
Beneficial Regulation of Cellular Oxidative Stress Effects, and Expression of Inflammatory, Angiogenic, and the Extracellular Matrix Remodeling Proteins by 1α,25-Dihydroxyvitamin D3 in a Melanoma Cell Line

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
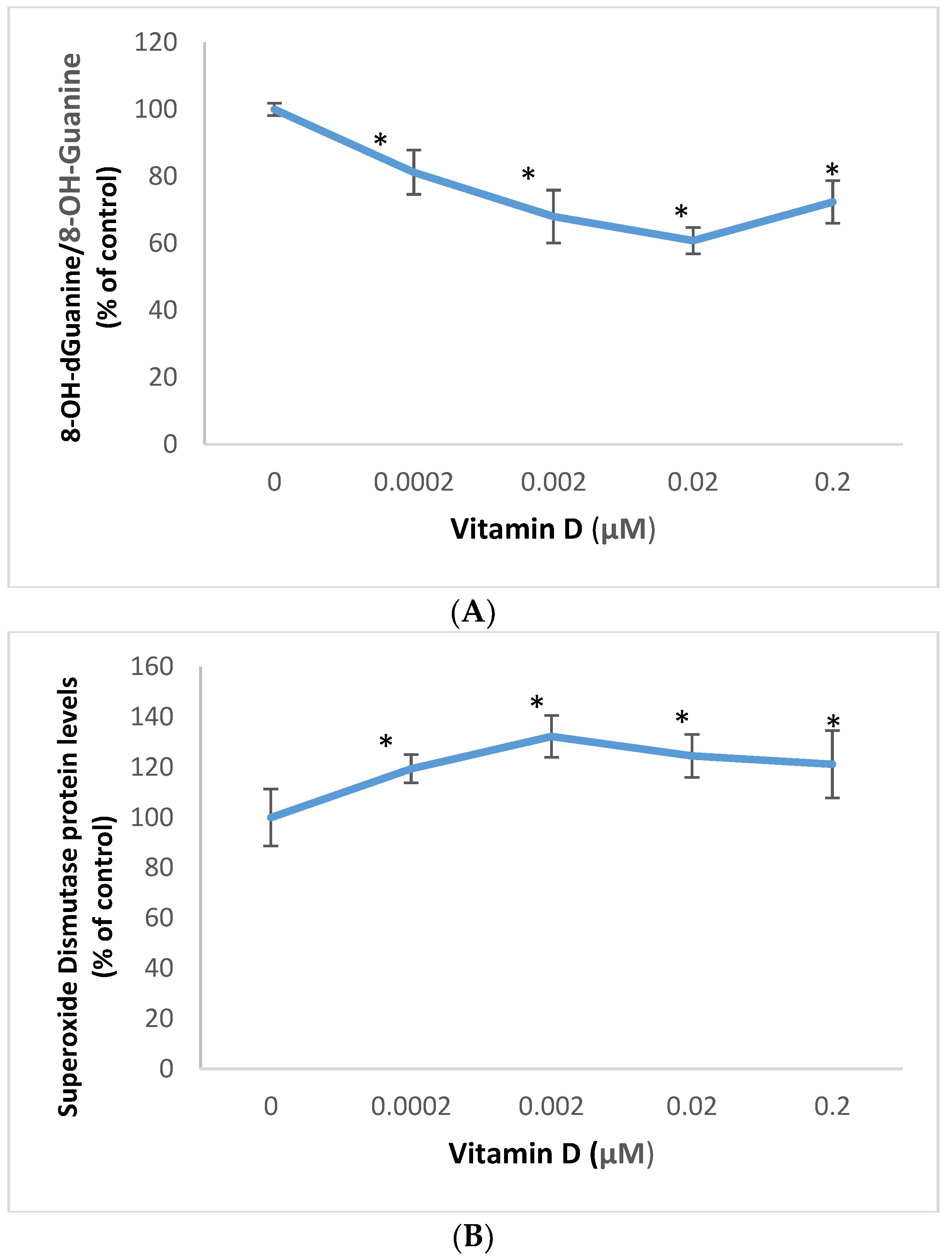
2.1. Effect of 1α,25-Dihydroxyvitamin D3 (Vitamin D) on Oxidative DNA Damage, and Superoxide Dismutase Expression in Melanoma Cells
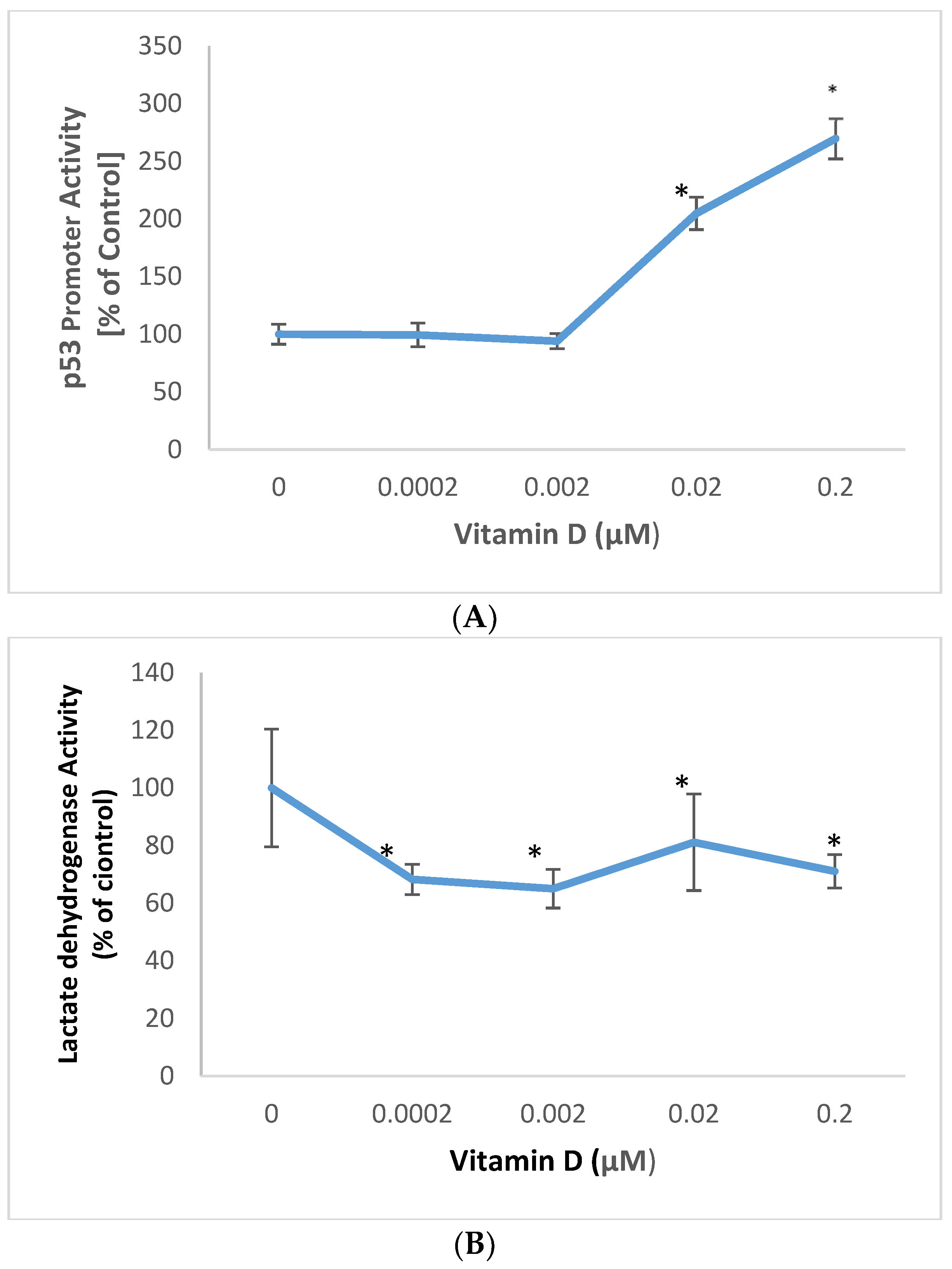
2.2. Effect of 1α,25-Dihydroxyvitamin D3 (vitamin D) on p53 Promoter Activity and Membrane Damage in Melanoma Cells
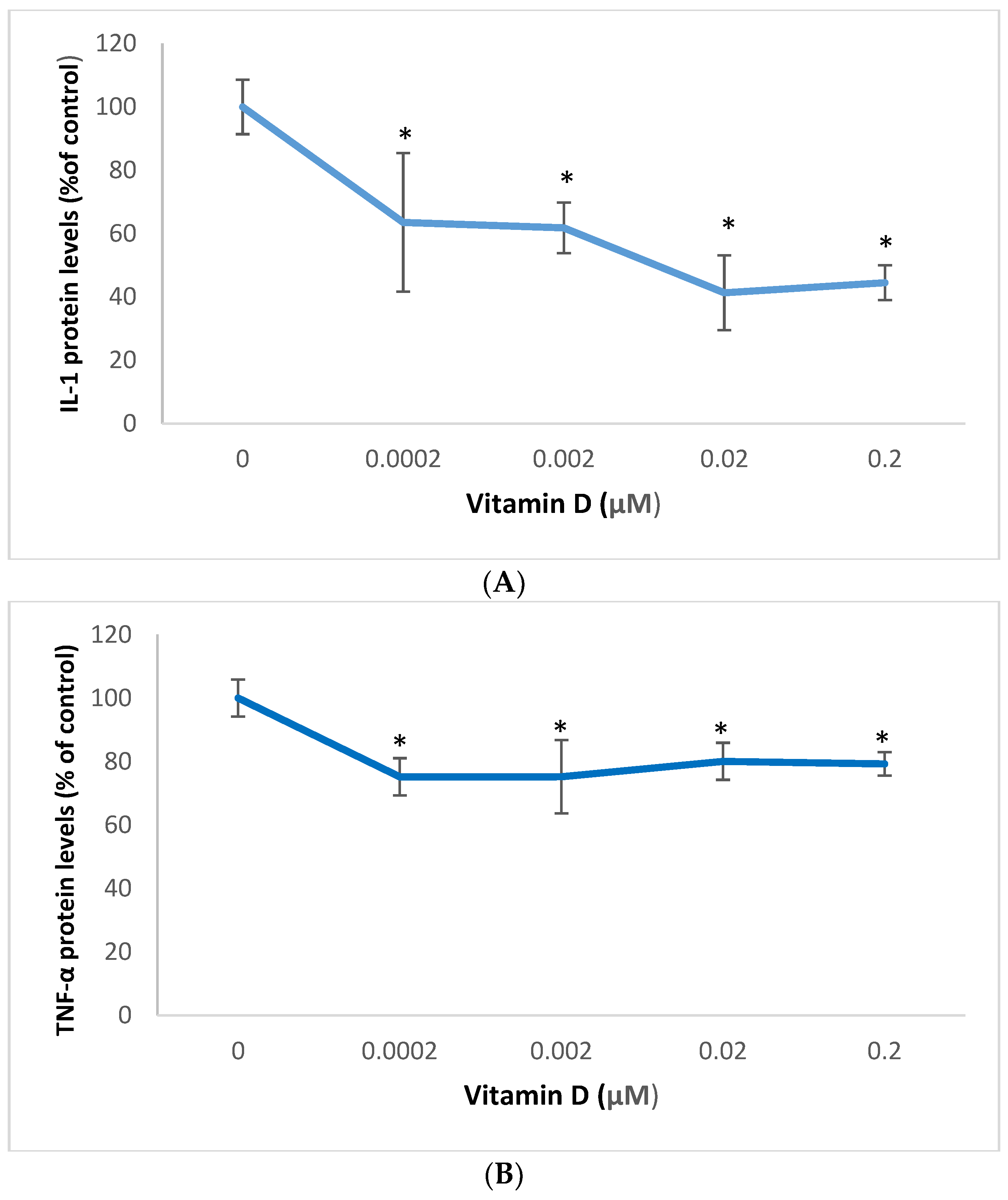
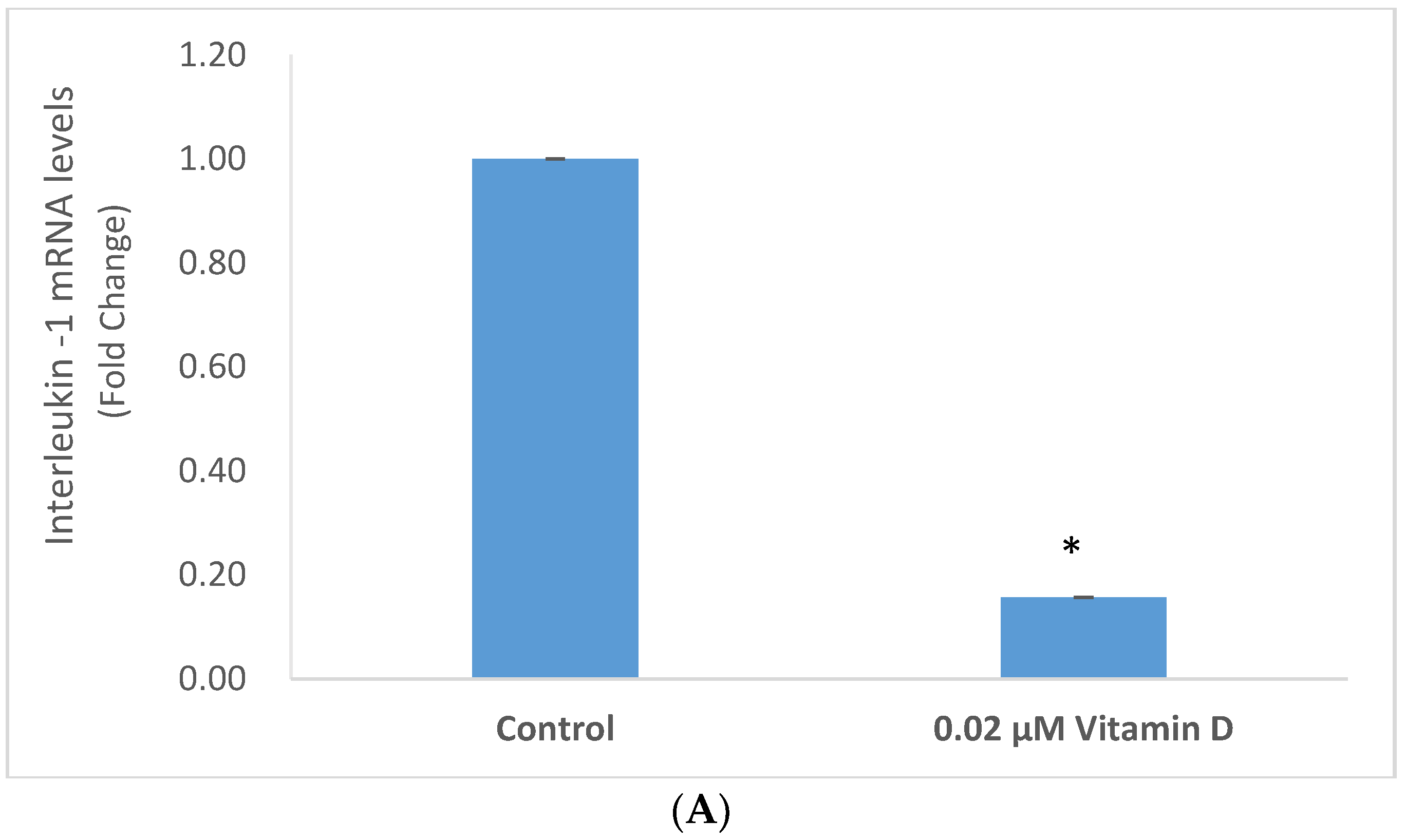
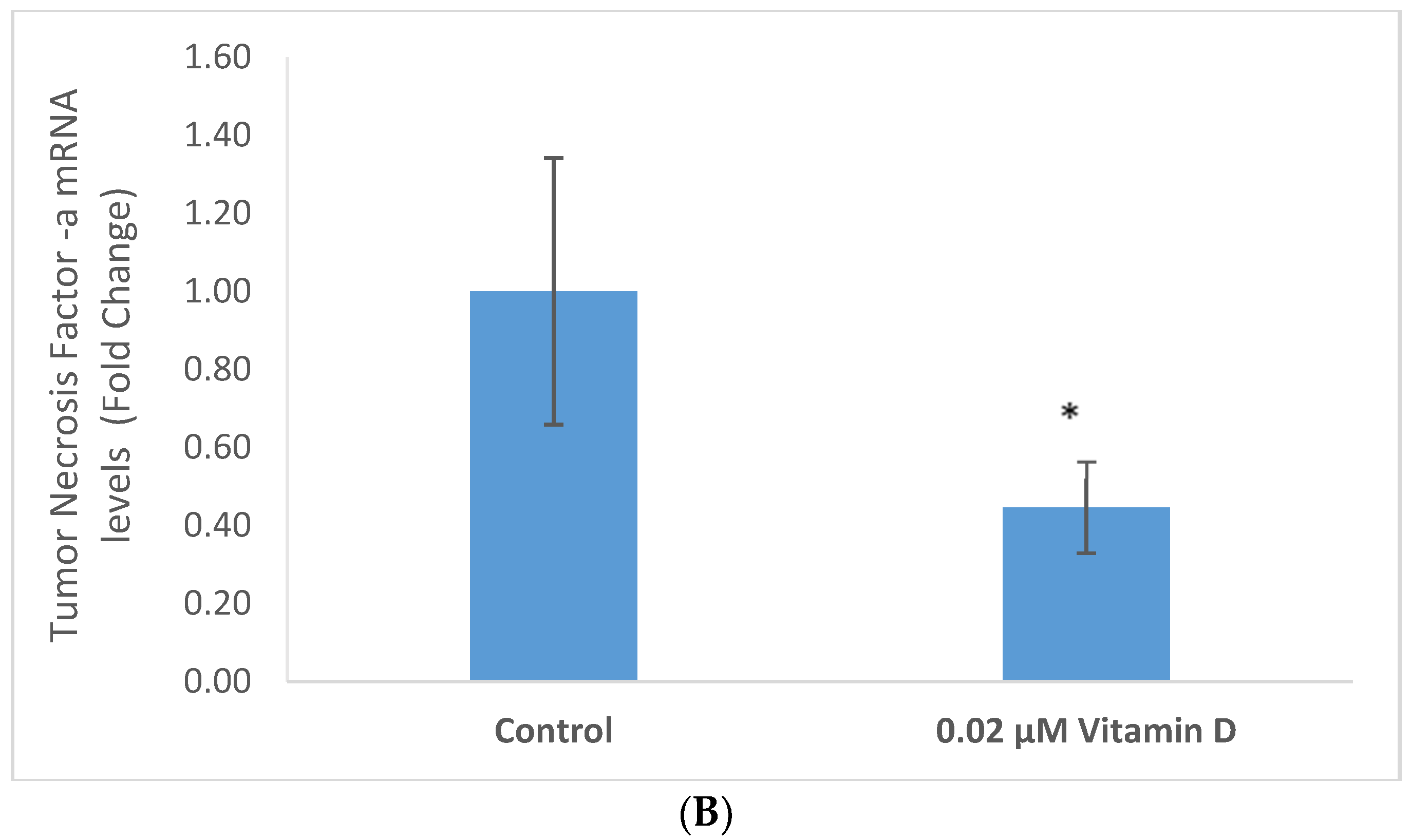
2.3. Effect of 1α,25-Dihydroxyvitamin D3 (Vitamin D) on Interleukin-1 (IL-1) and Tumor Necrosis Factor Alpha (TNF-α) in Melanoma Cells
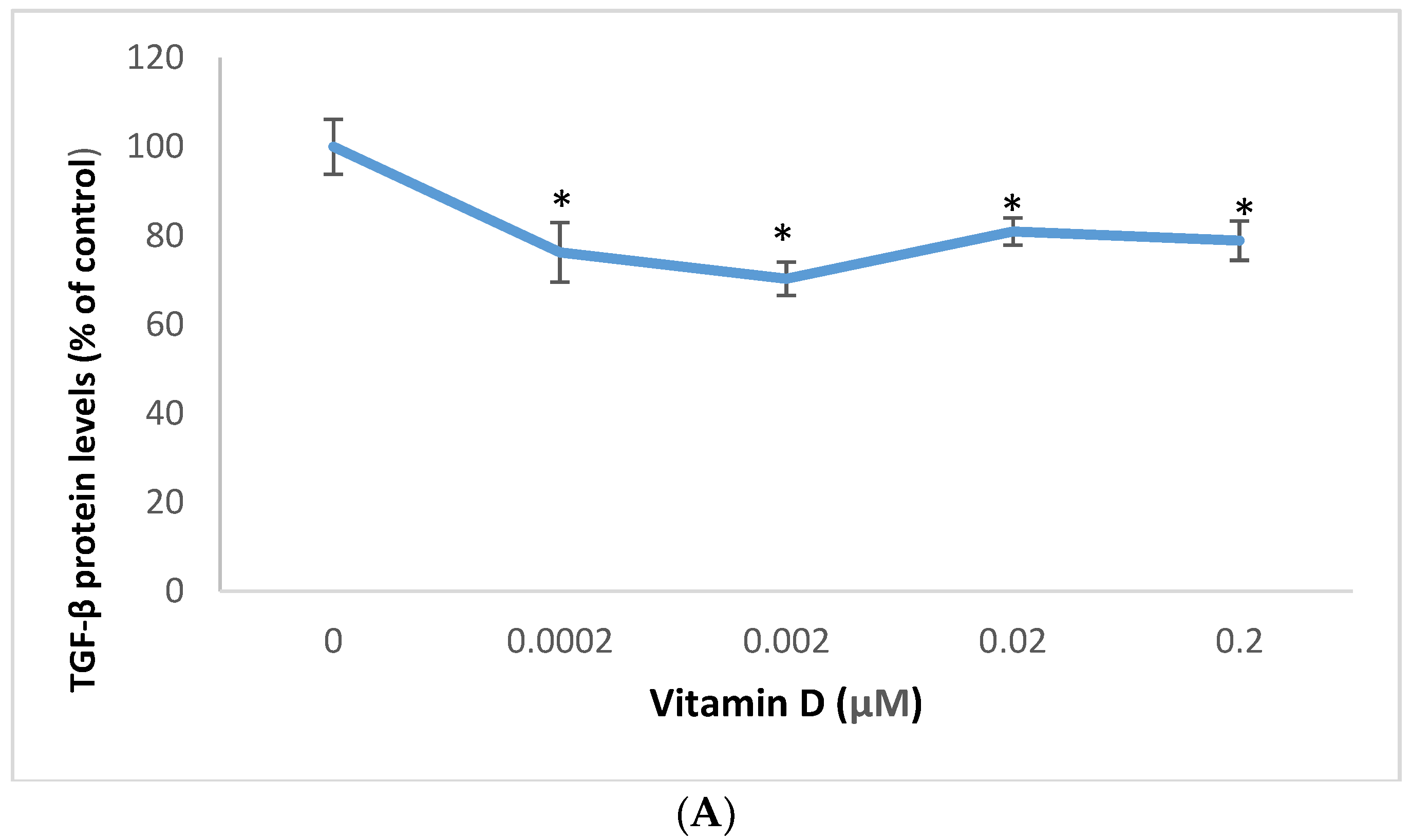
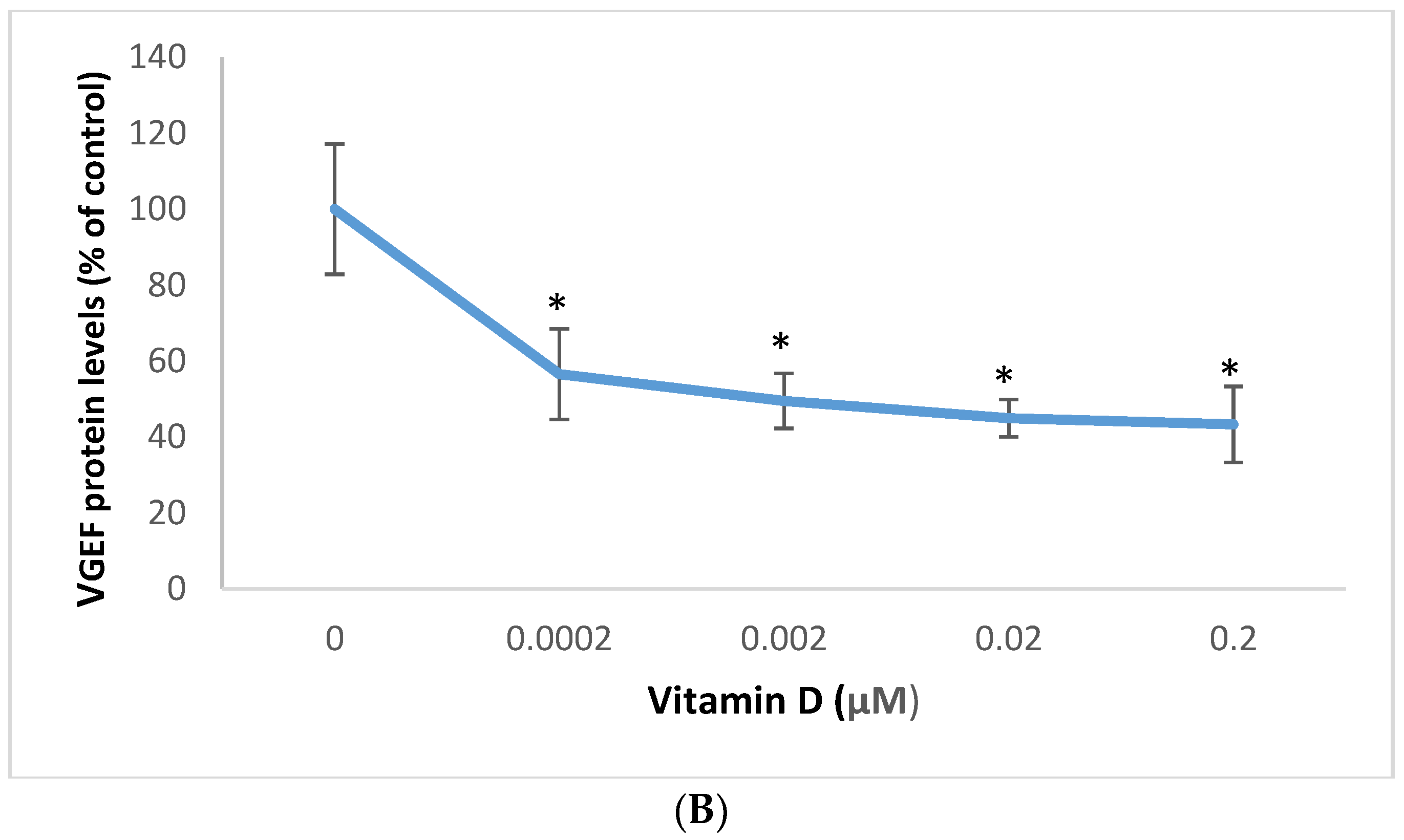
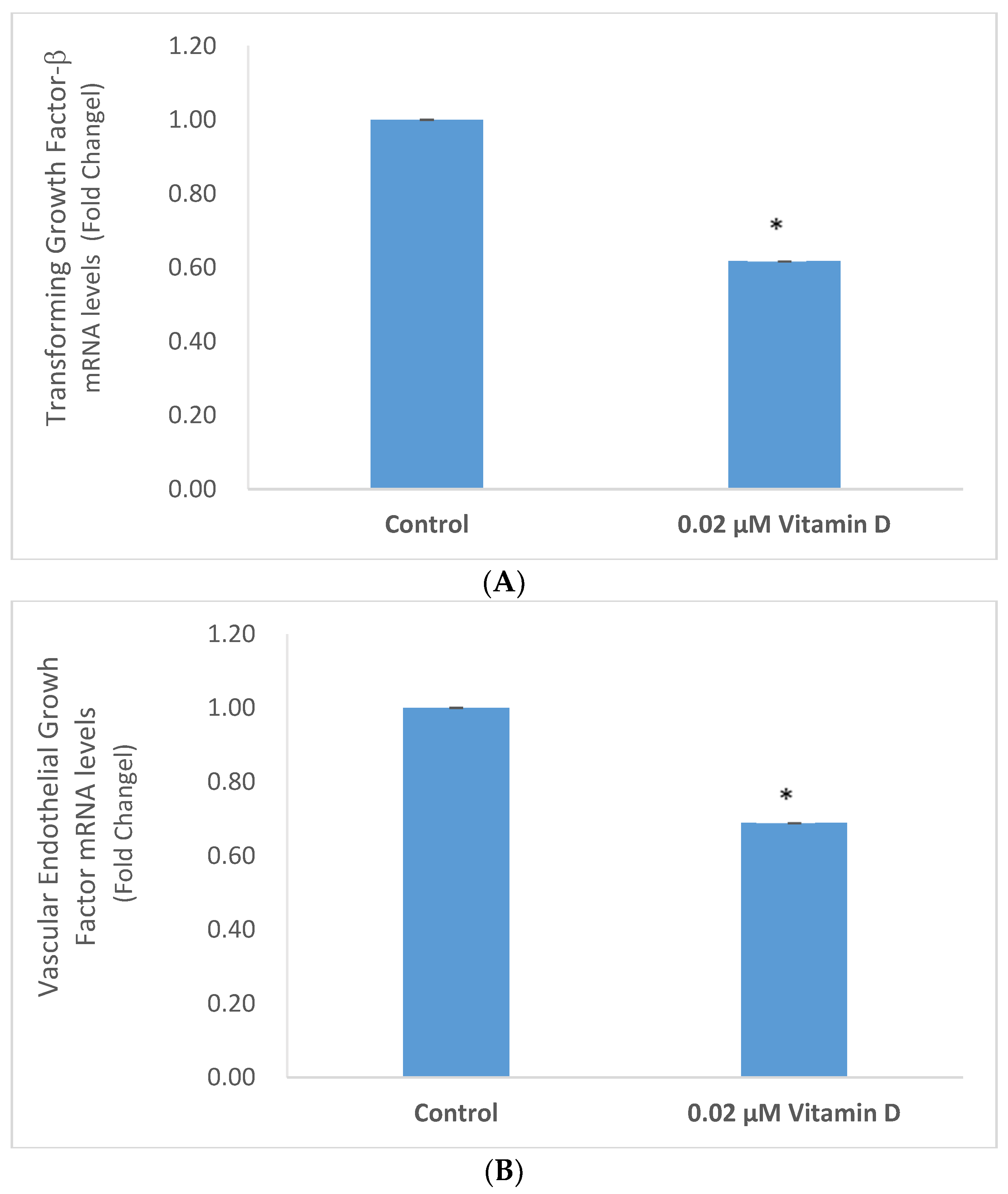
2.4. Effect of 1α,25-Dihydroxyvitamin D3 (vitamin D) on Transforming Growth Factor Beta (TGF-β), and Vascular Endothelial Growth Factor (VEGF) in Melanoma Cells
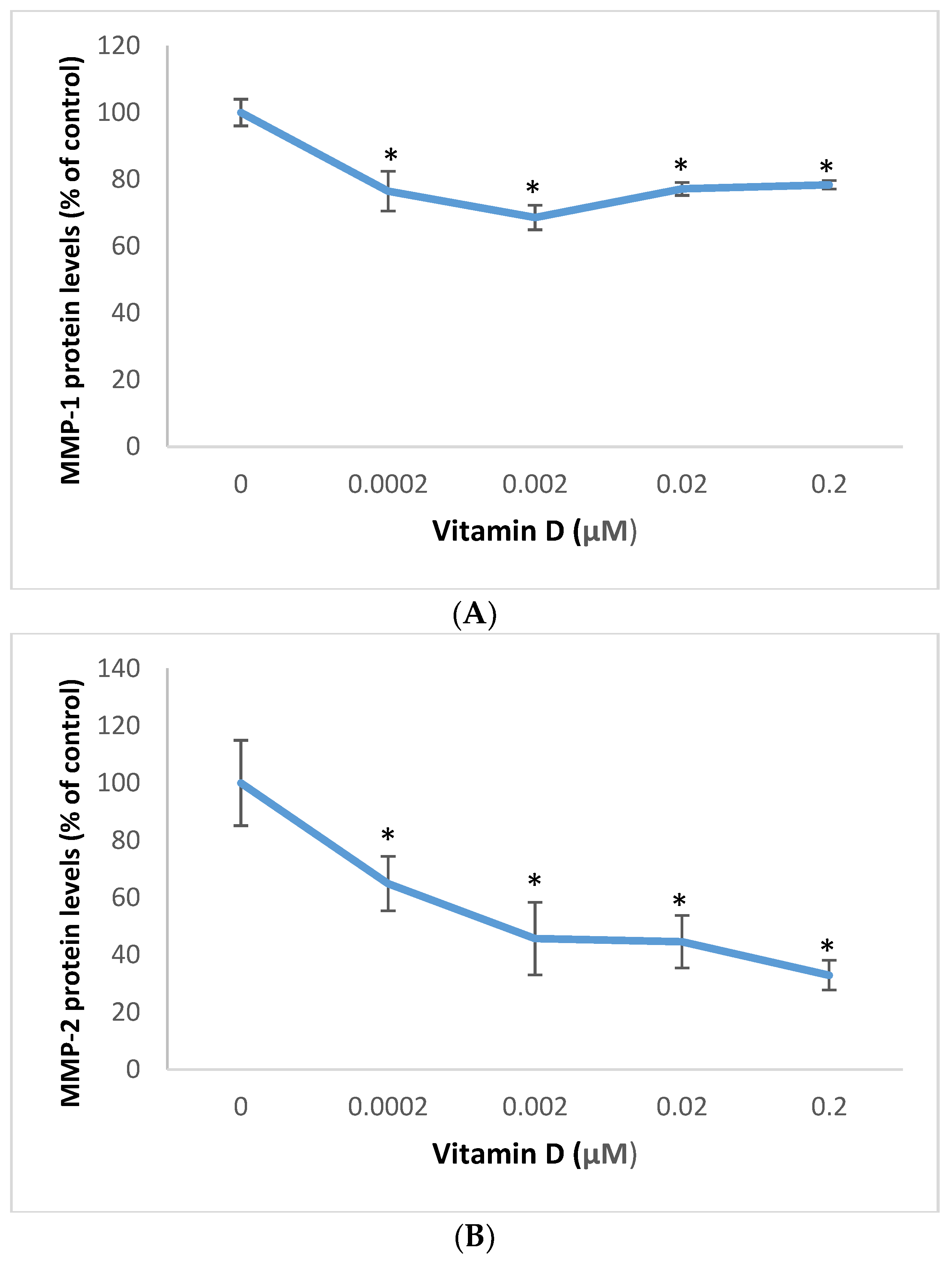
2.5. Effect of 1α,25=Dihydroxyvitamin D3 (vitamin D) on Matrixmetalloproteinase (MMP)-1 and MMP-2 in Melanoma Cells
3. Discussion
4. Method
4.1. Cell Culture/Experimental Design
4.2. Cell Viability
4.3. Oxidative DNA/RNA Damage
4.4. Protein Levels: Total Protein Content and the Protein Levels of Superoxide Dismutase (SOD), Interleukin-1 (IL-1), Tumor Necrosis Factor-α (TNF-α), Transforming Growth Factor-β (TGF-β), Vascular Endothelial Growth Factor (VEGF), Matrix Metalloproteinases (MMP)-1, and MMP-2
4.5. Promoter Activities: p53, and MMP-1
4.6. Membrane Damage
4.7. RNA Levels: IL-1, TNF-α, TGF-β, VEGF, MMP-1 and MMP-2 mRNA, and 18S RNA
4.8. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nichols, J.A.; Katiyar, S.K. Skin photoprotection by natural polyphenols: Anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Dermatol. Res. 2009, 302, 71–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briganti, S.; Picardo, M. Antioxidant activity, lipid peroxidation and skin diseases. What’s new? J. Eur. Acad. Dermatol. Venereol. 2003, 17, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Yaar, M.; Gilchrest, B.A. Photoaging: Mechanism, prevention and therapy. Br. J. Dermatol. 2007, 157, 874. [Google Scholar] [CrossRef] [PubMed]
- Che, M.; Wang, R.; Li, X.; Wang, H.; Zheng, X.S. Expanding roles of superoxide dismutases in cell regulation and cancer. Drug Discov. Today 2016, 21, 143–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaunig, J.E.; Wang, Z.; Pu, X.; Zhou, S. Oxidative stress and oxidative damage in chemical carcinogenesis. Toxicol. Appl. Pharm. 2011, 254, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Morry, J.; Ngamcherdtrakul, W.; Yantasee, W. Oxidative stress in cancer and fibrosis: Opportunity for therapeutic intervention with antioxidant compounds, enzymes, and nanoparticles. Redox Biol. 2017, 11, 240–253. [Google Scholar] [CrossRef]
- Bosch, R.; Philips, N.; Suárez-Pérez, J.; Juarranz, A.; Devmurari, A.; Chalensouk-Khaosaat, J.; González, S. Mechanisms of Photoaging and Cutaneous Photocarcinogenesis, and Photoprotective Strategies with Phytochemicals. Antioxidants 2015, 4, 248–268. [Google Scholar] [CrossRef] [Green Version]
- Philips, N.; Burchill, D.; O’Donoghue, D.; Keller, T.; Gonzalez, S. Identification of Benzene Metabolites in Dermal Fibroblasts as Nonphenolic: Regulation of Cell Viability, Apoptosis, Lipid Peroxidation and Expression of Matrix Metalloproteinase 1 and Elastin by Benzene Metabolites. Skin Pharmacol. Physiol. 2004, 17, 147–152. [Google Scholar] [CrossRef]
- Philips, N.; Chalensouk-Khaosaat, J.; Devmurari, A.; Patel, H. Polyphenolic nanobiomaterials as emerging therapies for combating physiology and clinical aspects of photoaging and photocarcinogenesis. In Skin Aging and Photoaging: Physiology, Clinical Aspects and Emerging Therapies; Nova Publishers Inc.: Hauppauge, NY, USA, 2015. [Google Scholar]
- Philips, N.; Siomyk, H.; Bynum, D.; Gonzalez, S. Skin Cancer, Polyphenols, and Oxidative Stress. In Cancer: Oxidative Stress and Dietary Antioxidants; Preedy, V.R., Ed.; Academic Press: London, UK, 2014; pp. 265–270. [Google Scholar]
- Parrado, C.; Juarranz, A.; Gilaberte, Y.; Philips, N.; Gonzalez, S. Fern extract, oxidative stress and skin cancer. In Cancer: Oxidative Stress and Dietary Antioxidants; Preedy, V.R., Ed.; Academic Press: London, UK, 2014. [Google Scholar]
- Philips, N.; Samuel, M.; Parakandi, H.; Siomyk, H.; Re, M.; Gopal, S.; Jia, H.; Shahin, H. Vitamins in the Therapy of Inflammatory and Oxidative Diseases. In Frontiers in Clinical Drug Research—Anti Allergy Agents; Bentham Science Publishers: Sharjah, UAE, 2013; pp. 240–264. [Google Scholar]
- Kindt, T.J.; Goldsby, R.A.; Osborne, B.A. Kuby Immunology; W. H. Freeman and Company: New York, NY, USA, 2007. [Google Scholar]
- Philips, N.; Samuel, P.; Samuel, M.; Perez, G.; Khundoker, R.; Alahmade, G. Interleukin-4 Signaling Pathway and Effects in Allergic Diseases. Curr. Signal Transduct. Ther. 2018, 13, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Philips, N.; Samuel, M. Inhibition of interleukin-4 signaling in the treatment of atopic dermatitis and allergic asthma. Glob. J. Allergy 2017, 3, 19–21. [Google Scholar] [CrossRef] [Green Version]
- Philips, N.; Bynum, D.; Hwang, H. Counteraction of Skin Inflammation and Aging or Cancer by Polyphenols and Flavonoids from Polypodium leucotomos and Xanthohumol. Anti Inflamm. Anti Allergy Agents. Med. Chem. 2010, 9, 142–149. [Google Scholar] [CrossRef]
- Philips, N.; Siomyk, H. Inhibition of Angiogenesis in Cancer Management by Antioxidants: Ascorbate and P. leucotomos. In Anti-Angiogenesis Drug Discovery and Development; Bentham Science Publishers: Sharjah, UAE, 2014; pp. 132–146. [Google Scholar]
- Philips, N. Reciprocal effects of ascorbate on cancer cell growth and the expression of matrix metalloproteinases and transforming growth factor-beta: Modulation by gene silencing or P. leucotomos. In Handbook of Vitamin C Research: Daily Requirements, Dietary Sources, and Adverse Effects; Nova Science: Hauppauge, NY, USA, 2009. [Google Scholar]
- Philips, N.; Samuel, P.; Siomyk, H.; Parakandi, H.; Gopal, S.; Shahin, H. Improved cell metabolism and strengthening of the extracellular matrix by nicotinamide, and copper for anti-skin aging. In Skin Aging: New Research; Nova Science: Hauppauge, NY, USA, 2012. [Google Scholar]
- Lodish, H.; Berk, A.; Kaiser, C.A.; Krieger, M.; Scott, M.P.; Bretscher, A.; Ploegh, H.; Matsudaira, P. Molecular Cell Biology; W.H. Freeman and Company: New York, NY, USA, 2016. [Google Scholar]
- Astner, S.; Wu, A.; Chen, J.; Philip, N.; Rius-Diaz, F.; Parrado, C.; Mihm, M.C.; Goukassian, D.A.; Pathak, M.A.; González, S. Dietary lutein/zeaxanthin reduces photoaging and photocarcinogenesis in chronically UVB irradiated SKH -1 hairless mice. Skin Pharmacol. Physiol. 2007, 20, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Philips, N.; Samuel, P.; Lozano, T.; Gvaladze, A.; Guzman, B.; Siomyk, H.; Haas, G. Effects of Humulus lupulus extract or its Components on Viability, Lipid Peroxidation, and expression of Vascular Endothelial Growth Factor in Melanoma Cells and Fibroblasts. Madridge J. Clin. Res. 2017, 1, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Philips, N.; Chalensouk-Khaosaat, J.; Gonzalez, S. Stimulation of the Fibrillar Collagen and Heat Shock Proteins by Nicotinamide or Its Derivatives in Non-Irradiated or UVA Radiated Fibroblasts, and Direct Anti-Oxidant Activity of Nicotinamide Derivatives. Cosmetics 2015, 2, 146–161. [Google Scholar] [CrossRef]
- Philips, N.; Gonzalez, S. Beneficial Regulation of Elastase Activity and Expression of Tissue Inhibitors of matrix metalloproteinases, Fibrillin, Transforming Growth Factor-β, and Heat Shock Proteins by P. leucotomos in Nonirradiated or Ultraviolet-Radiated Epidermal Keratinocytes. ISRN Oxidative Med. 2013, 2013, 1–7. [Google Scholar] [CrossRef]
- Philips, N.; Samuel, M.; Arena, R.; Chen, Y.; Conte, J.; Natrajan, P.; Haas, G.; Gonzalez, S. Direct inhibition of elastase and matrix metalloproteinases, and stimulation of biosynthesis of fibrillar collagens, elastin and fibrillins by xanthohumol. J. Cosmet. Sci. 2010, 61, 125–132. [Google Scholar]
- Philips, N.; Conte, J.; Chen, Y.-J.; Natrajan, P.; Taw, M.; Keller, T.; Givant, J.; Tuason, M.; Dulaj, L.; Leonardi, D.; et al. Beneficial regulation of matrix metalloproteinases and their inhibitors, fibrillar collagens and transforming growth factor-β by Polypodium leucotomos, directly or in dermal fibroblasts, ultraviolet radiated fibroblasts, and melanoma cells. Arch. Dermatol. Res. 2009, 301, 487–495. [Google Scholar] [CrossRef]
- Philips, N.; Smith, J.; Keller, T.; Gonzalez, S. Predominant effects of Polypodium leucotomos on membrane integrity, lipid peroxidation, and expression of elastin and matrix metalloproteinase-1 in ultraviolet radiation exposed fibroblasts, and keratinocytes. J. Dermatol. Sci. 2003, 32, 1–9. [Google Scholar] [CrossRef]
- Philips, N.; Keller, T.; Hendrix, C.; Hamilton, S.; Arena, R.; Tuason, M.; Gonzalez, S. Regulation of the extracellular matrix remodeling by lutein in dermal fibroblasts, melanoma cells, and ultraviolet radiation exposed fibroblasts. Arch. Dermatol. Res. 2007, 8, 373–379. [Google Scholar] [CrossRef]
- Philips, N.; Ding, X.; Kandalai, P.; Marte, I.; Krawczyk, H.; Richardson, R. The Beneficial Regulation of Extracellular Matrix and Heat Shock Proteins, and the Inhibition of Cellular Oxidative Stress Effects and Inflammatory Cytokines by 1α,25-dihydroxyvitamin D3 in Non-Irradiated and Ultraviolet Radiated Dermal Fibroblasts. Cosmetics 2019, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Garland, C.F.; Garland, F.C.; Gorham, E.D.; Lipkin, M.; Newmark, H.; Mohr, S.B.; Holick, M.F. The role of vitamin D in cancer prevention. Am. J. Public Health 2006, 96, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicka, J.M.; Żmijewski, M.A.; Piotrowska, A.; Nedoszytko, B.; Lange, M.; Tuckey, R.C.; Slominski, A.T. Bioactive forms of vitamin D selectively stimulate the skin analog of the hypothalamus-pituitary-adrenal axis in human epidermal keratinocytes. Mol. Cell Endocrinol. 2016, 437, 312–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.T.; Janjetovic, Z.; Kim, T.K.; Wright, A.C.; Grese, L.N.; Riney, S.J.; Nguyen, M.N.; Tuckey, R.C. Novel vitamin D hydroxyderivatives inhibit melanoma growth and show differential effects on normal melanocytes. Anticancer Res. 2012, 32, 3733–3742. [Google Scholar] [PubMed]
- Feldman, D.; Krishnan, A.V.; Swami, S.; Giovannucci, E.; Feldman, B.J. The role of vitamin D in reducing cancer risk and progression. Nat. Rev. Cancer 2014, 14, 342–357. [Google Scholar] [CrossRef]
- Gordon-Thomson, C.; Gupta, R.; Tongkao-On, W.; Ryan, A.; Halliday, G.M.; Mason, R.S. 1α,25 Dihydroxyvitamin D3 enhances cellular defences against UV-induced oxidative and other forms of DNA damage in skin. Photochem. Photobiol. Sci. 2012, 11, 1837. [Google Scholar] [CrossRef]
- Ke, C.-Y.; Yang, F.-L.; Wu, W.-T.; Chung, C.-H.; Lee, R.-P.; Yang, W.-T.; Liao, K.-W. Vitamin D3 Reduces Tissue Damage and Oxidative Stress Caused by Exhaustive Exercise. Int. J. Med. Sci. 2016, 13, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Keisala, T.; Minasyan, A.; Lou, Y.R.; Zou, J.; Kalueff, A.V.; Pyykkö, I.; Tuohimaa, P. Premature aging in vitamin D receptor mutant mice. J. Steroid Biochem. Mol. Biol. 2009, 115, 91–97. [Google Scholar] [CrossRef]
- Wei, R.; Christakos, S. Mechanisms Underlying the Regulation of Innate and Adaptive Immunity by Vitamin D. Nutrients 2015, 7, 8251–8260. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Xu, T. Expression of vitamin D receptor with inflammatory changes and prognosis of asthma. Expt. Ther. Med. 2018, 16, 5096–5102. [Google Scholar] [CrossRef] [Green Version]
- Cannell, J.J.; Grant, W.B.; Holick, M.F. Vitamin D and inflammation. Dermatoendocrinology 2015, 6, e983401. [Google Scholar] [CrossRef] [Green Version]
- Balato, A.; Schiattarella, M.; Lembo, S.; Mattii, M.; Prevete, N.; Balato, N.; Ayala, F. Interleukin-1 family members are enhanced in psoriasis and suppressed by vitamin D and retinoic acid. Arch. Dermatol. Res. 2013, 305, 255–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen-Lahav, M.; Shany, S.; Tobvin, D.; Chaimovitz, C.; Douvdevani, A. Vitamin D decreases NFκB activity by increasing IκBα levels. Nephrol. Dial. Transplant. 2006, 21, 889–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantell, D.J.; Owens, P.E.; Bundred, N.J.; Mawer, E.B.; Canfield, A.E. 1 α,25- dihydroxyvitamin D(3) inhibits angiogenesis in vitro and in vivo. Circ Res. 2000, 87, 214–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iseki, K.; Tatsuta, M.; Uehara, H.; Iishi, H.; Yano, H.; Sakai, N.; Ishiguro, S. Inhibition of angiogenesis as a mechanism for inhibition by 1α- hydroxyvitamin D3 and 1,25-dihydrox-y vitamin D3 of colon carcinogenesis induced by azoxymethane in Wistar rats. Int. J. Cancer 1999, 81, 730–733. [Google Scholar] [CrossRef]
- Kim, S.; Baek, M.; Yoon, D.; Park, J.; Yoon, B.; Oh, B.; Kim, H. Vitamin D Inhibits Expression and Activity of Matrix Metalloproteinase in Human Lung Fibroblasts (HFL-1) Cells. Tuberc. Respir. Dis. 2014, 77, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Halder, S.K.; Osteen, K.G.; Al-Hendy, A. Vitamin D3 inhibits expression and activities of matrix metalloproteinase-2 and -9 in human uterine fibroid cells. Hum. Reprod. 2013, 28, 2407–2416. [Google Scholar] [CrossRef] [Green Version]
- Philips, N.; Keller, T.; Holmes, C. Reciprocal effects of ascorbate on cancer cell growth and the expression of matrix metalloproteinases and transforming growth factor-β. Cancer Lett. 2007, 256, 49–55. [Google Scholar] [CrossRef]
- Philips, N.; Samuel, P.; Parakandi, H.; Gopal, S.; Siomyk, H.; Ministro, A.; Thompson, T.; Borkow, G. Beneficial Regulation of Fibrillar Collagens, Heat Shock Protein-47, Elastin Fiber Components, Transforming Growth Factor-β1, Vascular Endothelial Growth Factor and Oxidative Stress Effects by Copper in Dermal Fibroblasts. Connect. Tissue Res. 2012, 53, 373–378. [Google Scholar] [CrossRef]
- Philips, N.; Hwang, H.; Chauhan, S.; Leonardi, D.; Gonzalez, S. Stimulation of cell proliferation, and expression of matrixmetalloproteinase-1 and interluekin-8 genes in dermal fibroblasts by copper. Connect Tissue Res. 2010, 51, 224–229. [Google Scholar] [CrossRef]
- Philips, N.; Tuason, M.; Chang, T.; Lin, Y.; Tahir, M.; Rodriguez, S. Differential Effects of Ceramide on Cell Viability and Extracellular Matrix Remodeling in Keratinocytes and Fibroblasts. Skin Pharmacol. Physiol. 2009, 22, 151–157. [Google Scholar] [CrossRef]
- Philips, N.; Mcfadden, K. Inhibition of transforming growth factor-beta and matrix metalloproteinases by estrogen and prolactin in breast cancer cells. Cancer Lett. 2004, 206, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Philips, N. An anti TGF-β increased the expression of transforming growth factor-β, matrix metallproteinase-1, and elastin, and its effects were antagonized by ultraviolet radiation in epidermal keratinocytes. J. Dermatol. Sci. 2003, 33, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Philips, N.; Tahir, M.; Stellatella, J.; Stephan, K.; Givant, J.; Zhou, L.; Costa, A.; Taw, M. Differential regulation of growth factors and matrix metalloproteinase-1 by estrogen, progesterone, and tamoxifen in normal and cancerous endometrial cells. J. Cancer Mol. 2009, 4, 169–173. [Google Scholar]
- Karroum, A.; Mirshahi, P.; Faussat, A.; Therwath, A.; Mirshahi, M.; Hatmi, M. Tubular network formation by adriamycin-resistant MCF-7 breast cancer cells is closely linked to MMP-9 and VEGFR-2/VEGFR-3 over-expressions. Eur. J. Pharmacol. 2012, 685, 1–7. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: The research material is available commercially. |









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Philips, N.; Samuel, P.; Keller, T.; Alharbi, A.; Alshalan, S.; Shamlan, S.-A. Beneficial Regulation of Cellular Oxidative Stress Effects, and Expression of Inflammatory, Angiogenic, and the Extracellular Matrix Remodeling Proteins by 1α,25-Dihydroxyvitamin D3 in a Melanoma Cell Line. Molecules 2020, 25, 1164. https://doi.org/10.3390/molecules25051164
Philips N, Samuel P, Keller T, Alharbi A, Alshalan S, Shamlan S-A. Beneficial Regulation of Cellular Oxidative Stress Effects, and Expression of Inflammatory, Angiogenic, and the Extracellular Matrix Remodeling Proteins by 1α,25-Dihydroxyvitamin D3 in a Melanoma Cell Line. Molecules. 2020; 25(5):1164. https://doi.org/10.3390/molecules25051164
Chicago/Turabian StylePhilips, Neena, Philips Samuel, Thomas Keller, Asma Alharbi, Samar Alshalan, and Sara-Ali Shamlan. 2020. "Beneficial Regulation of Cellular Oxidative Stress Effects, and Expression of Inflammatory, Angiogenic, and the Extracellular Matrix Remodeling Proteins by 1α,25-Dihydroxyvitamin D3 in a Melanoma Cell Line" Molecules 25, no. 5: 1164. https://doi.org/10.3390/molecules25051164
APA StylePhilips, N., Samuel, P., Keller, T., Alharbi, A., Alshalan, S., & Shamlan, S. -A. (2020). Beneficial Regulation of Cellular Oxidative Stress Effects, and Expression of Inflammatory, Angiogenic, and the Extracellular Matrix Remodeling Proteins by 1α,25-Dihydroxyvitamin D3 in a Melanoma Cell Line. Molecules, 25(5), 1164. https://doi.org/10.3390/molecules25051164