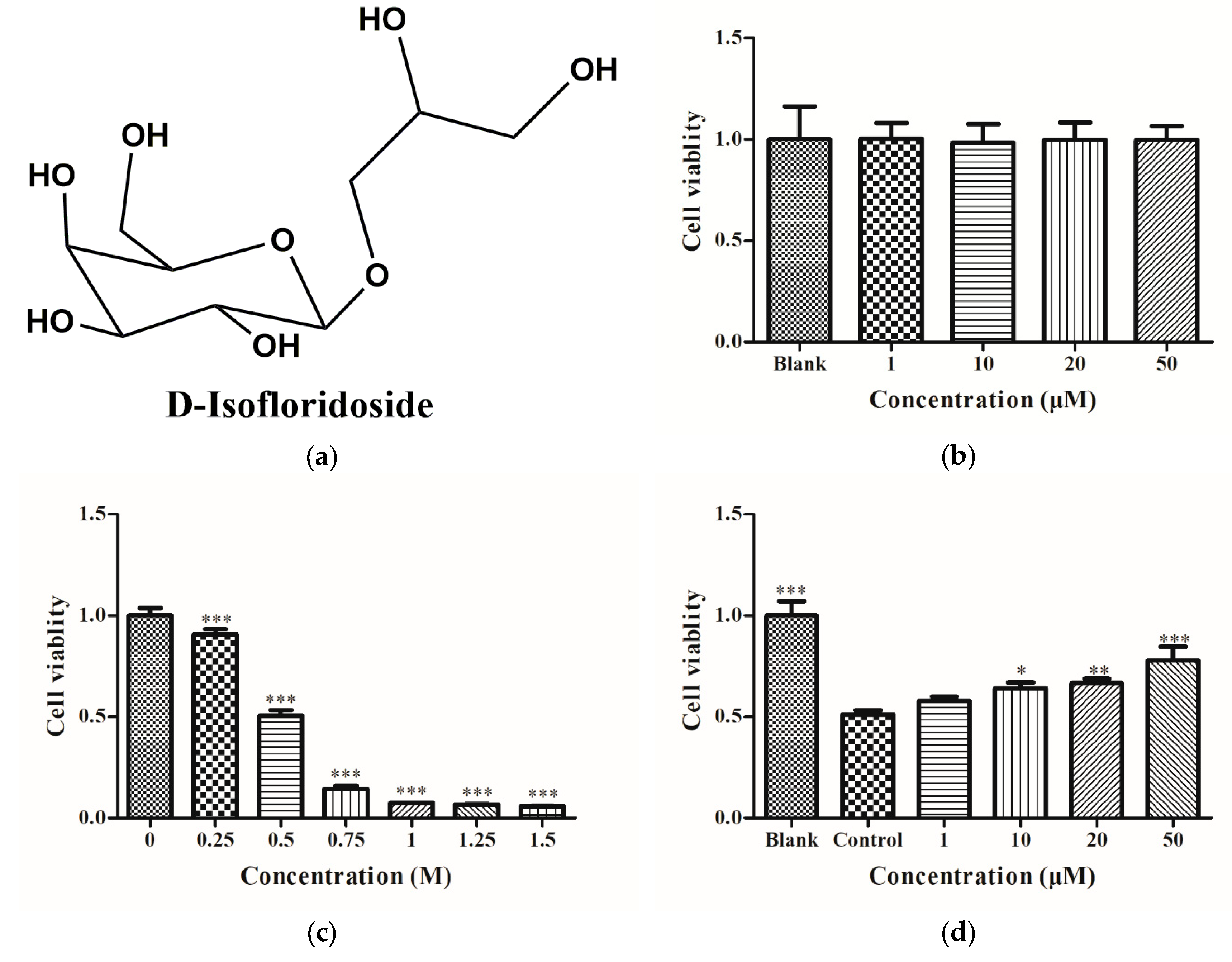
2.1. Cell Viability of HepG2 Cells
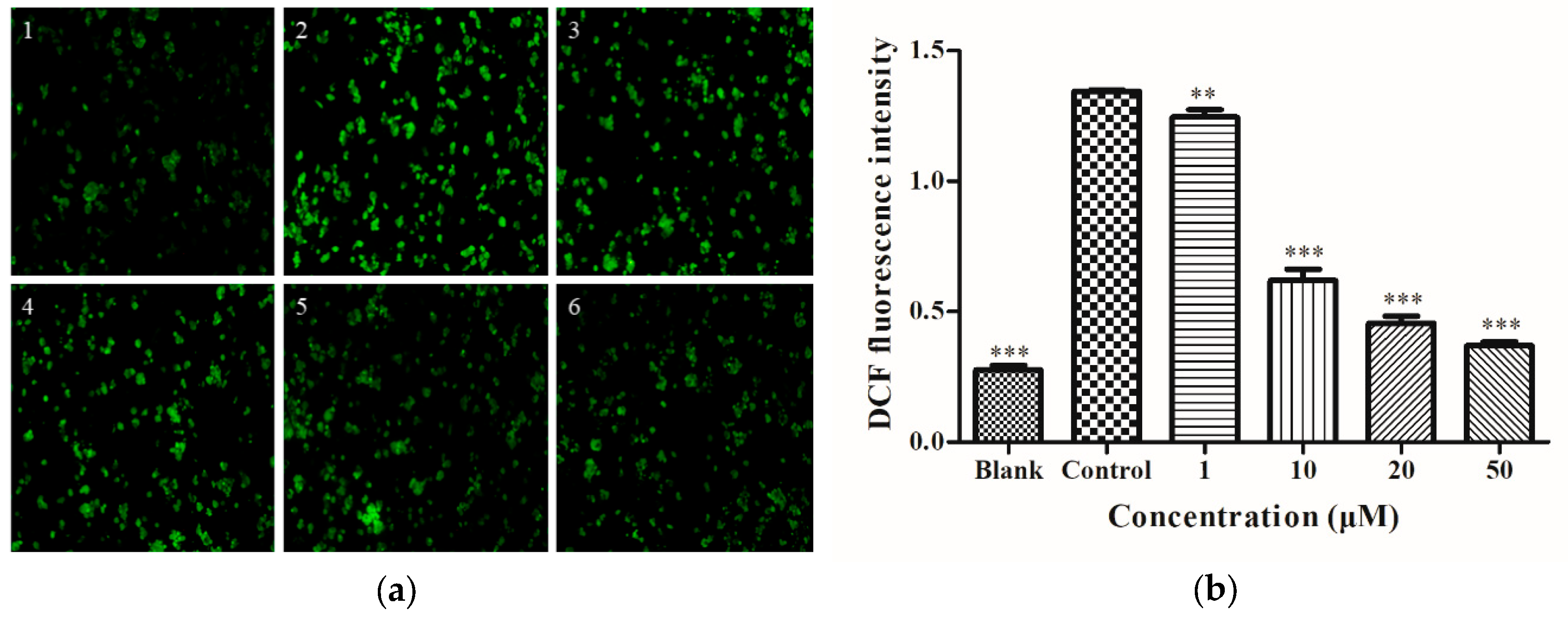
2.2. Determination of Intracellular ROS
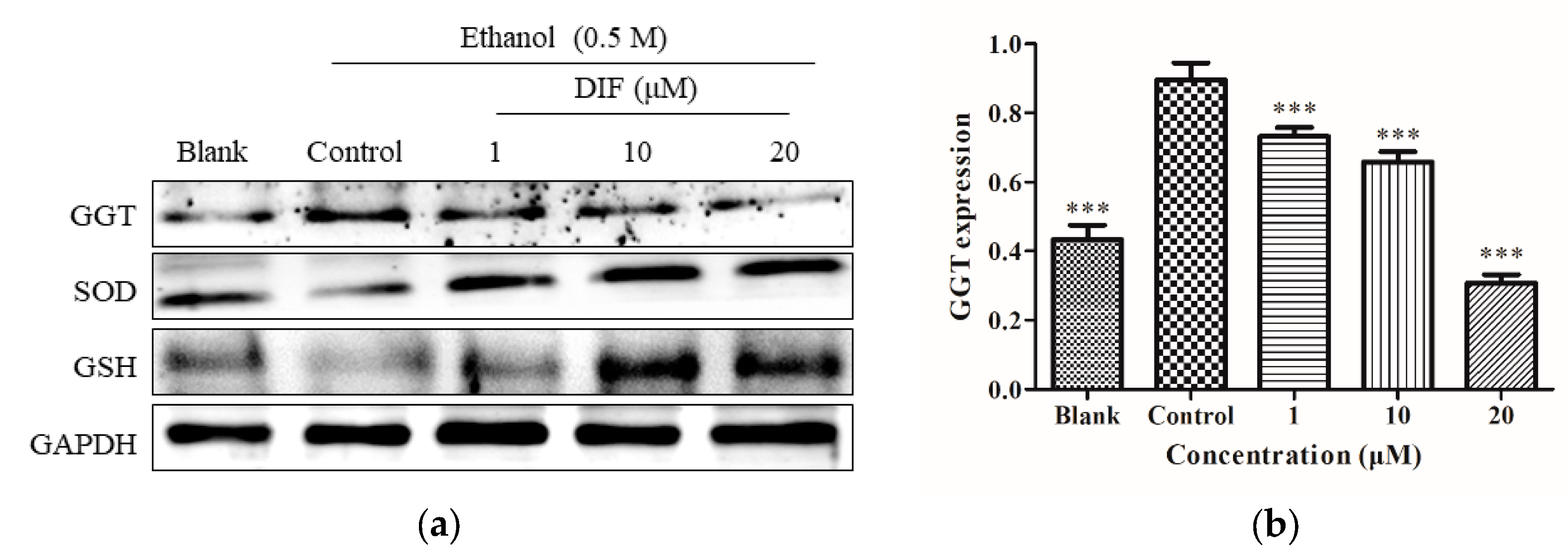
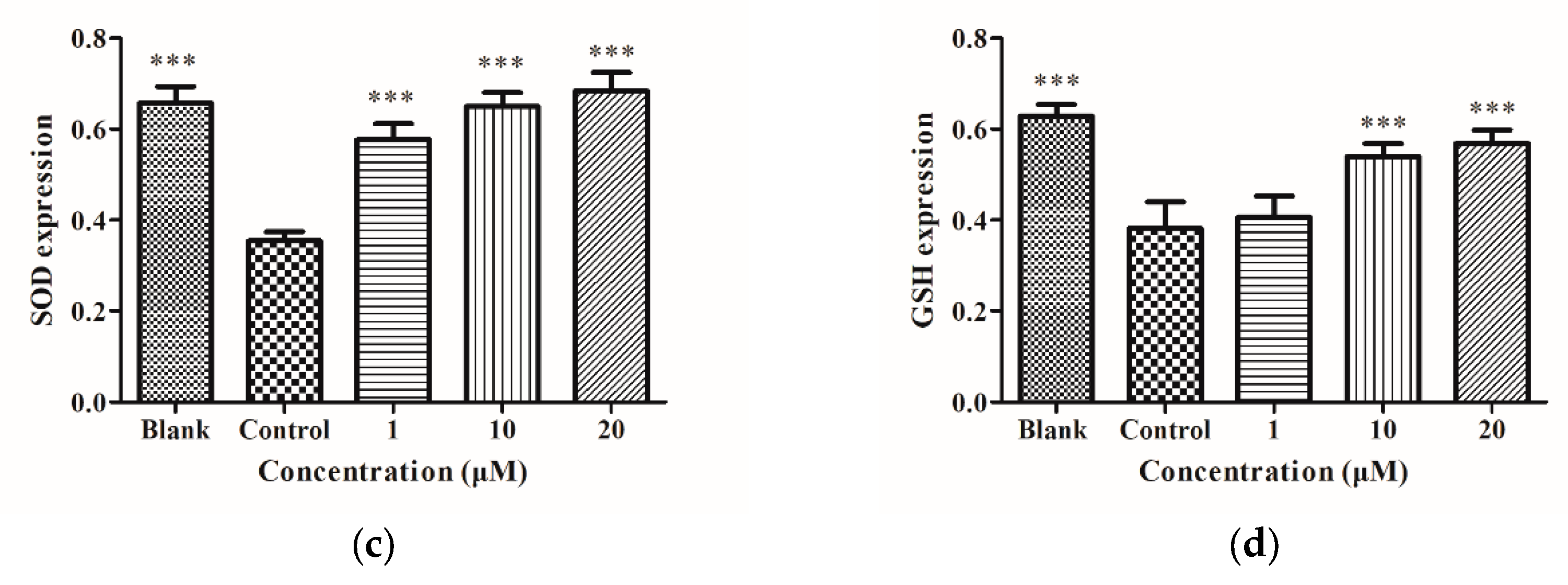
2.3. SOD, GSH, and GGT Protein Levels
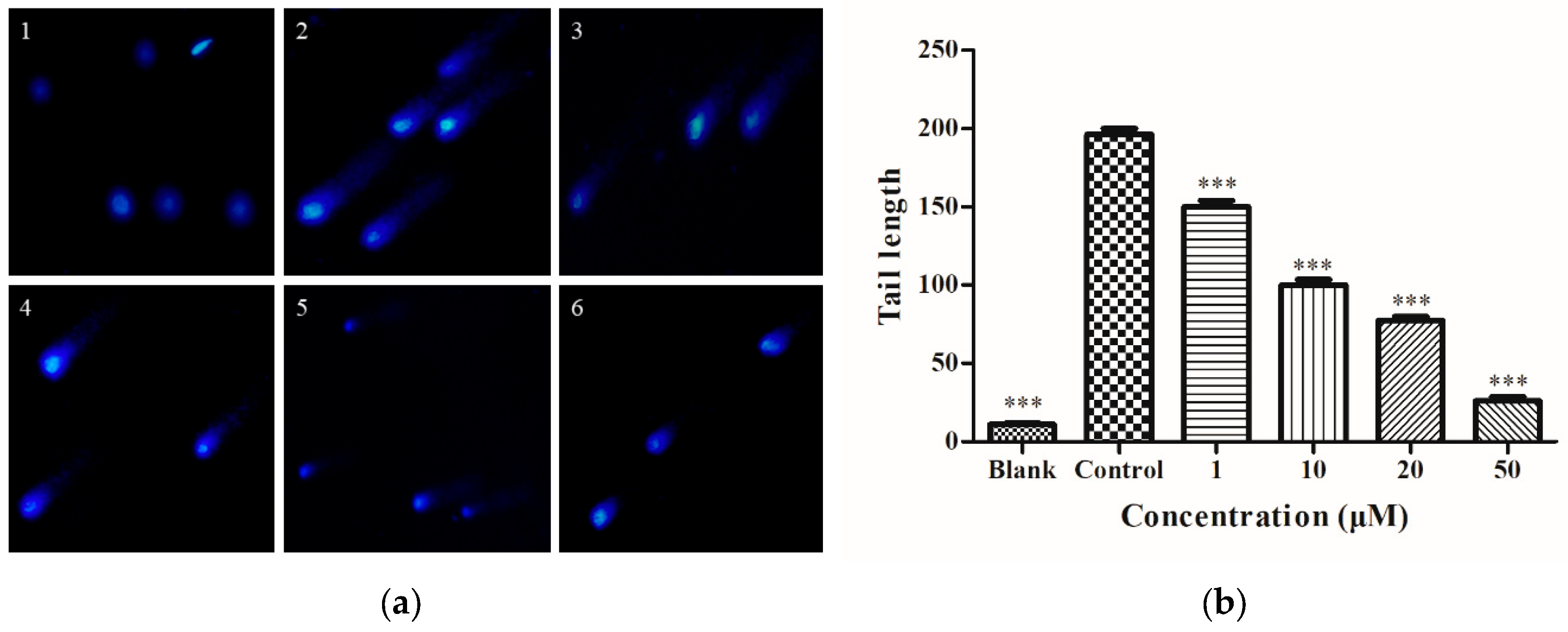
2.4. Determination of Intracellular DNA Damage
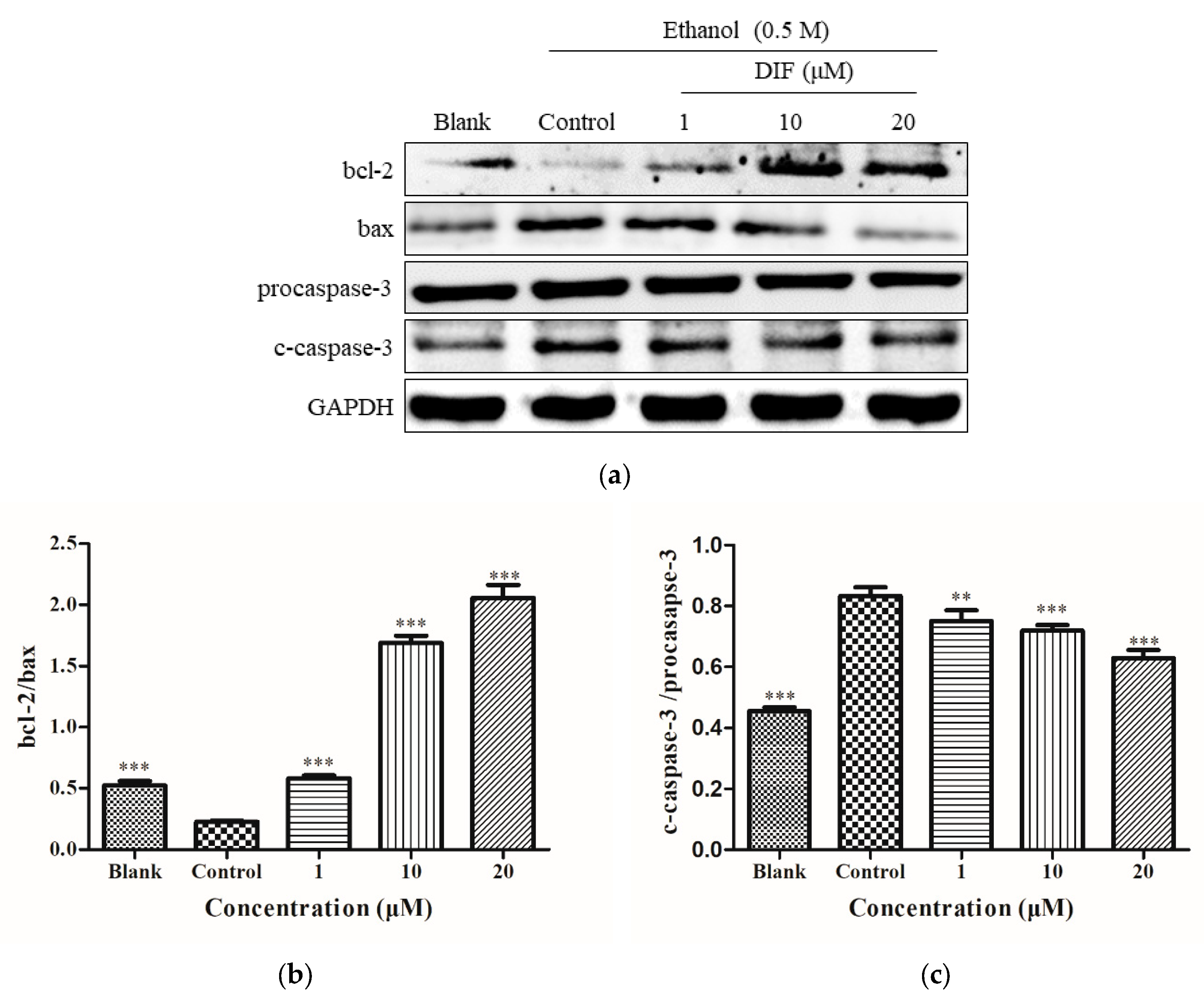
2.5. Caspase-3, bcl-2, and Bax Protein Levels
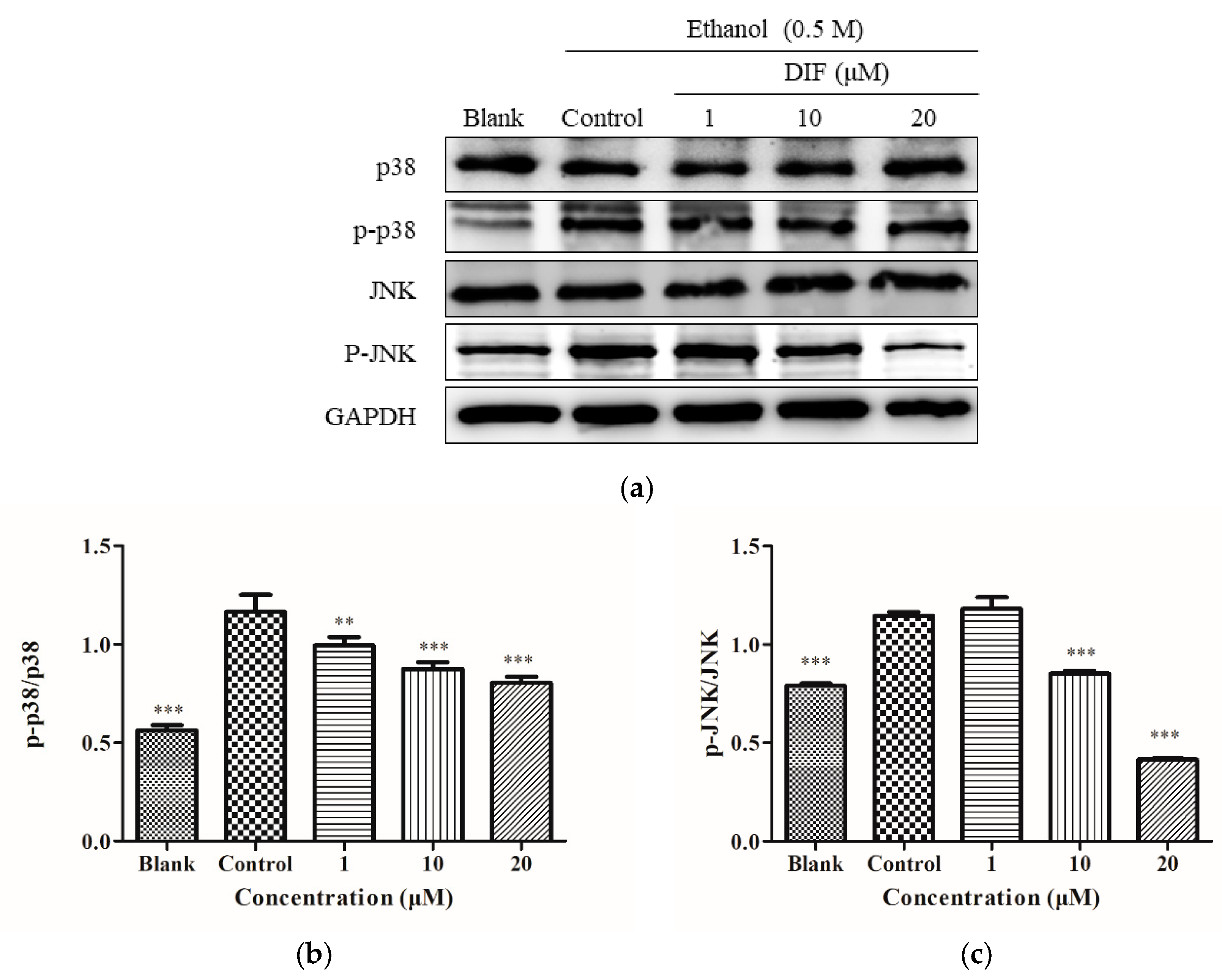
2.6. JNK and p38 Protein Levels
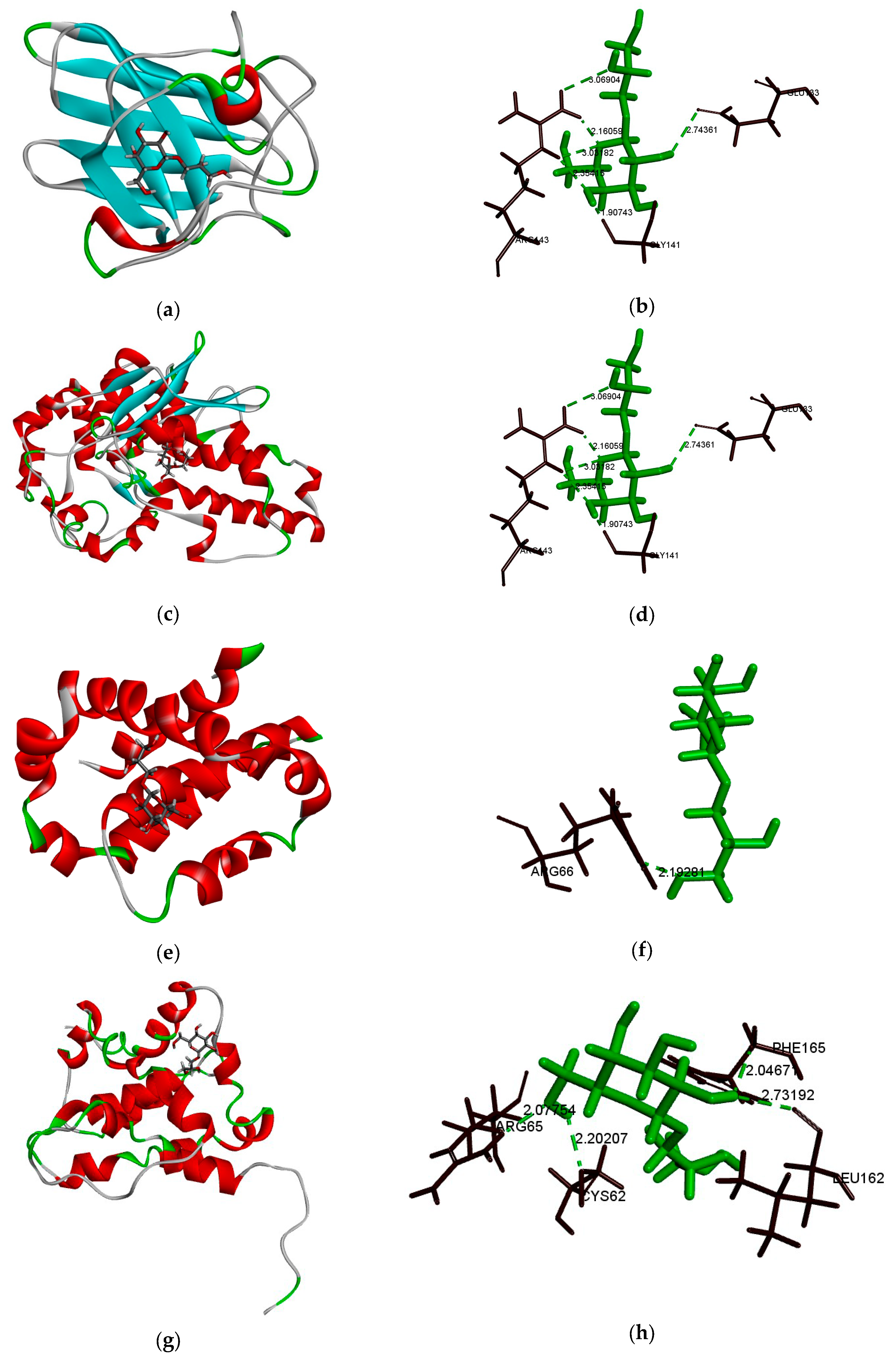
2.7. Bcl-2, Bax, SOD, and GGT Molecular Docking Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Cell ROS Analysis
4.5. Western Blot
4.6. Comet Assay
4.7. Molecular Docking
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mukherjee, S.; Das, S.K.; Vasudevan, D.M.; Cochin, E.P. Vasudevan Effects of Ethanol Consumption on Different Organs-A Brief Overview. Asian J. Biochem. 2007, 2, 386–394. [Google Scholar]
- Lieber, C.S. Alcohol and the liver: 1994 update. Gastroenterology 1994, 106, 1085–1105. [Google Scholar] [CrossRef]
- Arteel, G.E.; Crabb, D.W. Pathogenesis of Alcoholic Liver Disease. Clin. Liver Dis. 2016, 20, 445–456. [Google Scholar]
- Orman, E.S.; Odena, G.; Bataller, R. Alcoholic liver disease: Pathogenesis, management, and novel targets for therapy. J. Gastroenterol. Hepatol. 2013, 28, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Galicia-Moreno, M.; Gutiérrez-Reyes, G. The role of oxidative stress in the development of alcoholic liver disease. Mexican J. Gastroenterol. 2014, 79, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Ron, M. Oxidative stress, antioxidants and stress tolerance. Trends Plant. Sci. 2002, 7, 405–410. [Google Scholar]
- Hoek, J.B.; Cahill, A.; Pastorino, J.G. Alcohol and mitochondria: A dysfunctional relationship. Gastroenterology 2002, 122, 2049–2063. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.C.; Qiang, X.Y.; Zhang, M.; Ma, D.S.; Zhao, Z.; Zhou, C.S.; Liu, X.; Li, R.Y.; Chen, H.; Zhang, Y.B. Demethyleneberberine, a Natural Mitochondria-Targeted Antioxidant, Inhibits Mitochondrial Dysfunction, Oxidative Stress, and Steatosis in Alcoholic Liver Disease Mouse Model. J. Pharmacol. Exp. Ther. 2015, 352, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, L.; Wang, R.; Luo, X.; Li, Y.; Chen, Z. Protective effects of rice dreg protein hydrolysates against hydrogen peroxide-induced oxidative stress in HepG-2 cells. Food Funct 2016, 7, 1429–1437. [Google Scholar] [CrossRef]
- Chen, M.F.; Gong, F.; Zhang, Y.Y.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.J. Preventive Effect of YGDEY from Tilapia Fish Skin Gelatin Hydrolysates against Alcohol-Induced Damage in HepG2 Cells through ROS-Mediated Signaling Pathways. Nutrients 2019, 11, 392. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-F.; Zhang, Y.Y.; Di He, M.; Li, C.Y.; Zhou, C.X.; Hong, P.Z.; Qian, Z.-J. Antioxidant Peptide Purified from Enzymatic Hydrolysates of Isochrysis Zhanjiangensis and Its Protective Effect against Ethanol Induced Oxidative Stress of HepG2 Cells. Biotechnol. Bioprocess. Eng. 2019, 24, 308–317. [Google Scholar] [CrossRef]
- Praetorius Bjork, M.; Johansson, B. Gamma-Glutamyltransferase (GGT) as a biomarker of cognitive decline at the end of life: Contrasting age and time to death trajectories. Int. Psychogeriatr. 2018, 30, 981–990. [Google Scholar] [CrossRef]
- Kobayashi, T.; Masumoto, J.; Tada, T.; Nomiyama, T.; Hongo, K.; Nakayama, J. Prognostic significance of the immunohistochemical staining of cleaved caspase-3, an activated form of caspase-3, ion gliomas. Clin. Cancer Res. 2007, 13, 3868–3874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaysinski, M.; Ortalo-Magne, A.; Thomas, O.P.; Culioli, G. Extraction, Purification, and NMR Analysis of Terpenes from Brown Algae. Methods Mol. Biol. 2015, 1308, 207–223. [Google Scholar] [PubMed]
- Lucena, A.M.M.; Souza, C.R.M.; Jales, J.T.; Guedes, P.M.M.; de Miranda, G.E.C.; de Moura, A.M.A.; Araujo-Junior, J.X.; Nascimento, G.J.; Scortecci, K.C.; Santos, B.V.O.; et al. The Bisindole Alkaloid Caulerpin, from Seaweeds of the Genus Caulerpa, Attenuated Colon Damage in Murine Colitis Model. Mar. Drugs 2018, 16, 318. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, F.B.; Alves, R.C.; Harnedy, P.A.; FitzGerald, R.J.; Oliveira, M.B.P.P. Macroalgal-derived protein hydrolysates and bioactive peptides: Enzymatic release and potential health enhancing properties. Trends Food Sci. Technol. 2019, 93, 106–124. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, X.; Tang, Y.; Mao, J. Composition, isolation, purification and biological activities of Sargassum fusiforme polysaccharides: A review. Carbohydr. Polym. 2020, 228, 115381. [Google Scholar] [CrossRef]
- Santos, S.A.O.; Felix, R.; Pais, A.C.S.; Rocha, S.M.; Silvestre, A.J.D. The Quest for Phenolic Compounds from Macroalgae: A Review of Extraction and Identification Methodologies. Biomolecules 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Shilling, A.J.; von Salm, J.L.; Sanchez, A.R.; Kee, Y.; Amsler, C.D.; McClintock, J.B.; Baker, B.J.; Anverenes, B.-E. New Polyhalogenated Monoterpenes from the Antarctic Red Alga Plocamium cartilagineum. Mar. Drugs 2019, 17, 230. [Google Scholar] [CrossRef] [Green Version]
- Lebbar, S.; Fanuel, M.; Le Gall, S.; Falourd, X.; Ropartz, D.; Bressollier, P.; Gloaguen, V.; Faugeron-Girard, C. Agar Extraction By-Products from Gelidium sesquipedale as a Source of Glycerol-Galactosides. Molecules 2018, 23, 3364–3372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.Y.; Shabtai, Y.; Arad, S. Floridoside as a carbon precursor for the synthesis of cell-wall polysaccharide in the red microalga Porphyridium sp (Rhodophyta). J. Phycol. 2002, 38, 931–938. [Google Scholar] [CrossRef]
- Arad, S.M.; Friedman, O.D.; Rotem, A. Effect of Nitrogen on Polysaccharide Production in a Porphyridium sp. Appl. Environ. Microbiol. 1988, 54, 2411–2414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeuse, B.J.D.; Smith, B.N. A note on the amylolytic breakdown of some raw algal starches. Planta 1962, 57, 624–635. [Google Scholar] [CrossRef]
- Li, S.Y.; Lellouche, J.P.; Shabtai, Y.; Arad, S. Fixed carbon partitioning in the red microalga Porphyridium sp. (Rhodophyceae, Rhodophyta). J. Phycol. 2001, 37, 289–297. [Google Scholar] [CrossRef]
- Li, Y.X.; Li, Y.; Lee, S.H.; Qian, Z.J.; Kim, S.K. Inhibitors of oxidation and matrix metalloproteinases, floridoside, and D-isofloridoside from marine red alga Laurencia undulata. J. Agric. Food Chem. 2010, 58, 578–586. [Google Scholar] [CrossRef]
- Jin, M.; Liu, H.; Hou, Y.; Chan, Z.; Di, W.; Li, L.; Zeng, R. Preparation, characterization and alcoholic liver injury protective effects of algal oligosaccharides from Gracilaria lemaneiformis. Food Res. Int. 2017, 100, 186–195. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Mohibbullah, M.; An, J.H.; Choi, J.Y.; Hong, Y.K.; Sohn, J.H.; Kim, J.S.; Choi, J.S. In vivo antioxidant activity of mackerel (Scomber japonicus) muscle protein hydrolysate. PeerJ 2018, 6, e6181. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Tian, X.; Zhang, W.; Zheng, P.; Huang, F.; Ding, G.; Yang, Z. Protective Effects of Fucoxanthin against Alcoholic Liver Injury by Activation of Nrf2-Mediated Antioxidant Defense and Inhibition of TLR4-Mediated Inflammation. Mar. Drugs 2019, 17, 552. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Li, Y.X.; Dewapriya, P.; Ryu, B.M.; Kim, S.K. Floridoside suppresses pro-inflammatory responses by blocking MAPK signaling in activated microglia. BMB Rep. 2013, 46, 398–403. [Google Scholar] [CrossRef]
- Ryu, B.; Li, Y.-X.; Kang, K.-H.; Kim, S.-K.; Kim, D.G. Floridoside from Laurencia undulata promotes osteogenic differentiation in murine bone marrow mesenchymal cells. J. Funct. Food. 2015, 19, 505–511. [Google Scholar] [CrossRef]
- Barbosa, M.; Valentão, P.; Andrade, P.B. Bioactive Compounds from Macroalgae in the New Millennium: Implications for Neurodegenerative Diseases. Mar. Drugs 2014, 12, 4934–4972. [Google Scholar] [CrossRef] [PubMed]
- Putman, E.W.; Hassid, W.Z. Structure of Galactosylglycerol from Irideae laminarioides. J. Am. Chem. Soc. 1954, 76, 2221–2223. [Google Scholar] [CrossRef]
- KAUSS, H. Metabolism of Isofloridoside (O-α-D-Galactopyranosyl-(1→1)-Glycerol) and Osmotic Balance in the Fresh Water Alga Ochromonas. Nature 1967, 214, 1129–1130. [Google Scholar] [CrossRef]
- Pade, N.; Linka, N.; Ruth, W.; Weber, A.P.M.; Hagemann, M. Floridoside and isofloridoside are synthesized by trehalose 6-phosphate synthase-like enzymes in the red alga Galdieria sulphuraria. New Phytol. 2015, 205, 1227–1238. [Google Scholar] [CrossRef]
- Majak, W.; Craigie, J.S.; Mclachlan, J. Photosynthesis in algae. I. Accumulation products in the Rhodophyceae. Can. J. Bot. 1966, 44, 541–549. [Google Scholar] [CrossRef]
- Macler, B.A. Regulation of Carbon Flow by Nitrogen and Light in the Red Alga, Gelidium coulteri. Plant. Physiol. 1986, 82, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Ekman, P.; Yu, S.; Pedersen, M. Effects of altered salinity, darkness and algal nutrient status on floridoside and starch content, α-galactosidase activity and agar yield of cultivatedGracilaria sordida. Eur. J. Phycol. 1991, 26, 123–131. [Google Scholar] [CrossRef]
- Kroen, W.K.; Ramus, J. Allocation of Newly-Fixed Carbon in the Red Alga Rhodella. J. Plant. Physiol. 1991, 138, 522–527. [Google Scholar] [CrossRef]
- You, Y.; Min, S.; Lee, Y.H.; Hwang, K.; Jun, W. Hepatoprotective effect of 10% ethanolic extract from Curdrania tricuspidata leaves against ethanol-induced oxidative stress through suppression of CYP2E1. Food Chem. Toxicol. 2017, 108, 298–304. [Google Scholar] [CrossRef]
- Lee, J.; Yang, J.; Jeon, J.; Jeong, H.S.; Lee, J.; Sung, J. Hepatoprotective effect of esculetin on ethanol-induced liver injury in human HepG2 cells and C57BL/6J mice. J. Funct. Food. 2018, 40, 536–543. [Google Scholar] [CrossRef]
- Williams, G.T. Programmed cell death: Apoptosis and oncogenesis. Cell 1991, 65, 1097–1098. [Google Scholar] [CrossRef]
- Zou, H.; Henzel, W.J.; Liu, X.; Lutschg, A.; Wang, X. Apaf-1, a human protein homologous to C. elegans CED-4, participates in cytochrome c-dependent activation of caspase-3. Cell 1997, 90, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Ahmad Hidayat, A.F.; Chan, C.K.; Mohamad, J.; Abdul Kadir, H. Dioscorea bulbifera induced apoptosis through inhibition of ERK 1/2 and activation of JNK signaling pathways in HCT116 human colorectal carcinoma cells. Biomed. Pharmacother. 2018, 104, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Luo, H.; Xia, H. Theaflavins attenuate ethanolinduced oxidative stress and cell apoptosis in gastric mucosa epithelial cells via downregulation of the mitogenactivated protein kinase pathway. Mol. Med. Rep. 2018, 18, 3791–3799. [Google Scholar] [PubMed] [Green Version]
- Becatti, M.; Barygina, V.; Mannucci, A.; Emmi, G.; Prisco, D.; Lotti, T.; Fiorillo, C.; Taddei, N. Sirt1 Protects against Oxidative Stress-Induced Apoptosis in Fibroblasts from Psoriatic Patients: A New Insight into the Pathogenetic Mechanisms of Psoriasis. Int. J. Mol. Sci. 2018, 19, 1572–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Miao, L.S.; Cai, Y.M.; He, J.X.; Zhang, Z.N.; Wu, G.; Zheng, J. TXNIP knockdown alleviates hepatocyte ischemia reperfusion injury through preventing p38/JNK pathway activation. Biochem. Biophys. Res. Commun. 2018, 502, 409–414. [Google Scholar] [CrossRef]
- Feng, W.; Li, J.; Liao, S.; Ma, S.; Li, F.; Zhong, C.; Li, G.; Wei, Y.; Huang, H.; Wei, Q.; et al. Go6983 attenuates titanium particle-induced osteolysis and RANKL mediated osteoclastogenesis through the suppression of NFkappaB/JNK/p38 pathways. Biochem. Biophys. Res. Commun. 2018, 503, 62–70. [Google Scholar] [CrossRef]
- Shen, H.M.; Liu, Z.G. JNK signaling pathway is a key modulator in cell death mediated by reactive oxygen and nitrogen species. Free Radic. Biol. Med. 2006, 40, 928–939. [Google Scholar] [CrossRef]
- Nakagawa, H.; Maeda, S. Molecular mechanisms of liver injury and hepatocarcinogenesis: Focusing on the role of stress-activated MAPK. Pathol. Res. Int. 2012, 2012, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.F.; Zhong, Y.J.; Ma, Z.H.; Li, L.; Shi, L.; Chen, L.; Li, C.; Wu, D.; Chen, Q.; Li, Y.W. NOX4/ROS mediate ethanol-induced apoptosis via MAPK signal pathway in L-02 cells. Int. J. Mol. Med. 2018, 41, 2306–2316. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wei, X.; Wu, Y.; Wang, Y.; Qiu, Y.; Shi, J.; Zhou, H.; Lu, Z.; Shao, M.; Yu, L.; et al. Giganteaside D induces ROS-mediated apoptosis in human hepatocellular carcinoma cells through the MAPK pathway. Cell. Oncol. 2016, 39, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Shen, X.; Kong, J. Determining the purity of samples from natural products by coupling HPLC and CCD spectrometry. J. Sep. Sci. 2005, 28, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Fontaine, F.; Cooper, M.A. Natural product libraries: Assembly, maintenance, and screening. Planta Med. 2014, 80, 1161–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondu, S.; Kervarec, N.; Deslandes, E.; Pichon, R. Separation of floridoside and isofloridosides by HPLC and complete H-1 and C-13 NMR spectral assignments for D-isofloridoside. Carbohydr. Res. 2007, 342, 2470–2473. [Google Scholar] [CrossRef]
- Osseni, R.A.; Debbasch, C.; Christen, M.O.; Rat, P.; Warnet, J.M. Tacrine-induced Reactive Oxygen Species in a Human Liver Cell Line: The Role of Anethole Dithiolethione as a Scavenger. Toxicol. Vitro 1999, 13, 683–688. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, Y.; Yang, C. Evaluating In Vitro DNA Damage Using Comet Assay. J. Vis. Exp. 2017, 128, e56450. [Google Scholar] [CrossRef]
- Hansson, A.; Souza, P.C.T.; Silveira, R.L.; Martínez, L.; Skaf, M.S. CHARMM force field parameterization of rosiglitazone. Int. J. Quantum Chem. 2011, 111, 1346–1354. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds (D-Isofloridoside, from Laurencia undulate) are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Ligand | Receptor | -CDOCKER Interaction Energy (kcal/mol) |
|---|---|---|---|
| 1 | DIF | SOD | 29.0355 |
| 2 | 28.4436 | ||
| 3 | 28.3838 | ||
| 4 | 27.5988 | ||
| 5 | 26.8758 | ||
| 6 | 26.1050 | ||
| 1 | GGT | 34.7056 | |
| 2 | 32.5408 | ||
| 3 | 31.8090 | ||
| 4 | 29.3496 | ||
| 5 | 26.5751 | ||
| 6 | 26.0123 | ||
| 7 | 24.6752 |
| Number | Ligand | Receptor | -CDOCKER Interaction Energy (kcal/mol) |
|---|---|---|---|
| 1 | DIF | bcl-2 | 29.5175 |
| 2 | 29.0723 | ||
| 3 | 28.5170 | ||
| 4 | 28.3632 | ||
| 5 | 28.1850 | ||
| 6 | 27.7746 | ||
| 7 | 27.6238 | ||
| 1 | bax | 25.6085 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Chen, M.-F.; Ryu, B.; Chen, J.; Xiao, Z.; Hong, P.; Sun, S.; Wang, D.; Qian, Z.-J.; Zhou, C. The Protective Effect of the Polysaccharide Precursor, D-Isofloridoside, from Laurencia undulata on Alcohol-Induced Hepatotoxicity in HepG2 Cells. Molecules 2020, 25, 1024. https://doi.org/10.3390/molecules25051024
Yang S, Chen M-F, Ryu B, Chen J, Xiao Z, Hong P, Sun S, Wang D, Qian Z-J, Zhou C. The Protective Effect of the Polysaccharide Precursor, D-Isofloridoside, from Laurencia undulata on Alcohol-Induced Hepatotoxicity in HepG2 Cells. Molecules. 2020; 25(5):1024. https://doi.org/10.3390/molecules25051024
Chicago/Turabian StyleYang, Shengtao, Mei-Fang Chen, Bomi Ryu, Jiali Chen, Zhenbang Xiao, Pengzhi Hong, Shengli Sun, Di Wang, Zhong-Ji Qian, and Chunxia Zhou. 2020. "The Protective Effect of the Polysaccharide Precursor, D-Isofloridoside, from Laurencia undulata on Alcohol-Induced Hepatotoxicity in HepG2 Cells" Molecules 25, no. 5: 1024. https://doi.org/10.3390/molecules25051024
APA StyleYang, S., Chen, M.-F., Ryu, B., Chen, J., Xiao, Z., Hong, P., Sun, S., Wang, D., Qian, Z.-J., & Zhou, C. (2020). The Protective Effect of the Polysaccharide Precursor, D-Isofloridoside, from Laurencia undulata on Alcohol-Induced Hepatotoxicity in HepG2 Cells. Molecules, 25(5), 1024. https://doi.org/10.3390/molecules25051024