
Ginseng Gintonin Contains Ligands for GPR40 and GPR55

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Gintonin-Induced Insulin Secretion in INS-1 Cells and Rat Islets
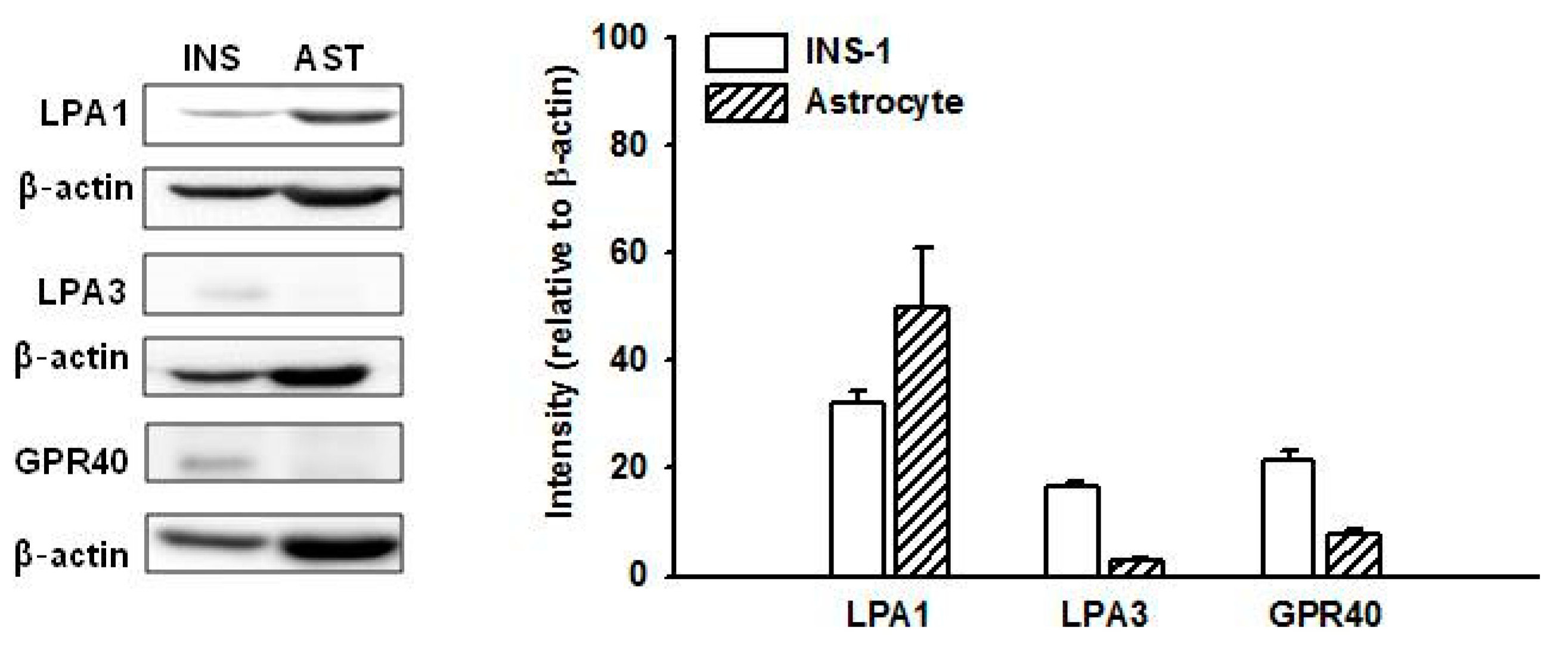
2.2. Expression of GPR40 and LPA Receptors in INS-1 Cells
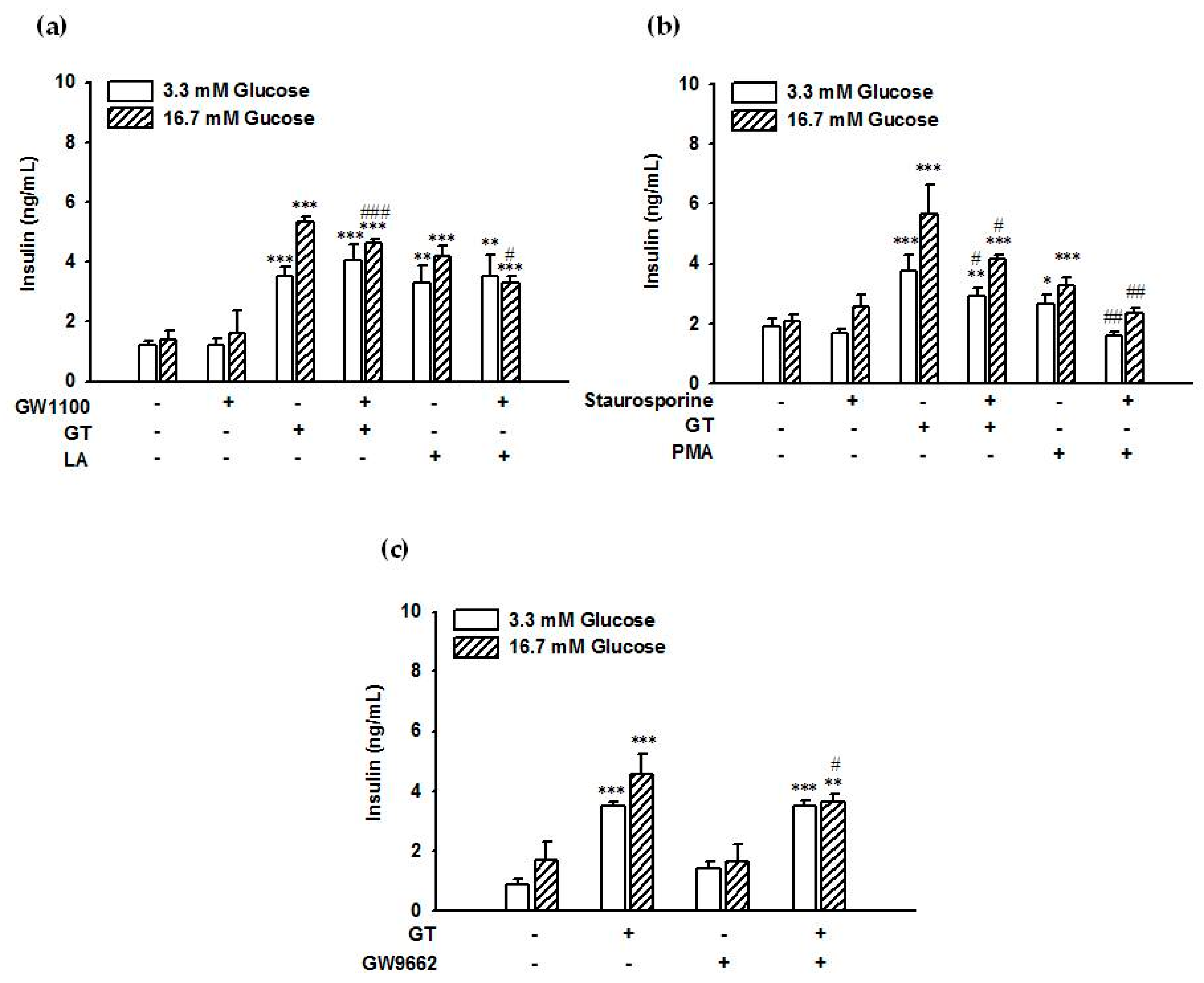
2.3. Suppression of GPR40 Expression Reduced Gintonin-Induced Insulin Secretion from INS-1 Cells
2.4. GPR40 and Protein Kinase C (PKC) Inhibitors Attenuated Gintonin-Induced Insulin Secretion from INS-1 Cells
2.5. The PPARγ Inhibitor Partially Attenuated Gintonin-Induced Insulin Secretion from INS-1 Cells
2.6. Effect of Gintonin on Intracellular Calcium Mobilization in PC-3 Cells
2.7. Effect of Gintonin on PC-3 Cell Migration
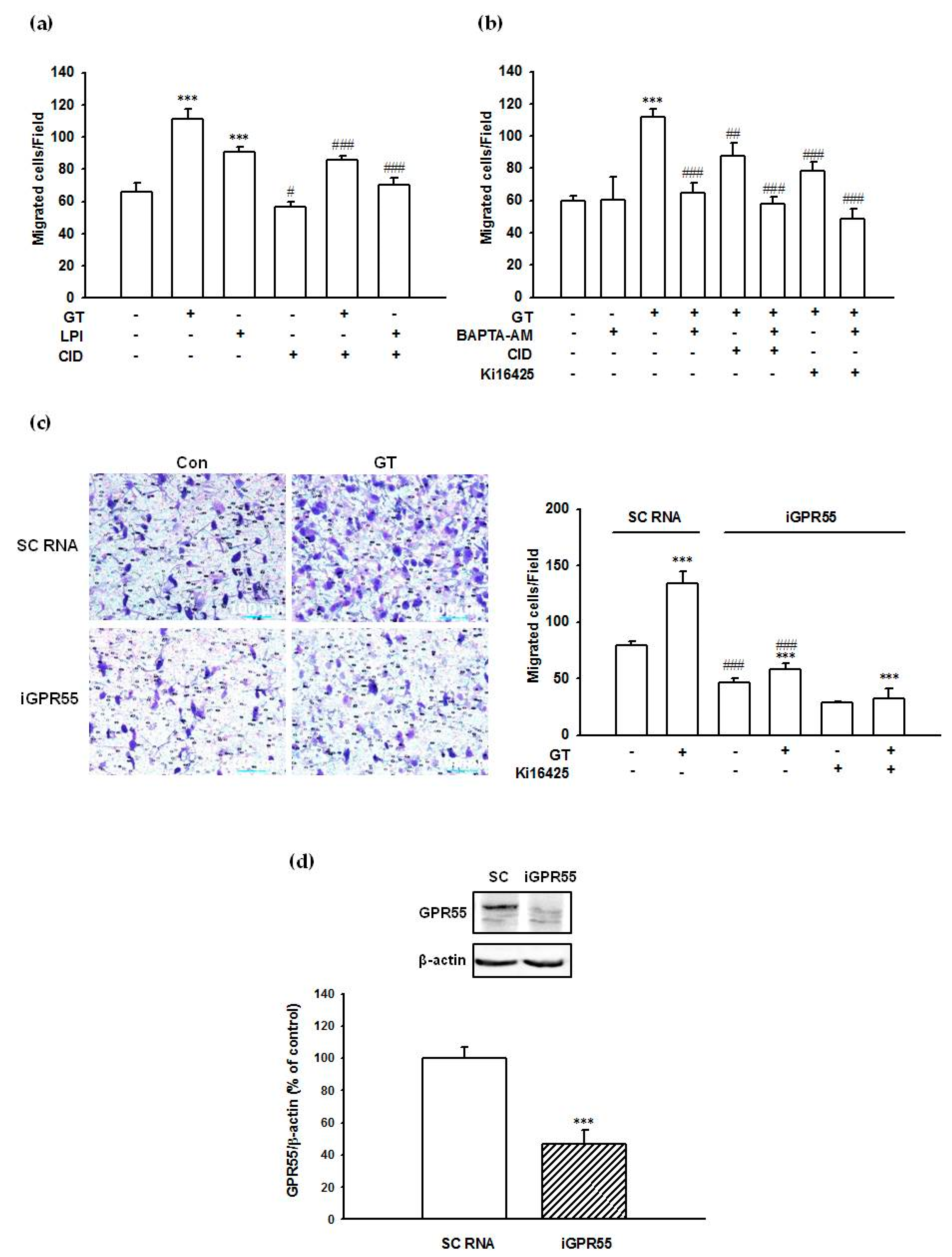
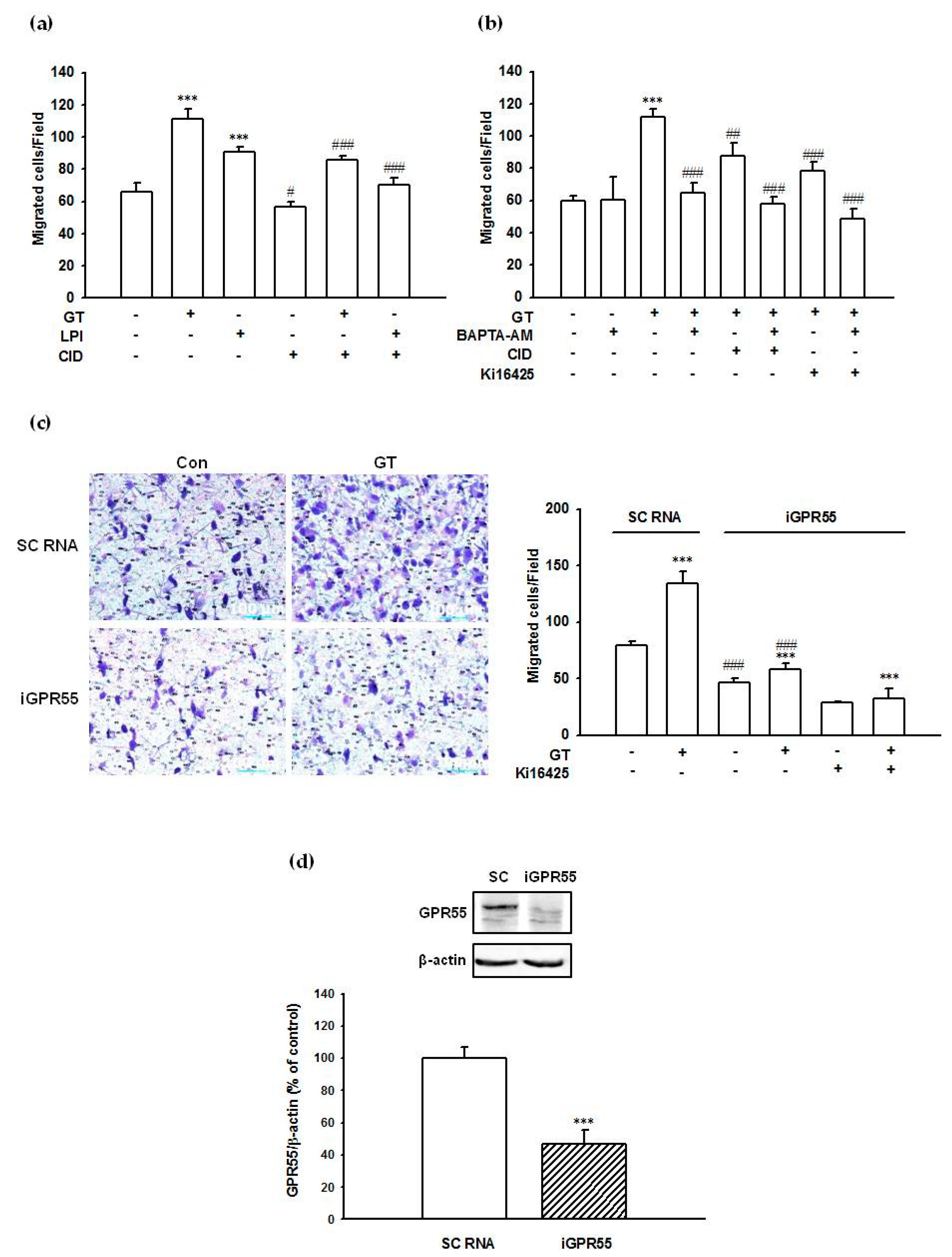
2.8. Gintonin-Mediated PC-3 Cell Migration Was Partially Inhibited by a GPR55 Inhibitor, an LPA1/3 Antagonist, and a Calcium Chelator
2.9. Suppression of GPR55 Expression by siRNA Reduced Gintonin-Mediated PC-3 Cell Migration
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Viability
4.4. Insulin Secretion from INS-1 Cells
4.5. Islet Isolation and Insulin Secretion Assay
4.6. Small-Interfering RNA Transfection
4.7. Western Blot Analysis
4.8. Migration Assay Using Modified Boyden Chambers
4.9. Measurement of Intracellular Calcium Concentrations
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Choi, K.T. Botanical characteristics, pharmacological effects and medicinal components of Korean Panax ginseng C A Meyer. Acta Pharm. Sin. 2008, 29, 1109–1118. [Google Scholar] [CrossRef] [Green Version]
- Wee, J.J.; Park, K.M.; Chung, A.S. Chapter 8: Biological activities of ginseng and its application to human health. In Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; Benzie, I.F.F., Wachtel-Galor, S., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2011. [Google Scholar]
- Pyo, M.K.; Choi, S.H.; Shin, T.J.; Hwang, S.H.; Lee, B.H.; Kang, J.; Kim, H.J.; Lee, S.H.; Nah, S.Y. A simple method for the preparation of crude gintonin from ginseng root, stem, and leaf. J. Ginseng Res. 2011, 35, 209–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.H.; Shin, T.J.; Choi, S.H.; Cho, H.J.; Lee, B.H.; Pyo, M.K.; Lee, J.H.; Kang, J.K.; Kim, H.J.; Park, C.W.; et al. Gintonin, newly identified compounds from ginseng, is novel lysophosphatidic acids-protein complexes and activates G protein-coupled lysophosphatidic acid receptors with high affinity. Mol. Cells 2012, 33, 151–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.H.; Shin, E.J.; Shin, T.J.; Lee, B.H.; Choi, S.H.; Kang, J.; Kim, H.J.; Kwon, S.H.; Jang, C.G.; Lee, J.H.; et al. Gintonin, a ginseng-derived lysophosphatidic acid receptor ligand, attenuates Alzheimer’s disease-related neuropathies: Involvement of non-amyloidogenic processing. J. Alzheimers Dis. 2012, 31, 207–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.H.; Lee, B.H.; Choi, S.H.; Kim, H.J.; Jung, S.W.; Kim, H.S.; Shin, H.C.; Park, H.J.; Park, K.H.; Lee, M.K.; et al. Gintonin, a novel ginseng-derived lysophosphatidic acid receptor ligand, stimulates neurotransmitter release. Neurosci. Lett. 2015, 584, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Kim, H.J.; Cho, H.J.; Park, S.D.; Lee, N.E.; Hwang, S.H.; Cho, I.H.; Hwang, H.; Rhim, H.; Kim, H.C.; et al. Gintonin, a ginseng-derived exogenous lysophosphatidic acid receptor ligand, protects astrocytes from hypoxic and re-oxygenation stresses through stimulation of astrocytic glycogenolysis. Mol. Neurobiol. 2019, 56, 3280–3294. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.H.; Lee, B.H.; Choi, S.H.; Kim, H.J.; Won, K.J.; Lee, H.M.; Rhim, H.; Kim, H.C.; Nah, S.Y. Effects of gintonin on the proliferation, migration, and tube formation of human umbilical-vein endothelial cells: Involvement of lysophosphatidic-acid receptors and vascular-endothelial-growth-factor signaling. J. Ginseng Res. 2016, 40, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Jung, S.W.; Lee, B.H.; Kim, H.J.; Hwang, S.H.; Kim, H.K.; Nah, S.Y. Ginseng pharmacology: A new paradigm based on gintonin-lysophosphatidic acid receptor interactions. Front. Pharm. 2015, 6, 245. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Kim, D.J.; Shin, E.J.; Lee, B.H.; Choi, S.H.; Hwang, S.H.; Rhim, H.; Cho, I.H.; Kim, H.C.; Nah, S.Y. Effects of gintonin-enriched fraction on hippocampal cell proliferation in wild-type mice and an APPswe/PSEN-1 double Tg mouse model of Alzheimer’s disease. Neurochem. Int. 2016, 101, 56–65. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, S.D.; Lee, R.M.; Lee, B.H.; Choi, S.H.; Hwang, S.H.; Rhim, H.; Kim, H.C.; Nah, S.Y. Gintonin attenuates depressive-like behaviors associated with alcohol withdrawal in mice. J. Affect. Disord. 2017, 215, 23–29. [Google Scholar] [CrossRef]
- Cho, H.J.; Choi, S.H.; Kim, H.J.; Lee, B.H.; Rhim, H.; Kim, H.C.; Hwang, S.H.; Nah, S.Y. Bioactive lipids in gintonin-enriched fraction from ginseng. J. Ginseng Res. 2019, 43, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Tomita, T.; Hosoda, K.; Fujikura, J.; Inagaki, N.; Nakao, K. The G-Protein-coupled long-chain fatty acid receptor GPR40 and glucose metabolism. Front. Endocrinol. (Lausanne) 2014, 5, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.T.; Leng, J.; Xie, Z.; Li, S.L.; Zhao, W.; Tang, Q.L. GPR40: A therapeutic target for mediating insulin secretion (review). Int. J. Mol. Med. 2012, 30, 1261–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burant, C.F.; Viswanathan, P.; Marcinak, J.; Cao, C.; Vakilynejad, M.; Xie, B.; Leifke, E. TAK-875 versus placebo or glimepiride in type 2 diabetes mellitus: A phase 2, randomised, double-blind, placebo-controlled trial. Lancet 2012, 379, 1403–1411. [Google Scholar] [CrossRef]
- Yamada, H.; Yoshida, M.; Ito, K.; Dezaki, K.; Yada, T.; Ishikawa, S.E.; Kakei, M. Potentiation of glucose-stimulated insulin secretion by the GPR40-PLC-TRPC Pathway in pancreatic β-cells. Sci. Rep. 2016, 6, 25912. [Google Scholar] [CrossRef]
- Oka, S.; Nakajima, K.; Yamashita, A.; Kishimoto, S.; Sugiura, T. Identification of GPR55 as a lysophosphatidylinositol receptor. Biochem. Biophys. Res. Commun. 2007, 362, 928–934. [Google Scholar] [CrossRef]
- Lauckner, J.E.; Jensen, J.B.; Chen, H.Y.; Lu, H.C.; Hille, B.; Mackie, K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar] [CrossRef] [Green Version]
- Piñeiro, R.; Maffucci, T.; Falasca, M. The putative cannabinoid receptor GPR55 defines a novel autocrine loop in cancer cell proliferation. Oncogene 2011, 30, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Oka, S.; Kimura, S.; Toshida, T.; Ota, R.; Yamashita, A.; Sugiura, T. Lysophosphatidylinositol induces rapid phosphorylation of p38 mitogen-activated protein kinase and activating transcription factor 2 in HEK293 cells expressing GPR55 and IM-9 lymphoblastoid cells. J. Biochem. 2010, 147, 671–678. [Google Scholar] [CrossRef]
- Piñeiro, R.; Falasca, M. Lysophosphatidylinositol signalling: New wine from an old bottle. Biochim. Biophys. Acta 2012, 1821, 694–705. [Google Scholar] [CrossRef] [Green Version]
- Marichal-Cancino, B.A.; Fajardo-Valdez, A.; Ruiz-Contreras, A.E.; Mendez-Díaz, M.; Prospero-García, O. Advances in the physiology of GPR55 in the central nervous system. Curr. Neuropharmacol. 2017, 15, 771–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henstridge, C.M. Off-target cannabinoid effects mediated by GPR55. Pharmacology 2012, 89, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Tudurí, E.; Imbernon, M.; Hernández-Bautista, R.J.; Tojo, M.; Fernø, J.; Diéguez, C.; Nogueiras, R. GPR55: A new promising target for metabolism? J. Mol. Endocrinol. 2017, 58, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Spohr, T.C.; Choi, J.W.; Gardell, S.E.; Herr, D.R.; Rehen, S.K.; Gomes, F.C.; Chun, J. Lysophosphatidic acid receptor-dependent secondary effects via astrocytes promote neuronal differentiation. J. Biol. Chem. 2008, 283, 7470–7479. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, K.; Nishinaka, T.; Sato, N.; Mankura, M.; Koyama, Y.; Tokuyama, S. Hypothalamic GPR40 signaling activated by free long chain fatty acids suppresses CFA-induced inflammatory chronic pain. PLoS ONE 2013, 8, e81563. [Google Scholar] [CrossRef] [Green Version]
- Malenczyk, K.; Jazurek, M.; Keimpema, E.; Silvestri, C.; Janikiewicz, J.; Mackie, K.; Di Marzo, V.; Redowicz, M.J.; Harkany, T.; Dobrzyn, A. CB1 cannabinoid receptors couple to focal adhesion kinase to control insulin release. J. Biol. Chem. 2013, 288, 32685–32699. [Google Scholar] [CrossRef] [Green Version]
- Monet, M.; Gkika, D.; Lehen’kyi, V.; Pourtier, A.; Vanden Abeele, F.; Bidaux, G.; Juvin, V.; Rassendren, F.; Humez, S.; Prevarsakaya, N. Lysophospholipids stimulate prostate cancer cell migration via TRPV2 channel activation. Biochim. Biophys. Acta 2009, 1793, 528–539. [Google Scholar] [CrossRef] [Green Version]
- Hao, F.; Tan, M.; Xu, X.; Han, J.; Miller, D.D.; Tigyi, G.; Cui, M.Z. Lysophosphatidic acid induces prostate cancer PC3 cell migration via activation of LPA(1), p42 and p38alpha. Biochim. Biophys. Acta 2007, 1771, 883–892. [Google Scholar] [CrossRef] [Green Version]
- Briscoe, C.P.; Tadayyon, M.; Andrews, J.L.; Benson, W.G.; Chambers, J.K.; Eilert, M.M.; Ellis, C.; Elshourbagy, N.A.; Goetz, A.S.; Minnick, D.T.; et al. The orphan G protein-coupled receptor GPR40 is activated by medium and long chain fatty acids. J. Biol. Chem. 2003, 278, 11303–11311. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Kawamata, Y.; Harada, M.; Kobayashi, M.; Fujii, R.; Fukusumi, S.; Fukusumi, S.; Ogi, K.; Hosoya, M.; Tanaka, Y.; et al. Free fatty acids regulate insulin secretion from pancreatic beta cells through GPR40. Nature 2003, 422, 173–176. [Google Scholar] [CrossRef]
- Poitout, V.; Lin, D.C. Modulating GPR40: Therapeutic promise and potential in diabetes. Drug Discov. Today 2013, 18, 1301–1308. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, H.; Shachar, S.; Sekler, I.; Hershfinkel, M.; Walker, M.D. Role of GPR40 in fatty acid action on the beta cell line INS-1E. Biochem. Biophys. Res. Commun. 2005, 335, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Riddy, D.M.; Delerive, P.; Summers, R.J.; Sexton, P.M.; Langmead, C.J. G Protein-coupled receptors targeting insulin resistance, obesity, and type 2 diabetes mellitus. Pharm. Rev. 2018, 70, 39–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.H.; Kim, J.; Lee, R.M.; Choi, S.H.; Kim, H.J.; Hwang, S.H.; Lee, M.K.; Bae, C.S.; Kim, H.C.; Rhim, H.; et al. Gintonin enhances performance of mice in rotarod test: Involvement of lysophosphatidic acid receptors and catecholamine release. Neurosci. Lett. 2016, 612, 256–260. [Google Scholar] [CrossRef]
- Kim, S.; Kim, M.S.; Park, K.; Kim, H.J.; Jung, S.W.; Nah, S.Y.; Han, J.S.; Chung, C. Hippocampus-dependent cognitive enhancement induced by systemic gintonin administration. J. Ginseng Res. 2016, 40, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Asfari, M.; Janjic, D.; Meda, P.; Li, G.; Halban, P.A.; Wollheim, C.B. Establishment of 2-mercaptoethanol-dependent differentiated insulin-secreting cell lines. Endocrinology 1992, 130, 167–178. [Google Scholar] [CrossRef]
- Kaighn, M.E.; Narayan, K.S.; Ohnuki, Y.; Lechner, J.F.; Jones, L.W. Establishment and characterization of a human prostatic carcinoma cell line (PC-3). Invest. Urol. 1979, 17, 16–23. [Google Scholar]
- Lacy, P.E.; Kostianovsky, M. Method for the isolation of intact islets of Langerhans from the rat pancreas. Diabetes 1967, 16, 35–39. [Google Scholar] [CrossRef]
Sample Availability: Samples of gintonin are available from the authors. |






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, Y.-J.; Choi, S.-H.; Lee, R.; Hwang, H.; Rhim, H.; Cho, I.-H.; Kim, H.-C.; Lee, J.-I.; Hwang, S.-H.; Nah, S.-Y. Ginseng Gintonin Contains Ligands for GPR40 and GPR55. Molecules 2020, 25, 1102. https://doi.org/10.3390/molecules25051102
Cho Y-J, Choi S-H, Lee R, Hwang H, Rhim H, Cho I-H, Kim H-C, Lee J-I, Hwang S-H, Nah S-Y. Ginseng Gintonin Contains Ligands for GPR40 and GPR55. Molecules. 2020; 25(5):1102. https://doi.org/10.3390/molecules25051102
Chicago/Turabian StyleCho, Yeon-Jin, Sun-Hye Choi, Rami Lee, Hongik Hwang, Hyewhon Rhim, Ik-Hyun Cho, Hyoung-Chun Kim, Jeong-Ik Lee, Sung-Hee Hwang, and Seung-Yeol Nah. 2020. "Ginseng Gintonin Contains Ligands for GPR40 and GPR55" Molecules 25, no. 5: 1102. https://doi.org/10.3390/molecules25051102
APA StyleCho, Y. -J., Choi, S. -H., Lee, R., Hwang, H., Rhim, H., Cho, I. -H., Kim, H. -C., Lee, J. -I., Hwang, S. -H., & Nah, S. -Y. (2020). Ginseng Gintonin Contains Ligands for GPR40 and GPR55. Molecules, 25(5), 1102. https://doi.org/10.3390/molecules25051102