Differentiation of South African Game Meat Using Near-Infrared (NIR) Spectroscopy and Hierarchical Modelling

 , , , ,
, , , ,  and
and
Abstract
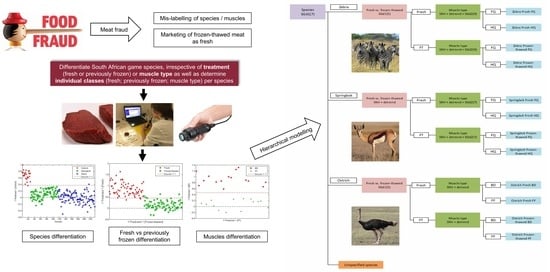
:
1. Introduction
2. Results and Discussion
2.1. Species Determination
2.1.1. Principal Component Analysis
2.1.2. Partial Least Squares Discriminant Analysis (PLS-DA)
2.2. Fresh vs. Previously Frozen Meat Determination
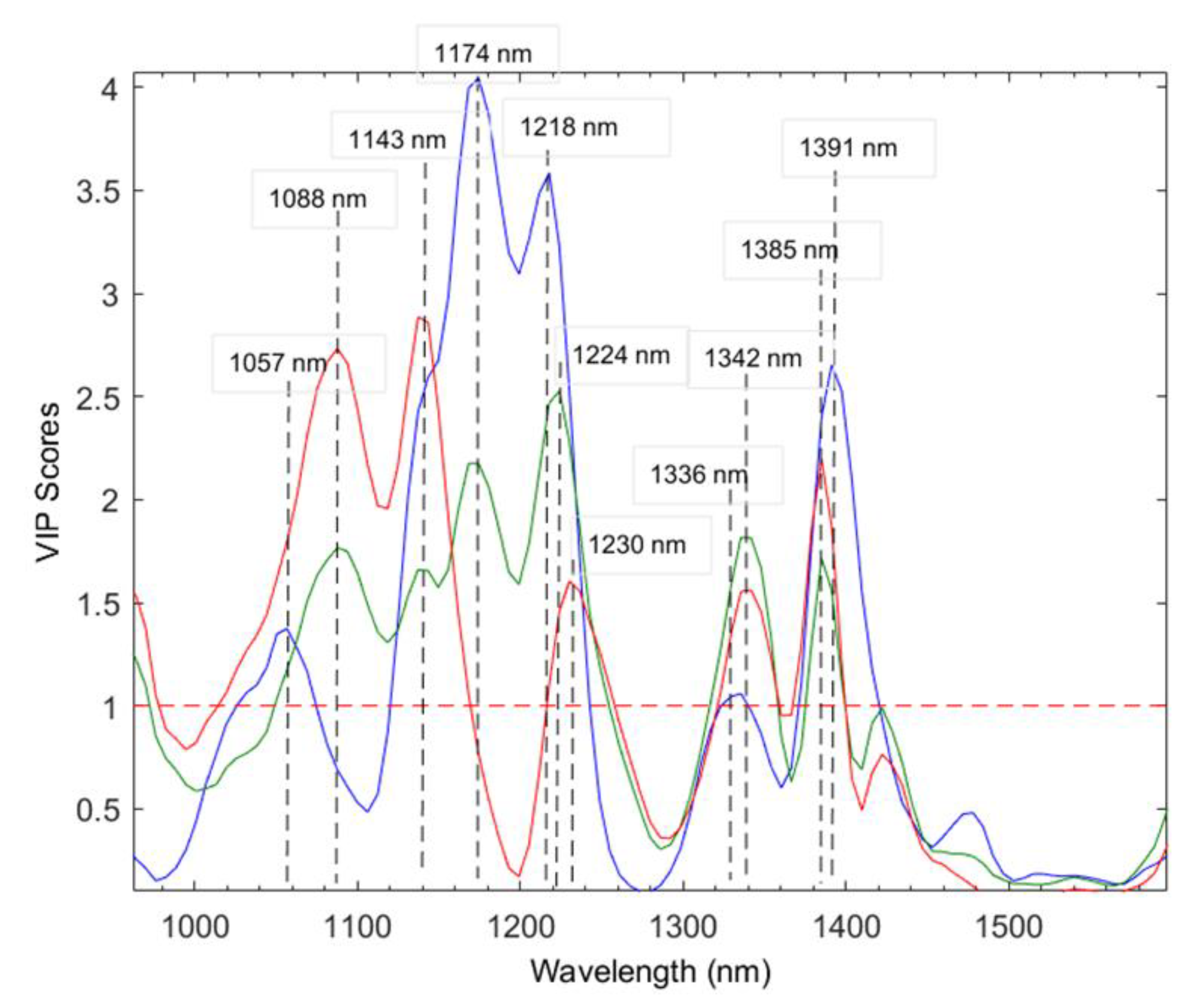
2.2.1. Spectral Analysis
2.2.2. Principal Component Analysis
2.2.3. Partial Least Squares Discriminant Analysis
2.3. Muscle Type Determination
2.3.1. Principal Component Analysis
2.3.2. Partial Least Squares Discriminant Analysis
2.4. Hierarchical Model Validation
3. Materials and Methods
3.1. Samples, Sampling, and Sample Preparation
3.2. NIR Instrumentation and Acquisition
3.3. Data Analysis
3.3.1. Spectral Analysis
3.3.2. Exploratory Data Analysis
3.3.3. Multivariate Data Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Steenkamp, J.-B.E.M. Dynamics in consumer behavior with respect to agricultural and food products. In Agricultural Marketing and Consumer Behavior in a Changing World; Kluwer Academic Publishers: Boston, MA, USA, 1997; pp. 143–188. [Google Scholar]
- Hoffman, L.C.; Bigalke, R.C. Utilising wild ungulates from southern Africa for meat production: Potential research requirements for the new millennium. In Proceedings of the 37th Congress of the Wildlife Management Association of South Africa, Pretoria, South Africa, 21–25 September 1999; pp. 20–21. [Google Scholar]
- D’Amato, M.E.; Alechine, E.; Cloete, K.W.; Davison, S.; Corach, D. Where is the game? Wild meat products authentication in South Africa: A case study. Invest. Gen. 2013, 4, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, L.C.; Muller, M.; Schutte, D.W.; Crafford, K. The retail of South African game meat: Current trade and marketing trends. S. Afr. J. Wildl. Res. 2004, 34, 123–134. [Google Scholar]
- Hoffman, L.C. The effect of different culling methodologies on the physical meat quality attributes of various game species. In Proceedings of the 5th International Wildlife Ranching Symposium, Pretoria, South Africa, 12–24 March 2001; pp. 212–221. [Google Scholar]
- Ballin, N.Z. Authentication of meat and meat products. Meat Sci. 2010, 86, 577–587. [Google Scholar] [CrossRef]
- Cawthorn, D.-M.; Steinman, H.A.; Hoffman, L.C. A high incidence of species substitution and mislabelling detected in meat products sold in South Africa. Food Control. 2013, 32, 440–449. [Google Scholar] [CrossRef] [Green Version]
- Alamprese, C.; Amigo, J.M.; Casiraghi, E.; Engelsen, S.B. Identification and quantification of turkey meat adulteration in fresh, frozen-thawed and cooked minced beef by FT-NIR spectroscopy and chemometrics. Meat Sci. 2016, 121, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Dean, N.; Murphy, T.B.; Downey, G. Using unlabelled data to update classification rules with applications in food authenticity studies. J. R. Stat. Soc. Appl. Stat. 2006, 55, 1–14. [Google Scholar] [CrossRef]
- Manley, M.; Baeten, V. Spectroscopic Technique: Near Infrared (NIR) Spectroscopy. In Modern Techniques for Food Authentication; Sun, D.-W., Ed.; Elsevier: Oxford, UK, 2018; pp. 51–102. [Google Scholar]
- Manley, M. Near-infrared spectroscopy and hyperspectral imaging: Non-destructive analysis of biological materials. Chem. Soc. Rev. 2014, 43, 8200–8214. [Google Scholar] [CrossRef] [Green Version]
- Prevolnik, M.; Čandek-Potokar, M.; Škorjanc, D. Predicting pork water-holding capacity with NIR spectroscopy in relation to different reference methods. J. Food Eng. 2010, 98, 347–352. [Google Scholar] [CrossRef]
- ElMasry, G.; Sun, D.-W.; Allen, P. Near-infrared hyperspectral imaging for predicting colour, pH and tenderness of fresh beef. J. Food Eng. 2012, 110, 127–140. [Google Scholar] [CrossRef]
- Kamruzzaman, M.; ElMasry, G.; Sun, D.-W.; Allen, P. Non-destructive assessment of instrumental and sensory tenderness of lamb meat using NIR hyperspectral imaging. Food Chem. 2013, 141, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Prieto, N.; Roehe, R.; Lavin, P.; Batten, G.; Andres, S. Application of near infrared reflectance spectroscopy to predict meat and meat products quality: A review. Meat Sci. 2009, 83, 175–186. [Google Scholar] [CrossRef]
- Alomar, D.; Gallo, C.; Castaneda, M.; Fuchslocher, R. Chemical and discriminant analysis of bovine meat by near infrared reflectance spectroscopy (NIRS). Meat Sci. 2003, 63, 441–450. [Google Scholar] [CrossRef]
- Downey, G.; Beauchêne, D. Discrimination between fresh and frozen-then-thawed beef m. longissimus dorsi by combined visible-near infrared reflectance spectroscopy: A feasibility study. Meat Sci. 1997, 45, 353–363. [Google Scholar] [CrossRef]
- Thyholt, K.; Isaksson, T. Differentiation of frozen and unfrozen beef using near-infrared spectroscopy. J. Sci. Food Agri. 1997, 73, 525–532. [Google Scholar] [CrossRef]
- Ropodi, A.I.; Panagou, E.Z.; Nychas, G.J.E. Rapid detection of frozen-then-thawed minced beef using multispectral imaging and Fourier transform infrared spectroscopy. Meat Sci. 2018, 135, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Xu, R. Differentiation of Beef and Kangaroo Meat by Visible/Near-Infrared Reflectance Spectroscopy. J. Food Sci. 1999, 64, 814–817. [Google Scholar] [CrossRef]
- Cozzolino, D.; Murray, I. Identification of animal meat muscles by visible and near infrared reflectance spectroscopy. LWT-Food Sci. Tech. 2004, 37, 447–452. [Google Scholar] [CrossRef]
- Schmutzler, M.; Beganovic, A.; Böhler, G.; Huck, C.W. Methods for detection of pork adulteration in veal product based on FT-NIR spectroscopy for laboratory, industrial and on-site analysis. Food Control 2015, 57, 258–267. [Google Scholar] [CrossRef]
- Mitsumoto, M.; Maeda, S.; Mitsuhashi, T.; Ozawa, S. Near-infrared spectroscopy determination of physical and chemical characteristics in beef cuts. J. Food Sci. 1991, 56, 1493–1496. [Google Scholar] [CrossRef]
- Osborne, B.G.; Fearn, T.; Hindle, P.H. Practical NIR Spectroscopy with Applications in Food and Beverage Analysis; Longman Scientific and Technical: Essex, UK, 1993; ISBN 0582099463. [Google Scholar]
- Esbensen, K.H.; Guyot, D.; Westad, F.; Houmoller, L.P. Multivariate Data Analysis in Practice: An. Introduction to Multivariate Data Analysis and Experimental Design; CAMO Process AS: Oslo, Norway, 2002; p. 598. ISBN 8299333032. [Google Scholar]
- Miller, C.E. Chemometrics in Process Analytical Chemistry. In Process Analytical Technology: Spectroscopic Tools and Implementation Strategies for the Chemical and Pharmaceutical Industries; Bakeev, K.A., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2005; pp. 226–324. ISBN 978-1-4051-2103-3. [Google Scholar]
- Barnes, R.; Dhanoa, M.; Lister, S.J. Standard normal variate transformation and de-trending of near-infrared diffuse reflectance spectra. Appl. Spectrosc. 1989, 43, 772–777. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- McElhinney, J.; Downey, G.; Fearn, T. Chemometric processing of visible and near infrared reflectance spectra for species identification in selected raw homogenised meats. J. Near Infrared Spectrosc. 1999, 7, 145–154. [Google Scholar] [CrossRef]
- Mamani-Linares, L.; Gallo, C.; Alomar, D. Identification of cattle, llama and horse meat by near infrared reflectance or transflectance spectroscopy. Meat Sci. 2012, 90, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Farrés, M.; Platikanov, S.; Tsakovski, S.; Tauler, R. Comparison of the variable importance in projection (VIP) and of the selectivity ratio (SR) methods for variable selection and interpretation. J. Chemometr. 2015, 29, 528–536. [Google Scholar] [CrossRef]
- Kamruzzaman, M.; ElMasry, G.; Sun, D.-W.; Allen, P. Non-destructive prediction and visualization of chemical composition in lamb meat using NIR hyperspectral imaging and multivariate regression. Innov. Food Sci. Emerg. Technol. 2012, 16, 218–226. [Google Scholar] [CrossRef]
- ElMasry, G.; Nakauchi, S. Prediction of meat spectral patterns based on optical properties and concentrations of the major constituents. Food Sci. Nutr. 2016, 4, 269–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbin, D.F.; Sun, D.-W.; Su, C. NIR hyperspectral imaging as non-destructive evaluation tool for the recognition of fresh and frozen–thawed porcine longissimus dorsi muscles. Innov. Food Sci. Emerg. Technol. 2013, 18, 226–236. [Google Scholar] [CrossRef]
- Leygonie, C.; Britz, T.J.; Hoffman, L.C. Meat quality comparison between fresh and frozen/thawed ostrich M. iliofibularis. Meat Sci. 2012, 91, 364–368. [Google Scholar] [CrossRef]
- Leygonie, C.; Britz, T.J.; Hoffman, L.C. Impact of freezing and thawing on the quality of meat: Review. Meat Sci. 2012, 91, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, R.A.; Ledward, D.A. Lawrie’s Meat Science, 7th ed.; Woodhead Publishing Limited: Cambridge, UK, 2006; pp. 1–417. ISBN 978-1-84569-159-2. [Google Scholar]
- ElMasry, G.; Sun, D.-W.; Allen, P. Chemical-free assessment and mapping of major constituents in beef using hyperspectral imaging. J. Food Eng. 2013, 117, 235–246. [Google Scholar] [CrossRef]
- Barbin, D.F.; ElMasry, G.; Sun, D.-W.; Allen, P. Non-destructive determination of chemical composition in intact and minced pork using near-infrared hyperspectral imaging. Food Chem. 2013, 138, 1162–1171. [Google Scholar] [CrossRef] [PubMed]
- DAFF. Meat Safety Act (Act No. 119 of 1990, No. R. 342). In Department of Agriculture, Forestry and Fisheries (DAFF); Government Printer: Pretoria, South Africa, 2000. [Google Scholar]
- Leygonie, C.; Britz, T.J.; Hoffman, L.C. Visual colour development in the ostrich M. iliofibularis muscle. In Proceedings of the 15th World Congress of Food Science and Technology (IUFoST), Cape Town, South Africa, 22–26 August 2010; p. 23. [Google Scholar]
- Wold, S.; Esbensen, K.; Geladi, P. Principal component analysis. Chemometr. Intel. Lab. Syst. 1987, 2, 37–52. [Google Scholar] [CrossRef]
- Wold, S.; Sjöström, M. SIMCA: A method for analyzing chemical data in terms of similarity and analogy. In Chemometrics: Theory and Application; Kowalski, B.R., Ed.; ACS Publications: Washington, DC, USA, 1977; Volume 52, pp. 243–282. ISBN 978-08-4120-439-3. [Google Scholar]
- Sebestyen, G. Pattern recognition by an adaptive process of sample set construction. IRE Trans. Inform. Theory 1962, 8, 82–91. [Google Scholar] [CrossRef]
- Wold, S.; Geladi, P.; Esbensen, K.; Öhman, J. Multi-way principal components-and PLS-analysis. J. Chemometr. 1987, 1, 41–56. [Google Scholar] [CrossRef]
- Kennard, R.W.; Stone, L.A. Computer aided design of experiments. Technometrics 1969, 11, 137–148. [Google Scholar] [CrossRef]
Sample Availability: Spectral data are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Game Species | Pre-Processing | Calibration | Validation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No of LVs 1 | nT6 | nC7 | Classification Accuracy (%) | nC7 | CV 2 (%) | nT6 | nP8 | Prediction Accuracy (%) | ||
| Overall | SNV 3 + DT 4 | 8 | 233 | 223 | 95.7 | 220 | 94.4 | 120 | 106 | 88.3 |
| SGd1(7) 5 | 7 | 235 | 219 | 93.2 | 209 | 88.9 | 118 | 111 | 94.1 | |
| Zebra | SNV 3 + DT 4 | 8 | 120 | 118 | 98.2 | 118 | 97.4 | 72 | 70 | 92.2 |
| SGd1(7) 5 | 7 | 111 | 103 | 94.4 | 99 | 91.3 | 80 | 74 | 94.9 | |
| Springbok | SNV 3 + DT 4 | 8 | 81 | 74 | 95.7 | 72 | 94.4 | 43 | 31 | 88.3 |
| SGd1(7) 5 | 7 | 95 | 88 | 93.6 | 83 | 89.3 | 29 | 28 | 94.1 | |
| Ostrich | SNV 3 + DT 4 | 8 | 32 | 31 | 97.4 | 30 | 96.9 | 5 | 5 | 95.5 |
| SGd1(7) 5 | 7 | 29 | 28 | 98.2 | 27 | 96.8 | 9 | 9 | 99.1 | |
| Game Species | Pre-Processing | Calibration | Validation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No of LVs 1 | nT7 | nC8 | Classification Accuracy (%) | nC8 | CV 2 (%) | nT7 | nP9 | Prediction Accuracy (%) | ||
| Zebra | SNV 3 + DT 4 | 3 | 144 | 137 | 95.1 | 134 | 93.1 | 48 | 48 | 100 |
| SGd1(5) 5 | 5 | 144 | 143 | 99.3 | 139 | 96.5 | 48 | 48 | 100 | |
| Springbok | SNV 3 + DT 4 | 2 | 93 | 93 | 100 | 93 | 100 | 31 | 31 | 100 |
| SNV 3 + DT 4 + SGd2(7) 6 | 2 | 93 | 93 | 100 | 90 | 96.7 | 31 | 31 | 100 | |
| Ostrich | SNV 3 + DT 4 | 1 | 30 | 27 | 90 | 25 | 88.3 | 10 | 5 | 50 |
| SGd1(5) 5 | 3 | 30 | 27 | 90 | 26 | 86.7 | 10 | 9 | 90 | |
| Game Species | Pre-Processing | Calibration | Validation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No of LVs 1 | nT7 | nC8 | Classification Accuracy (%) | nC8 | CV 2 (%) | nT7 | nP9 | Prediction Accuracy (%) | ||
| Zebra | SNV 3 + DT 4 | 4 | 144 | 118 | 81.9 | 116 | 80.6 | 48 | 39 | 81.3 |
| SNV 3 + DT 4 + SGd2(9) 6 | 5 | 144 | 130 | 90.3 | 120 | 83.3 | 48 | 41 | 85.4 | |
| Springbok | SNV 3 + DT 4 | 3 | 93 | 78 | 83.9 | 75 | 80.7 | 31 | 19 | 61.3 |
| SNV 3 + DT 4 + SGd2(7) 5 | 5 | 93 | 91 | 97.9 | 83 | 89.3 | 31 | 30 | 96.8 | |
| Ostrich | SNV 3 + DT 4 | 5 | 30 | 30 | 100 | 30 | 100 | 10 | 10 | 100 |
| SGd2(7) 5 | 5 | 30 | 30 | 100 | 29 | 96.7 | 10 | 10 | 100 | |
| Data Pre-Treatment/LVs 1 | Correct Prediction | Data Pre-Treatment/LVs 1 | Correct Prediction | Data Pre-Treatment/LVs 1 | Correct Prediction | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species Classification | Fresh vs. Frozen-Thawed | Muscle Type | |||||||||
| SGd1(7)7/8 LVs1 | nT10 | nP11 | % | SGd1(5)6/5 LVs1 | nT10 | nP11 | % | SNV2 + DT 3 + SGd2(9) 5/6 LVs 1 | nT10 | nP11 | % |
| Zebra | 52 | 48 | 92.3 | Fresh Frozen-thawed | 21 27 | 21 27 | 100 100 | Forequarters Hindquarters | 21 27 | 17 24 | 80.9 88.9 |
| Springbok | nT10 | nP11 | % | SNV2 + DT 3/2 LVs 1 | nT10 | nP11 | % | SNV2 + DT 3 + SGd2(7) 4/5 LVs 1 | nT10 | nP11 | % |
| 32 | 31 | 96.9 | Fresh Frozen-thawed | 18 13 | 18 13 | 100 100 | Forequarters Hindquarters | 15 16 | 15 15 | 100 93.8 | |
| Ostrich | nT10 | nP11 | % | SGd1(5)6/3 LVs1 | nT10 | nP11 | % | SNV2 + DT 3/5 LVs 1 | nT10 | nP11 | % |
| 9 | 9 | 100 | Fresh Frozen-thawed | 5 4 | 4 4 | 80 100 | BD8 FF9 | 4 4 | 4 4 | 100 100 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edwards, K.; Manley, M.; Hoffman, L.C.; Beganovic, A.; Kirchler, C.G.; Huck, C.W.; Williams, P.J. Differentiation of South African Game Meat Using Near-Infrared (NIR) Spectroscopy and Hierarchical Modelling. Molecules 2020, 25, 1845. https://doi.org/10.3390/molecules25081845
Edwards K, Manley M, Hoffman LC, Beganovic A, Kirchler CG, Huck CW, Williams PJ. Differentiation of South African Game Meat Using Near-Infrared (NIR) Spectroscopy and Hierarchical Modelling. Molecules. 2020; 25(8):1845. https://doi.org/10.3390/molecules25081845
Chicago/Turabian StyleEdwards, Kiah, Marena Manley, Louwrens C. Hoffman, Anel Beganovic, Christian G. Kirchler, Christian W. Huck, and Paul J. Williams. 2020. "Differentiation of South African Game Meat Using Near-Infrared (NIR) Spectroscopy and Hierarchical Modelling" Molecules 25, no. 8: 1845. https://doi.org/10.3390/molecules25081845