Abstract
Bacterioruberin and its derivatives have been described as the major carotenoids produced by haloarchaea (halophilic microbes belonging to the Archaea domain). Recently, different works have revealed that some haloarchaea synthetize other carotenoids at very low concentrations, like lycopene, lycopersene, cis- and trans-phytoene, cis- and trans-phytofluene, neo-β-carotene, and neo-α-carotene. However, there is still controversy about the nature of the pathways for carotenogenesis in haloarchaea. During the last decade, the number of haloarchaeal genomes fully sequenced and assembled has increased significantly. Although some of these genomes are not fully annotated, and many others are drafts, this information provides a new approach to exploring the capability of haloarchaea to produce carotenoids. This work conducts a deeply bioinformatic analysis to establish a hypothetical metabolic map connecting all the potential pathways involved in carotenogenesis in haloarchaea. Special interest has been focused on the synthesis of bacterioruberin in members of the Haloferax genus. The main finding is that in almost all the genus analyzed, a functioning alternative mevalonic acid (MVA) pathway provides isopentenyl pyrophosphate (IPP) in haloarchaea. Then, the main branch to synthesized carotenoids proceeds up to lycopene from which β-carotene or bacterioruberin (and its precursors: monoanhydrobacterioriberin, bisanhydrobacterioruberin, dihydrobisanhydrobacteriuberin, isopentenyldehydrorhodopsin, and dihydroisopenthenyldehydrorhodopsin) can be made.
1. Introduction
Carotenoids are widespread isoprenoid pigments in nature synthesized by bacteria, archaea, plants, algae, and yeasts [1,2]. Their main roles range from acting as accessory pigments in photosynthesis and as antioxidants affecting the regulation of gene expression [3]. Carotenoids are well-known for their color and potential beneficial effects on human health, and for that reason, they are frequently used in medical, nutraceutical, and pharmaceutical industries. These compounds are generally produced by chemical synthesis; however, the interest in the use of natural sources for carotenoid production is currently increasing [4,5,6].
Carotenoids are usually classified into three categories regarding the number of carbons present in their carotene backbones: C30, C40, and C50. Most carotenoids belong to the C40 group, as it is in the case of β-carotene. C30 and C50 carotenoids represent smaller percentages, especially the C50 category. C50 carotenoid structure results from the addition of two 5-carbon isoprene units to the C40 backbone [7]. So far, only the biosynthesis pathway of three C50 carotenoids has been determined: the ε-cyclic C50 carotenoid decaprenoxanthin [8], the β-cyclic carotenoid from Corynebacterium poinsettiae [9], and the γ-cyclic C50 carotenoid from Micrococcus luteus [10]. C50 carotenoids show higher antioxidative properties, given their longer conjugated double bonds and the presence of at least one hydroxyl group [11]. Therefore, this rare group of carotenoids is of interest to a wide range of industrial applications. The synthesis of C50 carotenoids has been barely studied, especially in the Archaea domain [7,12], whilst studies regarding plant, algae and bacteria C30 and C40 synthesis pathways are abundant in the literature [13,14,15].
Halophiles are a type of extremophile organism that requires high concentrations of salts for optimal growth, and that can be found in all three domains of life: Archaea, Bacteria, and Eukarya [16,17]. Within the Archaea domain, haloarchaea are those microbes showing mid or high requirements of salt to be alive. They are mainly grouped into two families: Haloferacaceae and Halobacteriaceae [18,19].
Haloferax is a halophilic genus that, in most cases, is pink-red colored given the production of C50 carotenoids, mainly bacterioruberin and its derivatives. These microorganisms also produce C40 carotenoids, although in lower percentages [7,16]. The acyclic C50 carotenoid bacterioruberin has been frequently observed in lipid membranes of halophilic archaea since these carotenoids are part of their mechanism of defense against salinity and temperature changes or sun radiation environments [20]. There is a clear consensus on the fact that most members of the families Halobacteriaceae and Haloferacaceae (to which Haloferax genus belong to) are able to synthesize C50 carotenoids, particularly bacterioruberin [20,21,22,23,24,25,26,27]. These characteristics make it interesting to suggest haloarchaeal species, such as Haloferax members, as good natural sources for the biosynthesis of carotenoids, particularly of those made of C50 backbones [16]. However, there is a lack of knowledge of carotenogenesis in haloarchaea.
Falb and co-workers addressed a systematic metabolic reconstruction and comparative analysis of four completely sequenced genomes from haloarchaeal species in 2008 (Halobacterium salinarum, Haloarcula marismortui, Haloquadratum walsbyi, and the haloalkalophile Natronomonas pharaonic) [20]. In this study, the authors stated that the isoprenoid precursor isopentenyl pyrophosphate (IPP) is synthesized via the mevalonate pathway. It was also demonstrated that various isoprenoids detected in membranes of H. salinarum are synthesized by a series of condensation reactions with IPP, which is added in head-tail or head–head fashion, and through desaturase reactions. They also concluded that the enzymatic gene set for isoprenoid synthesis differs only slightly between the haloarchaea compared [20].
More recently, it was reported that lycopene is the branching point for bacterioruberin synthesis, although the reactions involved in this step in the Haloferax genus have not been brought to light yet. Moreover, experiments on Haloarcula japonica have led to the identification of three genes: c0507, c0506, and c0505; encoding respectively the following enzymes: carotenoid 3,4-desaturase (CrtD), a bifunctional lycopene elongase and 1,2-hydratase (LyeJ), and a C50 carotenoid 2″,3″-hydratase (CruF). These enzymes are responsible for the conversions from lycopene to bacterioruberin in Haloarcula japonica [12].
Considering these last findings and in order to optimize C50 carotenoid production using Haloferax sp. as a natural source, it is mandatory to assess the identification of the C50 biosynthesis pathway. The reason for using Haloferax sp. is because this is one of the best-characterized haloarchaeal genera from a biochemical and physiological point of view. Thus, this work conducts a deeply bioinformatic analysis to establish a hypothetical metabolic map connecting all the potential pathways involved in carotenogenesis in haloarchaea. A potential pathway for the synthesis of bacterioruberin in the genus Haloferax is also explored in detail. This proposal will open new research lines promoting the characterization of carotenogenesis in haloarchaea as well as the production of mutants able to overproduce carotenoids for biotechnological purposes like cosmetics, pharmacy and biomedicine.
2. Results
2.1. Reconstruction of a Metabolic Map for Global Carotenogenesis.
Due to the lack of specific information related to carotenogenesis in haloarchaea, the first step in this work was to reconstruct a metabolic map including all the pathways described so far from different type of organisms (Eukarya, Bacteria, and Archaea), by integrating all the information available at BRENDA, KEGG, MetaCyc, NCBI, and Uniprot. The result of this integration is displayed in Figure 1.
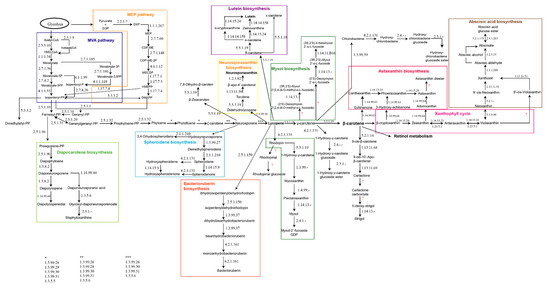
Figure 1.
Metabolic map for global carotenogenesis. The enzymes catalyzing each reaction are indicated using their respective EC numbers (Enzyme commission numbers). Interrogation marks indicate unidentified enzymes.
The initiation of the carotenogenesis depends on the general isoprenoid biosynthetic pathway, along with a variety of other important natural substances such as steroids and gibberellic acid. Thus, the starting product required to synthetize all the isoprene derivatives is in general mevalonic acid, which is transformed into phosphorylated isoprene upon phosphorylation; this isoprene subsequently polymerizes. The cytosol-localized mevalonic acid (MVA) pathway delivers the basic isoprene unit isopentenyl diphosphate (IPP) in most of the organisms. However, in higher plants, this central metabolic intermediate is also synthesized by the plastid-localized methylerythritol phosphate (MEP) pathway. Both MVA and MEP pathways conspire through an exchange of intermediates and regulatory interactions (Figure 1, left panel) [28]. In the course of isoprene polymerization, the number and position of the double bonds are fixed [16]. Then, the conversion of two molecules of geranylgeranyl pyrophosphate (GGPP) to phytoene, a compound common to all C40 carotenogenic organisms, constitutes the first reaction unique to the carotenoid branch of isoprenoid metabolism [16,29]. From this step, slightly different reactions can be found in different organisms. As examples of this diversity, it is worthy of highlighting that organisms like anoxygenic photosynthetic bacteria, non-photosynthetic bacteria, and fungi desaturate phytoene either three or four times to yield neurosporene or lycopene, respectively. However, oxygenic photosynthetic organisms (cyanobacteria, algae, and higher plants) convert phytoene to lycopene via carotene in two distinct sets of reactions. At the level of neurosporene, lycopene, or β-carotene, the carotenoid biosynthesis pathways from different organisms branch to generate the significant diversity of carotenoids found in nature [16]. In photosynthetic organisms and tissues, the lipophilic carotenoid and bacteriochlorophyll (Bchl) or chlorophyll (Chl) pigment molecules associate non-covalently, but specifically, with integral membrane proteins. In non-photosynthetic organisms and tissues, carotenoids, often protein-bound, occur in cytoplasmic or cell wall membranes, oil droplets, crystals, and fibrils [16,30,31].
Thus, nine main groups of pathways related to carotenogenesis could be identified apart from other minor significant branches. Some of the main features of these nine groups are described following (Figure 1). At the level of farnesyl-PP, the diapocarotene biosynthesis pathway allows the synthesis of several compounds like diapophytoene, diapolycopene, or diaponeurosporene [32,33,34]. In this pathway, the carotenoid pigments are synthesized mainly via the desaturation of squalene rather than the direct synthesis of dehydrosqualene through the non-reductive condensation of prenyl diphosphate precursors, indicating the possible existence of a “squalene route” and a “lycopersene route” for C30 and C40 carotenoids, respectively [35]. In some bacteria, the following genes coding for the enzymes catalyzing this pathway have been described: 4,4’-diapophytoene synthase (crtM), 4,4’-diapophytoene desaturase (crtNa), 4,4’-diapolycopene ketolase (crtNb) and 4,4’-diapolycopene aldehyde oxidase (crtNc) [36].
From neurosporene (Figure 1), the main pathway can continue to lycopene, or two more different branches can be distinguished: one of them for the synthesis of zeacarotene and the other one the spheroidene biosynthesis pathway [37]. The compounds made through the spheroidene biosynthesis pathway are mainly related to antenna complexes of sulfur photosynthetic bacteria [38,39].
Lycopene is one of the key molecules in the global carotenogenesis because it is the precursor for several branches sustaining carotenogenesis. Thus, relevant carotenoids in nature like lutein and its precursors and derivatives [40,41,42], neurosporaxanthin, and the C50-carotenoid called bacterioruberin [43] are synthesized from lycopene. Neurosporaxanthin is a carboxylic xanthophyll synthesized from geranylgeranyl pyrophosphate through the activity of four enzymes, encoded by the genes carRA, carB, carT, and carD [44]. It is mainly produced by fungi, being Fusarium and Neurospora, the genera from which its synthesis has been characterized [45,46]. Bacterioruberin is the main carotenoid synthesized by microorganisms belonging to the haloarchaea group, and only a few species of Micrococcus genus apart from haloarchaea can produce it [43,47]. The following sections are focused on the elucidation of the potential pathways that may be involved in the synthesis of bacterioruberin in haloarchaea. Finally, lycopene is the precursor for the synthesis of the carotenoids of the rhodophinal series (Figure 1, central panels) [48].
Lycopene can also be converted to γ-carotene. This molecule acts as a precursor for the synthesis of myxol and its derivatives, γ-carotene glucosides, or β-carotene. Myxol biosynthesis has been less explored than other pathways related to carotenogenesis, and most of the results reported came from studies on cyanobacteria [49,50,51]. γ-carotene glucosides are poorly described in the literature, and they are usually described as “rare” carotenoids [34,52].
γ-carotene can be converted to β-carotene, which is also one of the key molecules in the global carotenogenesis because it is the precursor for the synthesis of astaxanthin, xanthophylls, and retinol. Retinol metabolism has not been included in Figure 1 due to its complexity and its connections to other pathways like those related to lipids, bile acids, and vitamins [53,54,55]. The xanthophyll cycle represents one of the important photoprotection mechanisms in plant cells [56] and it has been also related to structural stabilization of membranes and to modulation of protein-membrane function [57,58]. Astaxanthin is one of the carotenoids being successfully produced at large scale using Haematococcus pluvialis [59,60]. This carotenoid is a potent lipid-soluble keto-carotenoid with auspicious effects on human health. It protects organisms against a wide range of diseases with excellent safety and tolerability [61,62].
Finally, it is important to highlight that recently, a new carotenoid-derived pathway has been described as responsible for the synthesis of apocarotenoids, which include isoprenoids with important functions in plant-environment interactions such as the attraction of pollinators and the defense against pathogens and herbivores. Apocarotenoids also include volatile aromatic compounds that act as repellents, chemoattractants, growth stimulators, and inhibitors, as well as the phytohormones abscisic acid and strigolactones [63,64,65].
2.2. A Search of the Genes for Carotenogenesis in Haloarchaeal Genomes
Considering all the information obtained for the reconstruction of the metabolic map for global carotenogenesis as well as the results already reported on the synthesis of carotenoids in haloarchaea, a two-step strategy was developed:
- (i)
- Genes involved in the synthesis of bacterioruberin in Haloarcula japonica DSM 6131 were used as a query to look for their homologs in haloarchaeal available genomes at NCBI genome database [12]. Special interest has been paid on the following genomes because these are the haloarchaeal species better described from a biochemical point of view at the time of writing this work: Haloferax mediterranei ATCC3500, Haloferax volcanii DS2, Haloferax gibbonsii ARA6, Haloarcula marismortui ATCC43049, Haloarcula hispanica N601, Halorubrum trapanicum CBA1232, Halorubrum ezzemoulense Fb21, Natronobacterium gregoryi SP2, Natronomonas moolapensis 8.8.11 and Haloquadratum walsbyi C23.
- (ii)
- All these sequences identified in the previous step were used as a query in BlastN to look for their homologs in other haloarchaeal genomes available at NCBI, in a one-to-one comparison. The enzymes encoded by these genes were identified using BlastX and Uniprot. Data about genes homology and identity are summarized in tables displayed in Supplementary Materials (for more details, see Section 4.2) (Supplementary materials: Tables S2–S14).
The results from this in silico analysis (Figure 2) demonstrated that in H. mediterranei genome, genes homologous to those involved in most of the carotenogenesis pathways were not identified (Figure 1). Haloarchaea synthesize terpenoid backbones through the mevalonate pathway, leading to the main branch of C40 carotenoids, common to the rest of organisms. At the lycopene level, the bacterioruberin synthesis pathway allows the synthesis of this C50 carotenoid and its derivatives. All of the studied haloarchaea share the genes involved in bacterioruberin synthesis, which were firstly described in Haloarcula japonica [12]. Thus, the pathway summarized in Figure 2 seems to be the main pathway sustaining the synthesis of these kinds of pigments in haloarchaea. Apart from bacterioruberin, haloarchaea could also be able to synthesize β-zeacaroten and 7,8-dihydro-β-carotene, using a minor branch from neurosporene (Figure 2, center).
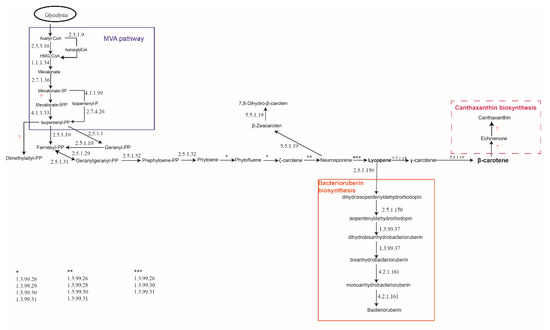
Figure 2.
Carotenogenesis predicted in haloarchaea based on in silico analysis.
Given this information, haloarchaea would synthesize almost exclusively bacterioruberin and its derivatives. However, few previous works have demonstrated that other carotenoids like canthaxanthin can be produced by Haloferax alexandrinus or Halobacterium species [66,67,68]. Nevertheless, the genes and enzymes involved in the synthesis of canthaxanthin were not clearly identified in neither of these species during this bioinformatic analysis. Another possible explanation would be the existence of undescribed pathways for the synthesis of other carotenoids, which would justify the allusion to them in some bibliography [68,69]. However, to this day, no other pigments have been clearly identified in haloarchaea.
2.3. Organization of the Genomes around the Genes Coding for the Enzymes Involved in the Synthesis of Bacterioruberin
The organization of ORFs around the genes coding for enzymes involved in the synthesis of bacterioruberin was examined in both, fully sequenced and drafts of haloarchaeal genomes, from eleven species that are representative of the genera Haloarcula, Haloferax, Halorubrum, Haloquadratum, Natronobacterium, and Natronomonas. Although there is little evidence of conservation in this region of the genome, all the genomes analyzed have at least one copy of genes coding the following enzymes: phytoene synthase, phytoene desaturase, prenyltransferase and a hypothetical protein annotated as “carotenoid biosynthesis protein” (Figure 3). All studied haloarchaea conserve a three-gene cluster containing one copy of genes coding phytoene desaturase, prenyltransferase and the already mentioned ‘’carotenoid biosynthesis protein’’. Apart from that, most species analyzed present also one or more other genes coding phytoene desaturase, phytoene synthase and prenyltransferase. However, these genes are usually nothing similar to the ones in the cluster nor among them. The total number of genes encoding the same enzyme is also variable, as, for example, in the case of the genus Haloferax, in which Haloferax mediterranei shows two genes for each of these enzymes, while Haloferax volcanii shows three genes coding prenyltransferase and only one for phytoene desaturase. On the opposite, Haloferax gibbonsii has three genes coding phytoene desaturase and only one for phytoene synthase and prenyltransferase. Regarding the genus Haloarcula, they all have a very conserved number of their genes, with the exception of Haloarcula japonica, which has two genes coding a prenyltransferase. Regarding the genus Halorubrum, most of the species analyzed present three copies of genes coding phytoene desaturase and two for phytoene synthase. However, Halorubrum trapanicum has one copy of the prenyltransferase gene, while Halorubrum ezzemoulense has two. Natronobacterium gregoryi and Natronomonas moolapensis have three genes coding for prenyltransferase and three encoding phytoene desaturases, but they differ in the number of phytoene synthase genes since Natronobacterium has one while Natronomonas has two Haloquadratum walsbyi presents two genes coding each of the three enzymes.
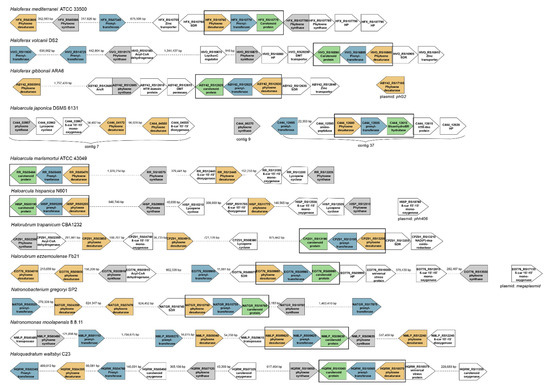
Figure 3.
Organization of the genome around the genes involved in carotenogenesis in selected haloarchaeal species. Colors have been used to highlight those genes well conserved between species. Green color represents the gene coding for carotenoid-like protein; blue color represents the gene coding for prenyltransferase; grey color highlights the gene encoding phytoene synthase, and yellow color represents the gene coding phytoene desaturase. The main conserved gene cluster is framed in black.
Some other genes coding enzymes related to oxidative processes in carotenogenesis (β,β-carotene-15′-15′-monooxygenase and β-carotene-15′-15′-dioxygenase) have been observed in the genera Haloarcula, Halorubrum, and Haloquadratum.
Given the fact that most species present have more than one gene coding the same enzyme, a comparative analysis of the homology of the genes included in the clusters and those copies located in other areas of the genome has been carried out. The data obtained are summarized in Tables S2–S14 included in the Supplementary Material. Since Haloferax mediterranei is our model organisms, the results of this comparison in Hfx. mediterranei are discussed into detail hereunder:
- (1)
- The analysis of the Hfx. mediterranei genome has revealed that this strain presents two genes encoding a phytoene desaturase: one inside the three-gene cluster and another one in another locus of the genome, both showing no resemblance between them. The one included in the three-gene cluster shows an identity between 68% and 87% with the correspondent gene of all the other species included in this study. On the other hand, when observing the gene located outside the cluster, it shares the highest homology (identity = 85.71%) with a phytoene desaturase-coding gene of Haloferax gibbonsii, also located outside the cluster; and shows also some identity with the correspondent gene of Natronobacterium gregoryi (67.13%) and with one of the two genes outside the cluster coding for this enzyme in Natronomonas moolapensis (65.67%), being both species haloalkalophiles.
- (2)
- Hfx. mediterranei genome also has two non-homolog genes coding prenyltransferase: one being part of the cluster and the other one positioned upstream in the genome. The one inside the cluster shows a homology between 69% and 82% with the correspondent gene of all the other studied species; while the one located out of the cluster is very similar to the out-of-cluster gene coding the same enzyme of Haloferax volcanii (84.22%) and also, to the one of Natronobacterium gregoryi (66.81%) and Natronomonas moolapensis (69.58%).
- (3)
- There is only one copy of the gene coding the unidentified ‘’carotenoid biosynthesis protein’’ in all the genomes studied. The one from Hfx. mediterranei genome shows a homology between 64% and 86% with the correspondent gene in the rest of the studied species, being the most similar Hfx. volcanii and Hfx. gibbonsii. However, the most remarkable hit is that this gene shows a 66.58% of homology with the gene coding the enzyme identified as bisanhydrobacterioruberin hydratase, also called C50 carotenoid 2′’,3′’-hydratase, in Haloarcula japonica [12].
- (4)
- Finally, as a significant feature of the Haloferax genus, it is worth mentioning the presence of a gene coding a zinc transporter next to the carotenogenesis three-gene cluster in all the species of this genus. The possible implication of this gene is still unknown.
The considerable diversity in the organization of genes around those ORFs coding for carotenoid protein, prenyltransferase, phytoene desaturase, and phytoene synthase, and the existence of more than one copy for these genes in some of the analyzed species could be accounted for by the recent acquisition of those genes through horizontal gene transfer and subsequent recombination events, as it has also been reported for other genes like those related to denitrification in haloarchaea [70]. In order to get a complete view of the evolutionary relationship of carotenogenesis in selected haloarchaeal species, a phylogenetic tree for each of the three genes included in the cluster has been elaborated and analyzed (Figure 4). For all the three genes, it can be observed that in general, each of the sequences is highly conserved between species belonging to the same genus. The exception to this general pattern is the prenyltransferase-coding gene and the unidentified carotenoid protein-coding gene from Halorubrum trapanicum, which are closer to the genus Haloarcula than to its own. Hfx. mediterranei gene sequences are close to those from another member of the Haloferax genus; however, it is interesting to highlight that for the three genes Hfx. mediterranei constitute a branch. The pattern of branching in the phylogenetic trees in the case of Hfx. mediterranei could reflect a slightly different evolution of carotenogenesis or different frequencies of events of recombination and or/gene transfer.

Figure 4.
Unrooted phylogenetic tree of genes encoding the following enzymes: (A) Prenyltransferase; (B) phytoene desaturase; (C) putative bisanhydrobacterioruberin hydratase. Colors have been used to group species belonging to the same genus. Tree scale indicates the phylogenetic distance in branches. Trees were built with Clustal Omega (default settings) and annotation was done with iTol v4.
3. Discussion
Since the first publication on carotenoids from archaea in the early sixties (last century) [71,72] more than 850 works have been published in indexed journals about this subject (PUBMED: data of access December the 10th, 2019; 856 publications identify using “carotenoids” and “archaea” as keywords). Most of these works addressed the characterization of the pigments synthesized by archaea, mainly haloarchaea, some others look for potential applications (due to their high antioxidant capability) and more recently, several works describe molecular engineering to optimize de production of some pigments in order to upscale their production [73,74,75,76]. However, there is still controversy about the nature of the pathways for carotenogenesis in haloarchaea, and consequently, the production of overpigmented mutants as cell factories for carotenoid production is still a limitation.
Considering that the number of haloarchaeal genomes fully sequenced has increased significantly during the last decade, and using plants, yeast, and cyanobacteria as model organisms from which carotenogenesis has been very well described so far, a three steps analysis has been conducted in order to elucidate key pathways involved in carotenogenesis in haloarchaea.
The first step was to reconstruct a metabolic map, including all the pathways described so far from a different types of organisms (involving Eukarya, Bacteria, and Archaea) (Figure 1). Secondly, the global map of carotenogenesis has been debugged through an in silico analysis to determine which pathways could be working in haloarchaea to sustain carotenogenesis. We analyzed around 100 genomes and, in particular, eleven species that are representative of the genera Haloarcula, Haloferax, Halorubrum, Haloterrigena, Haloquadratum, Natronobacterium and Natronomonas (Supplementary materials: Table S1). The results of the analysis suggest that although two components of the MVA pathway are often absent in archaeal genomes, the search for these missing elements led to the discovery of isopentenyl phosphate kinase (IPK) and phosphomevalonate decarboxylase (PMD). IPK and PMD carry out the two activities necessary to furnish the universal five-carbon isoprenoid building block, isopentenyl diphosphate (IPP) [77]. Thus, a functioning alternative MVA pathway could be the main pathway to provide isopentenyl-PP in haloarchaea from our results, which are in concordance with the results from other previous works [77,78,79].
Then, lycopene is obtained after a series of reactions in which geranylgeranyl-PP and prephytoene are produced. This main branch continues as a central metabolic of the backbone of carotenogenesis in which lycopene acts as a precursor (Figure 2). These results are in concordance with those described for Haloarcula japonica [12]. From lycopene, the major carotenoid produced by haloarchaea (bacterioruberin) is finally synthesized. Genes coding for the enzymes catalyzing reactions to produce β-carotene or β-zeacarotene have also been identified. So, on the base of nutritional status of the cells, and the physicochemical parameters fixed for cellular growth (temperature, radiation, pH, salt concentration, etc.), haloarchaea could synthetize phytoene, phytofluene, γ-carotene, β-carotene, neurosporene, lycopene, zeacarotene and bacterioruberin (and its derivatives and precursors) (Figure 2). The final concentration of each of those carotenoids, quantified from cellular extracts, will depend on the conditions of the culture as well as on the phase of growth in which the cells are harvested [80].
Some of the first studies on carotenoids in haloarchaea in the later seventies demonstrated that some haloarchaeal species could produce higher amounts of C40 carotenoids (lycopene and β-carotene) than C50 (bacterioruberin and its derivatives) when using glycerol as carbon source. Thus, the supplementation of the culture with up to 0.5% of glycerol stimulated carotenogenesis in Halobacterium cutirubrum, but the content of the major red pigment, bacterioruberin, was reduced fourfold and that of the minor red pigments, mono- and bisanhydrobacterioruberins, was also reduced but to a lesser extent [81]. Shortly after these discoveries, it was stated that the C50 bacterioruberin is made by the addition of a C5-isoprene unit to each end of the C40-lycopene chain, followed by the introduction of four hydroxyl groups [82].
In principle, all the genomes analyzed have at least one copy of genes coding the following enzymes: phytoene synthase, phytoene desaturase, prenyltransferase, and the hypothetical protein annotated as “carotenoid biosynthesis protein”. However, one of the main problems and limitations found during this study was the presence of ambiguous annotations regarding several genes. Hfx. mediterranei genome is an example of this limitation. Apparently, this species has two genes coding a phytoene desaturase, which are indeed nothing alike. One of these two genes, which is located outside the three-gene cluster, can be identified as a FAD-dependent oxidoreductase but is also annotated as an identical protein of phytoene desaturase. This conflict of annotation and the problems related to the incomplete or not completely accurate genome assembling have been previously described in other haloarchaeal studies [70]. Consequently, one of the main conclusions from this study is that more effort must be made in the next future to assembly haloarchaeal draft genomes and to review and update gene annotations in completely sequenced haloarchaeal genomes.
Regarding the unidentified protein, a similarity of the sequence has been found to the gene coding the bisanhydrobacterioruberin hydratase or C50 carotenoid 2′’,3′’-hydratase enzyme of Haloarcula japonica [12]. However, much experimental research, including mutants, is needed to confirm the putative function of this gene in Hfx. mediterranei.
The presence of a gene coding a zinc transporter is a peculiar feature only found in species of the Haloferax genus. This transporter could be related to the fact that prenyltransferases are usually metalloenzymes that use zinc to catalyze their reactions [83,84]. It is still unknown if the prenyltransferases from haloarchaea present a zinc-site recognition moiety. Nevertheless, from the results of our in silico analysis using the ab initio ligand predictor IonCom, it can be assumed that the two prenyltransferases encoded in the genome of Hfx. mediterranei possess potential zinc recognition sites [85]. Another potential connection between this zinc transporter and carotenogenesis could be the “bop-gene regulon [69,86,87,88]. bop gene is part of a cluster of genes, which is tightly regulated by a sensor regulator called bat and potentially also by a small zinc-finger containing a protein called Brz [69]. bop gene codes for bacterio-opsin protein: part of the light-driven proton pump called bacteriorhodopsin [60,88]. However, this potential connection would not be related to Hfx. mediterranei because this species is not able to synthesize bacteriorhodopsin [89].
The phylogenetic relationship of the sequences of the three genes included in the cluster shows that each of the sequences is highly conserved between species belonging to the same genus, with the exception of those from Halorubrum trapanicum. In this case, sequences were closer phylogenetically to the genus Haloarcula than to its own in two out of three of the cluster genes.
Finally, the presence of genes coding carotenoid oxygenases in Haloarcula, Halorubrum, and Haloquadratum could explain the more reddish pigmentation found in these genera when compared to other species such as Haloferax mediterranei, which show an intense pink color.
In summary, although haloarchaea could synthesize other carotenoids apart from bacterioruberin (like β-carotene, lycopene, or in some species canthaxanthin), the major carotenoid is bacterioruberin and its relatives. The pathway deciphered in this work (Figure 2) sheds light on the knowledge of carotenogenesis in archaea and will contribute to the production of mutants over pigmented in order to upscale carotenoids production using archaea as biofactories [80,90]. Other issues like regulation of carotenogenesis [91] in haloarchaea and connections between carotenogenesis, synthesis of bacteriorhodopsin, and monooxygenases [69] should be explored into detail in the near future in order to understand the whole process making possible cellular pigmentation in this kind of microorganisms.
4. Materials and Methods
4.1. Data Sample
To decipher pathways for carotenogenesis in haloarchaea, 100 haloarchaeal genomes (including fully sequenced genomes and drafts) available at NCB were analyzed (http://www.ncbi.nlm.nih.gov/genomes). Special interest was paid to the eleven following species because they are the best described from a physiological and biochemical point of view, up to the time of writing this work: Haloferax mediterranei ATCC3500, Haloferax volcanii DS2, Haloferax gibbonsii ARA6, Haloarcula japonica DSM6131, Haloarcula marismortui ATCC43049, Haloarcula hispanica N601, Halorubrum trapanicum CBA1232, Halorubrum ezzemoulense Fb21, Natronobacterium gregoryi SP2, Natronomonas moolapensis 8.8.11 and Haloquadratum walsbyi C23.
4.2. In Silico Analysis of the Haloarchaeal Genomes.
A three steps strategy was designed to carry out an in silico analysis: i) reconstruction of a metabolic map including all the pathways described so far from different type of organisms (Eukarya, Bacteria, and Archaea), by integrating all the information available at BRENDA (https://www.brenda-enzymes.org/index.php), KEGG (https://www.genome.jp/kegg/), MetaCyc (https://metacyc.org/), Uniprot (https://www.uniprot.org/) and NCBI (https://www-ncbi-nlm-nih-gov/biosystems); ii) prediction of pathways involved in carotenogenesis in haloarchaeal: genes already described as sequences involved in the synthesis of bacterioruberin in haloarchaea were used as a query to look for their homologs in selected haloarchaeal genomes at NCBI [12], and the organization of the genome around those genes was analyzed deeply; iii) search of homologs to gene sequences coding for the enzymes represented in Figure 1 in order to identify the main pathways sustaining carotenogenesis in haloarchaea.
Given the observed variability of sequences between the conserved cluster of different species, the errors found in the gene annotation, and the difficulty to identify the correct ORF in some cases, we did not establish a threshold to find orthologs. For this reason, in order to identify the putative orthologs in all studied species, each gene was compared to each other to determine the percentage of identity between them, and therefore, the conserved cluster (S2-14). In general, in this one-to-one comparison, a minimum query cover of 80% and a minimum identity of 60% was considered to identify the gene as an ortholog.
4.3. Bioinformatic Tools
BRENDA, KEGG, MetaCyc, and Uniprot databases were used to set up the metabolic map of carotenogenesis. NCBI Gene database was consulted to look for (i) genes previously related to bacterioruberin production; (ii) genes that encode the enzymes of the bacterioruberin pathway map in haloarchaeal genomes.
Blastn and BlastX online tools from NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi) were used respectively to find homologous genes and their proteins in haloarchaeal genomes [92]. The default setting and organism filter were selected. Gene location and genetic context studies were carried out by means of the bioinformatic tool NCBI Sequence Viewer 3.33.0 (https://www.ncbi.nlm.nih.gov/tools/sviewer/).
Alignments of the sequences and phylogenetic tree have been done using Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/) tool (web form) based on the HH algorithm described by Söding [93,94]. Default parameters were used. Annotation of the phylogenetic tree has been done with the online tool iTol v4 (https://itol.embl.de/) [95].
Zinc binding sites of Haloferax mediterranei prenyltransferases were analyzed using the IonCom server (https://zhanglab.ccmb.med.umich.edu/IonCom/) [85].
Supplementary Materials
The following are available online at https://www.mdpi.com/1420-3049/25/5/1197/s1. A total of 14 tables are available as Supplementary Materials.
Author Contributions
R.M.M.-E. conceived and designed the experiments, was the project administrator and oversaw the funding acquisition; M.G. and J.M.M.-R. conducted the bioinformatic analysis; M.G. designed all the conceptual figures; R.M.M.-E., C.P. and G.P. contributed to the analysis of the results. All the authors contributed equally to the writing of the original draft, review of the results and final editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially funded by research grants from MINECO Spain (RTI2018-099860-B-I00) and the University of Alicante (VIGROB-309).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mata-Gómez, L.; Montañez, J.; Méndez-Zavala, A.; Aguilar, C. Biotechnological production of carotenoids by yeasts: An overview. Microb. Cell Factories 2014, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Nelis, H.J.; De Leenheer, A.P. Microbial sources of carotenoid pigments used in foods and feeds. J. Appl. Bacteriol. 1991, 70, 181–191. [Google Scholar] [CrossRef]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. (Eds.) Carotenoids; Birkhäuser Verlag: Basel, Switzerland, 1995; ISBN 978-3-7643-2908-2. [Google Scholar]
- Sathasivam, R.; Ki, J.-S. A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, Z.; Sun, P.; Chen, T.; Chen, F. Microalgal carotenoids: Beneficial effects and potential in human health. Food Funct. 2014, 5, 413. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Heider, S.A.E.; Peters-Wendisch, P.; Wendisch, V.F.; Beekwilder, J.; Brautaset, T. Metabolic engineering for the microbial production of carotenoids and related products with a focus on the rare C50 carotenoids. Appl. Microbiol. Biotechnol. 2014, 98, 4355–4368. [Google Scholar] [CrossRef]
- Krubasik, P.; Kobayashi, M.; Sandmann, G. Expression and functional analysis of a gene cluster involved in the synthesis of decaprenoxanthin reveals the mechanisms for C50 carotenoid formation: Decaprenoxanthin formation. Eur. J. Biochem. 2001, 268, 3702–3708. [Google Scholar] [CrossRef]
- Norgård, S.; Aasen, A.J.; Liaaen-Jensen, S.; Holme, D.; Lamvik, A.; Sunde, E.; Sørensen, N.A. Bacterial Carotenoids. XXXII. C50-Carotenoids 6. Carotenoids from Corynebacterium poinsettiae Including Four New C50-Diols. Acta Chem. Scand. 1970, 24, 2183–2197. [Google Scholar] [CrossRef]
- Netzer, R.; Stafsnes, M.H.; Andreassen, T.; Goksoyr, A.; Bruheim, P.; Brautaset, T. Biosynthetic Pathway for -Cyclic Sarcinaxanthin in Micrococcus luteus: Heterologous Expression and Evidence for Diverse and Multiple Catalytic Functions of C50 Carotenoid Cyclases. J. Bacteriol. 2010, 192, 5688–5699. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids Database: Structures, chemical fingerprints and distribution among organisms. Database 2017, 2017. [Google Scholar] [CrossRef]
- Yang, Y.; Yatsunami, R.; Ando, A.; Miyoko, N.; Fukui, T.; Takaichi, S.; Nakamura, S. Complete Biosynthetic Pathway of the C 50 Carotenoid Bacterioruberin from Lycopene in the Extremely Halophilic Archaeon Haloarcula japonica. J. Bacteriol. 2015, 197, 1614–1623. [Google Scholar] [CrossRef]
- Rosas-Saavedra, C.; Stange, C. Biosynthesis of Carotenoids in Plants: Enzymes and Color. In Carotenoids in Nature; Stange, C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 79, pp. 35–69. ISBN 978-3-319-39124-3. [Google Scholar]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment: Carotenoids: Pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef]
- Henríquez, V.; Escobar, C.; Galarza, J.; Gimpel, J. Carotenoids in Microalgae. In Carotenoids in Nature; Stange, C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 79, pp. 219–237. ISBN 978-3-319-39124-3. [Google Scholar]
- Rodrigo-Baños, M.; Garbayo, I.; Vílchez, C.; Bonete, M.; Martínez-Espinosa, R. Carotenoids from Haloarchaea and Their Potential in Biotechnology. Mar. Drugs 2015, 13, 5508–5532. [Google Scholar] [CrossRef]
- Yin, J.; Chen, J.-C.; Wu, Q.; Chen, G.-Q. Halophiles, coming stars for industrial biotechnology. Biotechnol. Adv. 2015, 33, 1433–1442. [Google Scholar] [CrossRef]
- Abbes, M.; Baati, H.; Guermazi, S.; Messina, C.; Santulli, A.; Gharsallah, N.; Ammar, E. Biological properties of carotenoids extracted from Halobacterium halobium isolated from a Tunisian solar saltern. BMC Complementary Altern. Med. 2013, 13. [Google Scholar] [CrossRef]
- Gupta, R.S.; Naushad, S.; Baker, S. Phylogenomic analyses and molecular signatures for the class Halobacteria and its two major clades: A proposal for division of the class Halobacteria into an emended order Halobacteriales and two new orders, Haloferacales ord. nov. and Natrialbales ord. nov., containing the novel families Haloferacaceae fam. nov. and Natrialbaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 1050–1069. [Google Scholar]
- Falb, M.; Müller, K.; Königsmaier, L.; Oberwinkler, T.; Horn, P.; von Gronau, S.; Gonzalez, O.; Pfeiffer, F.; Bornberg-Bauer, E.; Oesterhelt, D. Metabolism of halophilic archaea. Extremophiles 2008, 12, 177–196. [Google Scholar] [CrossRef]
- Camacho-Córdova, D.I.; Camacho-Ruíz, R.M.; Córdova-López, J.A.; Cervantes-Martínez, J. Estimation of bacterioruberin by Raman spectroscopy during the growth of halophilic archaeon Haloarcula marismortui. Appl. Opt. 2014, 53, 7470. [Google Scholar] [CrossRef]
- Fang, C.-J.; Ku, K.-L.; Lee, M.-H.; Su, N.-W. Influence of nutritive factors on C50 carotenoids production by Haloferax mediterranei ATCC 33500 with two-stage cultivation. Bioresour. Technol. 2010, 101, 6487–6493. [Google Scholar] [CrossRef]
- Jehlička, J.; Edwards, H.G.M.; Oren, A. Bacterioruberin and salinixanthin carotenoids of extremely halophilic Archaea and Bacteria: A Raman spectroscopic study. Spectrochim. Acta Part A: Mol. Biomol. Spectrosc. 2013, 106, 99–103. [Google Scholar] [CrossRef]
- Mandelli, F.; Miranda, V.S.; Rodrigues, E.; Mercadante, A.Z. Identification of carotenoids with high antioxidant capacity produced by extremophile microorganisms. World J. Microbiol. Biotechnol. 2012, 28, 1781–1790. [Google Scholar] [CrossRef]
- Naziri, D.; Hamidi, M.; Hassanzadeh, S.; Tarhriz, V.; Maleki Zanjani, B.; Nazemyieh, H.; Hejazi, M.A.; Hejazi, M.S. Analysis of Carotenoid Production by Halorubrum sp. TBZ126; an Extremely Halophilic Archeon from Urmia Lake. Adv. Pharm. Bull. 2014, 4, 61–67. [Google Scholar]
- Matsumi, R.; Atomi, H.; Driessen, A.J.M.; van der Oost, J. Isoprenoid biosynthesis in Archaea – Biochemical and evolutionary implications. Res. Microbiol. 2011, 162, 39–52. [Google Scholar] [CrossRef]
- Sui, L.; Liu, L.; Deng, Y. Characterization of halophilic C50 carotenoid-producing archaea isolated from solar saltworks in Bohai Bay, China. Chin. J. Oceanol. Limnol. 2014, 32, 1280–1287. [Google Scholar] [CrossRef]
- Liao, P.; Hemmerlin, A.; Bach, T.J.; Chye, M.-L. The potential of the mevalonate pathway for enhanced isoprenoid production. Biotechnol. Adv. 2016, 34, 697–713. [Google Scholar] [CrossRef]
- Othman, R.; Mohd Zaifuddin, F.A.; Hassan, N.M. Carotenoid Biosynthesis Regulatory Mechanisms in Plants. J. Oleo Sci. 2014, 63, 753–760. [Google Scholar] [CrossRef]
- Arteni, A.-A.; LaFountain, A.M.; Alexandre, M.T.A.; Fradot, M.; Mendes-Pinto, M.M.; Sahel, J.-A.; Picaud, S.; Frank, H.A.; Robert, B.; Pascal, A.A. Carotenoid composition and conformation in retinal oil droplets of the domestic chicken. PLoS ONE 2019, 14, e0217418. [Google Scholar] [CrossRef]
- Li, L.; Yuan, H.; Zeng, Y.; Xu, Q. Plastids and Carotenoid Accumulation. In Carotenoids in Nature; Stange, C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 79, pp. 273–293. ISBN 978-3-319-39124-3. [Google Scholar]
- Hartz, P.; Milhim, M.; Trenkamp, S.; Bernhardt, R.; Hannemann, F. Characterization and engineering of a carotenoid biosynthesis operon from Bacillus megaterium. Metab. Eng. 2018, 49, 47–58. [Google Scholar] [CrossRef]
- Raisig, A.; Sandmann, G. Functional properties of diapophytoene and related desaturases of C30 and C40 carotenoid biosynthetic pathways. Biochim. Et Biophys. Acta (Bba) - Mol. Cell Biol. Lipids 2001, 1533, 164–170. [Google Scholar] [CrossRef]
- Takaichi, S.; Inoue, K.; Akaike, M.; Kobayashi, M.; Oh-oka, H.; Madigan, M.T. The major carotenoid in all known species of heliobacteria is the C30 carotenoid 4,4′-diaponeurosporene, not neurosporene. Arch. Microbiol. 1997, 168, 277–281. [Google Scholar] [CrossRef]
- Furubayashi, M.; Li, L.; Katabami, A.; Saito, K.; Umeno, D. Construction of carotenoid biosynthetic pathways using squalene synthase. Febs Lett. 2014, 588, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Steiger, S.; Perez-Fons, L.; Cutting, S.M.; Fraser, P.D.; Sandmann, G. Annotation and functional assignment of the genes for the C30 carotenoid pathways from the genomes of two bacteria: Bacillus indicus and Bacillus firmus. Microbiology 2015, 161, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Jirsakova, V.; Reiss-Husson, F. A specific carotenoid is required for reconstitution of the Rubrivivax gelatinosus B875 light harvesting complex from its subunit form B820. FEBS Lett. 1994, 353, 151–154. [Google Scholar] [CrossRef]
- Ashikhmin, A.A.; Makhneva, Z.K.; Bolshakov, M.A.; Shastik, E.S.; Moskalenko, A.A. Embedding carotenoids of spheroidene-branch biosynthesis into antenna complexes of sulfur photosynthetic bacteria. Dokl. Biochem. Biophys. 2016, 468, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Ashikhmin, A.; Makhneva, Z.; Bolshakov, M.; Moskalenko, A. Incorporation of spheroidene and spheroidenone into light-harvesting complexes from purple sulfur bacteria. J. Photochem. Photobiol. B Biol. 2017, 170, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-H.; Lee, D.-J.; Chang, J.-S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef]
- Sun, Z.; Li, T.; Zhou, Z.; Jiang, Y. Microalgae as a Source of Lutein: Chemistry, Biosynthesis, and Carotenogenesis. In Microalgae Biotechnology; Posten, C., Feng Chen, S., Eds.; Springer International Publishing: Cham, Switzerland, 2015; Volume 153, pp. 37–58. ISBN 978-3-319-23807-4. [Google Scholar]
- Van den Berg, H. Effect of lutein on beta-carotene absorption and cleavage. Int. J. Vitam Nutr. Res. 1998, 68, 360–365. [Google Scholar]
- Giani, M.; Garbayo, I.; Vílchez, C.; Martínez-Espinosa, R.M. Haloarchaeal Carotenoids: Healthy Novel Compounds from Extreme Environments. Mar. Drugs 2019, 17, 524. [Google Scholar] [CrossRef]
- Avalos, J.; Pardo-Medina, J.; Parra-Rivero, O.; Ruger-Herreros, M.; Rodríguez-Ortiz, R.; Hornero-Méndez, D.; Limón, M. Carotenoid Biosynthesis in Fusarium. J. Fungi 2017, 3, 39. [Google Scholar] [CrossRef]
- Hornero-Méndez, D.; Limón, M.C.; Avalos, J. HPLC Analysis of Carotenoids in Neurosporaxanthin-Producing Fungi. In Microbial Carotenoids; Barreiro, C., Barredo, J.-L., Eds.; Springer: New York, NY, USA, 2018; Volume 1852, pp. 269–281. ISBN 978-1-4939-8741-2. [Google Scholar]
- Avalos, J.; Prado-Cabrero, A.; Estrada, A.F. Neurosporaxanthin Production by Neurospora and Fusarium. In Microbial Carotenoids from Fungi; Barredo, J.-L., Ed.; Humana Press: Totowa, NJ, USA, 2012; Volume 898, pp. 263–274. ISBN 978-1-61779-917-4. [Google Scholar]
- Torregrosa-Crespo, J.; Montero, Z.; Fuentes, J.; Reig García-Galbis, M.; Garbayo, I.; Vílchez, C.; Martínez-Espinosa, R. Exploring the Valuable Carotenoids for the Large-Scale Production by Marine Microorganisms. Mar. Drugs 2018, 16, 203. [Google Scholar] [CrossRef]
- Francis, G.W.; Liaaen-Jensen, S.; Silvennoinen, K.; Vaahtera, K.; Shimizu, A. Bacterial Carotenoids. XXXIII. Carotenoids of Thiorhodaceae. 9. The Structures of the Carotenoids of the Rhodopinal Series. Acta Chemica Scandinavica 1970, 24, 2705–2712. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Teramoto, M.; Onodera, K.; Moriyama, H.; Komatsu, A.; Akakabe, M.; Nishijima, M. Aurantiacicella marina gen. nov., sp. nov., a myxol-producing bacterium from surface seawater. Int. J. Syst. Evol. Microbiol. 2016, 66, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Bryant, D.A. The Biosynthetic Pathway for Myxol-2′ Fucoside (Myxoxanthophyll) in the Cyanobacterium Synechococcus sp. Strain PCC 7002. J. Bacteriol. 2009, 191, 3292–3300. [Google Scholar] [CrossRef]
- Mochimaru, M.; Masukawa, H.; Maoka, T.; Mohamed, H.E.; Vermaas, W.F.J.; Takaichi, S. Substrate Specificities and Availability of Fucosyltransferase and -Carotene Hydroxylase for Myxol 2′-Fucoside Synthesis in Anabaena sp. Strain PCC 7120 Compared with Synechocystis sp. Strain PCC 6803. J. Bacteriol. 2008, 190, 6726–6733. [Google Scholar] [CrossRef]
- Takaichi, S.; Maoka, T.; Akimoto, N.; Carmona, M.L.; Yamaoka, Y. Carotenoids in a Corynebacterineae, Gordonia terrae AIST-1: Carotenoid Glucosyl Mycoloyl Esters. Biosci. Biotechnol. Biochem. 2008, 72, 2615–2622. [Google Scholar] [CrossRef] [PubMed]
- Villarroya, F.; Giralt, M.; Iglesias, R. Retinoids and adipose tissues: Metabolism, cell differentiation and gene expression. Int. J. Obes. 1999, 23, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.; Hoekstra, M.; Hoeke, M.O.; Heegsma, J.; Faber, K.N. The interrelationship between bile acid and vitamin A homeostasis. BBA-Mol. Cell Biol. L. 2017, 1862, 496–512. [Google Scholar] [CrossRef] [PubMed]
- Saari, J.C. Vitamin A and Vision. In The Biochemistry of Retinoid Signaling II.; Asson-Batres, M.A., Rochette-Egly, C., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2016; Volume 81, pp. 231–259. ISBN 978-94-024-0943-7. [Google Scholar]
- Goss, R.; Jakob, T. Regulation and function of xanthophyll cycle-dependent photoprotection in algae. Photosynth. Res. 2010, 106, 103–122. [Google Scholar] [CrossRef]
- Ruban, A.V.; Johnson, M.P. Xanthophylls as modulators of membrane protein function. Arch. Biochem. Biophys. 2010, 504, 78–85. [Google Scholar] [CrossRef]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Et Biophys. Acta (Bba)—Bioenerg. 2012, 1817, 182–193. [Google Scholar] [CrossRef]
- Brendler, T.; Williamson, E.M. Astaxanthin: How much is too much? A safety review. Phytother. Res. 2019, 33, 3090–3111. [Google Scholar] [CrossRef] [PubMed]
- Khoo, K.S.; Lee, S.Y.; Ooi, C.W.; Fu, X.; Miao, X.; Ling, T.C.; Show, P.L. Recent advances in biorefinery of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 2019, 288, 121606. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Astaxanthin: A mechanistic review on its biological activities and health benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Galasso, C.; Orefice, I.; Pellone, P.; Cirino, P.; Miele, R.; Ianora, A.; Brunet, C.; Sansone, C. On the Neuroprotective Role of Astaxanthin: New Perspectives? Mar. Drugs 2018, 16, 247. [Google Scholar] [CrossRef] [PubMed]
- Zwanenburg, B.; Blanco-Ania, D. Strigolactones: New plant hormones in the spotlight. J. Exp. Bot. 2018, 69, 2205–2218. [Google Scholar] [CrossRef] [PubMed]
- Beltran, J.C.M.; Stange, C. Apocarotenoids: A New Carotenoid-Derived Pathway. In Carotenoids in Nature; Stange, C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 79, pp. 239–272. ISBN 978-3-319-39124-3. [Google Scholar]
- Humphrey, A.; Beale, M. Strigol: Biogenesis and physiological activity. Phytochemistry 2006, 67, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Asker, D. Ohta Haloferax alexandrinus sp. nov., an extremely halophilic canthaxanthin-producing archaeon from a solar saltern in Alexandria (Egypt). Int. J. Syst. Evol. Microbiol. 2002, 52, 729–738. [Google Scholar]
- Asker, D.; Ohta, Y. Production of canthaxanthin by extremely halophilic bacteria. J. Biosci. Bioeng. 1999, 88, 617–621. [Google Scholar] [CrossRef]
- Calo, P.; Velazquez, J.B.; Sieiro, C.; Blanco, P.; Longo, E.; Villa, T.G. Analysis of astaxanthin and other carotenoids from several Phaffia rhodozyma mutants. J. Agric. Food Chem. 1995, 43, 1396–1399. [Google Scholar] [CrossRef]
- Müller, W.J.; Smit, M.S.; van Heerden, E.; Capes, M.D.; DasSarma, S. Complex Effects of Cytochrome P450 Monooxygenase on Purple Membrane and Bacterioruberin Production in an Extremely Halophilic Archaeon: Genetic, Phenotypic, and Transcriptomic Analyses. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Torregrosa-Crespo, J.; González-Torres, P.; Bautista, V.; Esclapez, J.M.; Pire, C.; Camacho, M.; Bonete, M.J.; Richardson, D.J.; Watmough, N.J.; Martínez-Espinosa, R.M. Analysis of multiple haloarchaeal genomes suggests that the quinone-dependent respiratory nitric oxide reductase is an important source of nitrous oxide in hypersaline environments: Analysis of multiple haloarchaeal genomes. Environ. Microbiol. Rep. 2017, 9, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.M. Carotenoid pigments of halophilic bacteria. Can. J. Microbiol. 1960, 6, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.L.; Brown, A.D. The membrane lipids of Halobacterium halobium. Biochem. J. 1968, 110, 441–448. [Google Scholar] [CrossRef]
- Calegari-Santos, R.; Diogo, R.A.; Fontana, J.D.; Bonfim, T.M.B. Carotenoid Production by Halophilic Archaea Under Different Culture Conditions. Curr. Microbiol. 2016, 72, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Chaari, M.; Theochari, I.; Papadimitriou, V.; Xenakis, A.; Ammar, E. Encapsulation of carotenoids extracted from halophilic Archaea in oil-in-water (O/W) micro- and nano-emulsions. Colloids Surf. B Biointerfaces 2018, 161, 219–227. [Google Scholar] [CrossRef]
- Hou, J.; Cui, H.-L. In Vitro Antioxidant, Antihemolytic, and Anticancer Activity of the Carotenoids from Halophilic Archaea. Curr. Microbiol. 2018, 75, 266–271. [Google Scholar] [CrossRef]
- Squillaci, G.; Parrella, R.; Carbone, V.; Minasi, P.; La Cara, F.; Morana, A. Carotenoids from the extreme halophilic archaeon Haloterrigena turkmenica: Identification and antioxidant activity. Extremophiles 2017, 21, 933–945. [Google Scholar] [CrossRef]
- Dellas, N.; Thomas, S.T.; Manning, G.; Noel, J.P. Discovery of a metabolic alternative to the classical mevalonate pathway. eLife 2013, 2. [Google Scholar] [CrossRef]
- Swift, I.E.; Milborrow, B.V. Stereochemistry of allene biosythesis and the formation of the acetylenic carotenoid diadinoxanthin and peridinin (C37) from neoxanthin. Biochem. J. 2005, 389, 919. [Google Scholar] [CrossRef]
- VanNice, J.C.; Skaff, D.A.; Keightley, A.; Addo, J.K.; Wyckoff, G.J.; Miziorko, H.M. Identification in Haloferax volcanii of Phosphomevalonate Decarboxylase and Isopentenyl Phosphate Kinase as Catalysts of the Terminal Enzyme Reactions in an Archaeal Alternate Mevalonate Pathway. J. Bacteriol. 2014, 196, 1055–1063. [Google Scholar] [CrossRef]
- Montero-Lobato, Z.; Ramos-Merchante, A.; Fuentes, J.; Sayago, A.; Fernández-Recamales, Á.; Martínez-Espinosa, R.; Vega, J.; Vílchez, C.; Garbayo, I. Optimization of Growth and Carotenoid Production by Haloferax mediterranei Using Response Surface Methodology. Mar. Drugs 2018, 16, 372. [Google Scholar] [CrossRef]
- Kushwaha, S.C.; Kates, M.K. Effect of glycerol on carotenogenesis in the extreme halophile, Halobacterium Cutirubrum. Can. J. Microbiol. 1979, 25, 1288–1291. [Google Scholar] [CrossRef]
- Kushwaha, S.C.; Kates, M. Studies of the biosynthesis of C50 carotenoids in Halobacterium Cutirubrum. Can. J. Microbiol. 1979, 25, 1292–1297. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Radwan, M.O.; Hamasaki, A.; Ejima, A.; Obata, E.; Koga, R.; Tateishi, H.; Okamoto, Y.; Fujita, M.; Nakao, M.; et al. A novel inhibitor of farnesyltransferase with a zinc site recognition moiety and a farnesyl group. Bioorganic Med. Chem. Lett. 2017, 27, 3862–3866. [Google Scholar] [CrossRef] [PubMed]
- Matthews, R.G.; Goulding, C.W. Enzyme-catalyzed methyl transfers to thiols: The role of zinc. Curr. Opin. Chem. Biol. 1997, 1, 332–339. [Google Scholar] [CrossRef]
- Hu, X.; Dong, Q.; Yang, J.; Zhang, Y. Recognizing metal and acid radical ion-binding sites by integrating ab initio modeling with template-based transferals. Bioinformatics 2016, 32, 3260–3269. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, V.Y.; Besir, H.; Schwaiger, R.; Klee, K.; Furtwängler, K.; Pfeiffer, F.; Oesterhelt, D. A small protein from the bop-brp intergenic region of Halobacterium salinarum contains a zinc finger motif and regulates bop and crtB1 transcription: Zinc finger regulator of bop and crtB1 expression. Mol. Microbiol. 2008, 67, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, V.; Schwaiger, R.; Furtwängler, K.; Dyall-Smith, M.; Oesterhelt, D. A small basic protein from the brz-brb operon is involved in regulation of bop transcription in Halobacterium salinarum. BMC Mol. Biol. 2011, 12, 42. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, P.; Zamora, R.C.; Muller, J.A.; DasSarma, S. Genome-Wide Responses of the Model Archaeon Halobacterium sp. Strain NRC-1 to Oxygen Limitation. J. Bacteriol. 2012, 194, 5530–5537. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Hallsworth, J.E. Microbial weeds in hypersaline habitats: The enigma of the weed-like Haloferax mediterranei. FEMS Microbiol. Lett. 2014, 359, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.Q.; Xue, Q.; Zhou, J.; Zhao, D.H.; Han, J.; Xiang, H. Engineering Haloferax mediterranei as an Efficient Platform for High Level Production of Lycopene. Front. Microbiol. 2018, 29, 2893. [Google Scholar] [CrossRef] [PubMed]
- Cerletti, N.; Paggi, R.; Troetschel, C.; Ferrari, M.C.; Ramallo Guevara, C.; Albaum, S.; Poetsch, A.; De Castro, R. LonB Protease is a novel regulator of carotenogenesis controlling degradation of phytoene synthase in Haloferax volcanii. J. Proteome Res. 2018, 17, 1158–1171. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2014, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Soding, J. Protein homology detection by HMM-HMM comparison. Bioinformatics 2005, 21, 951–960. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
Sample Availability: This study is based on bioinformatics analysis. Consequently, details about sample availability are not required. |




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).