
Apigenin Inhibits Histamine-Induced Cervical Cancer Tumor Growth by Regulating Estrogen Receptor Expression


Abstract
:
1. Introduction
2. Results
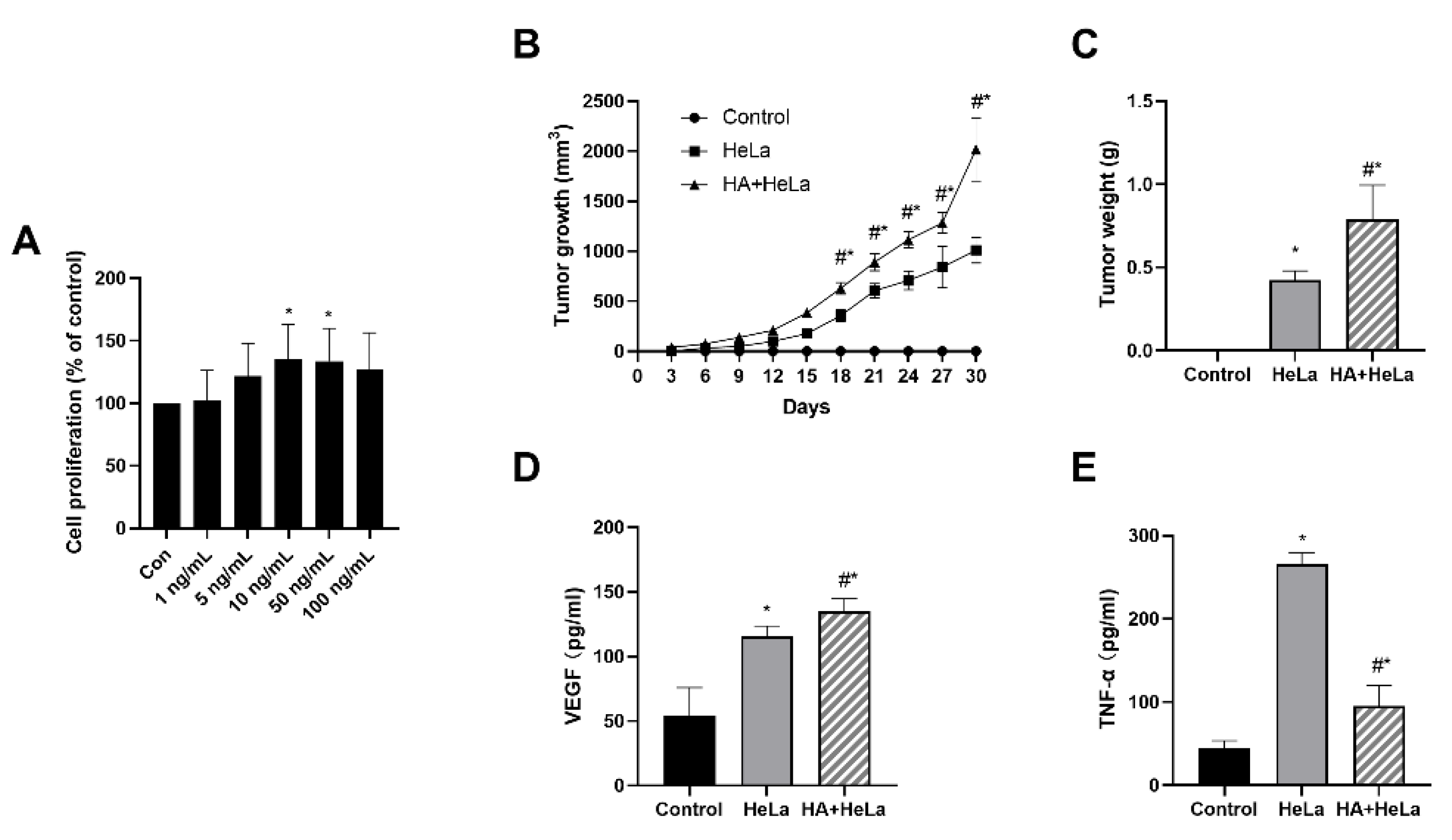
2.1. Histamine Treatment Stimulate Cervical Cancer Tumor Growth In Vivo and In Vitro
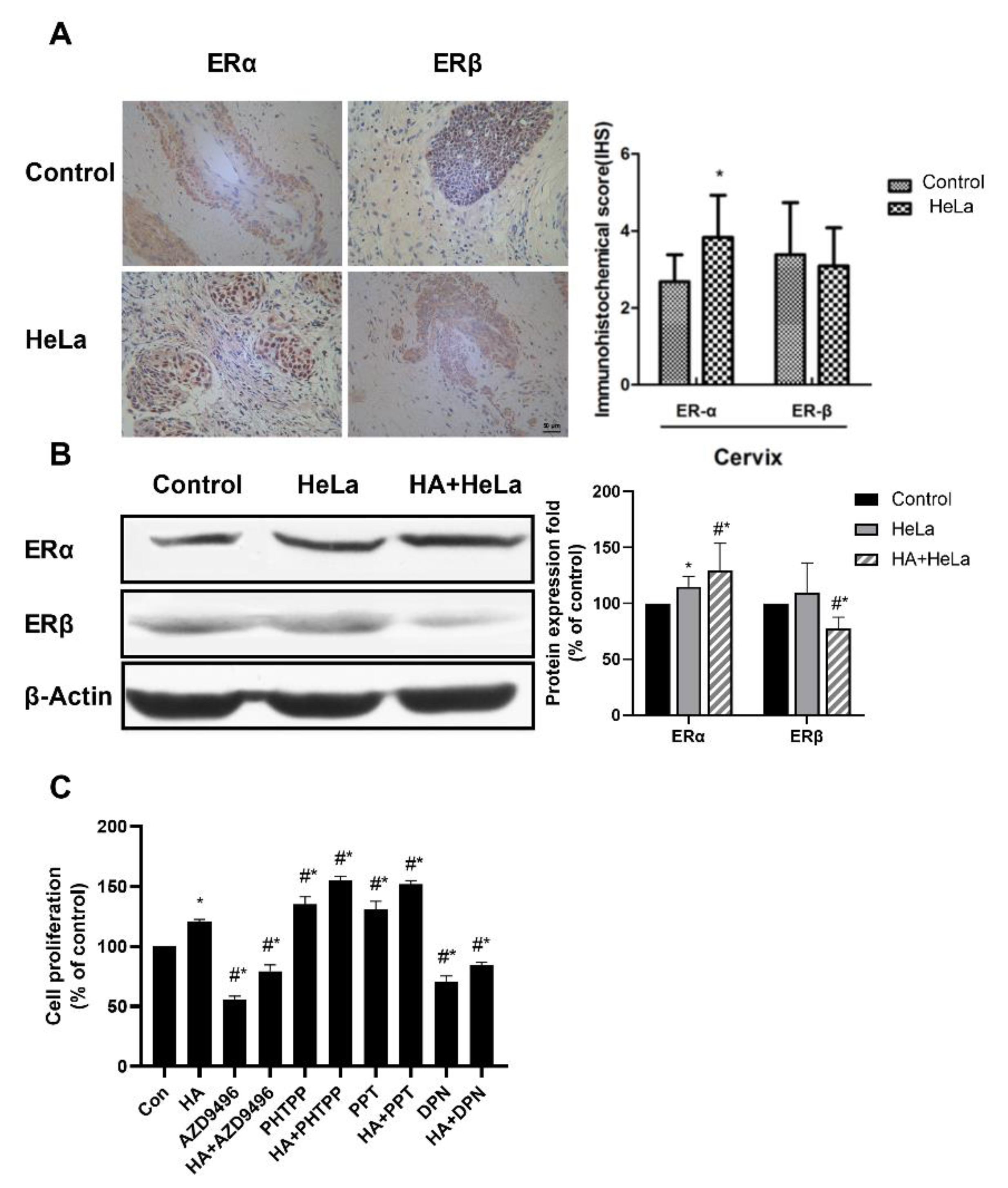
2.2. Histamine Induced Cervical Tumor Growth by Altering the Expression of Estrogen Receptor
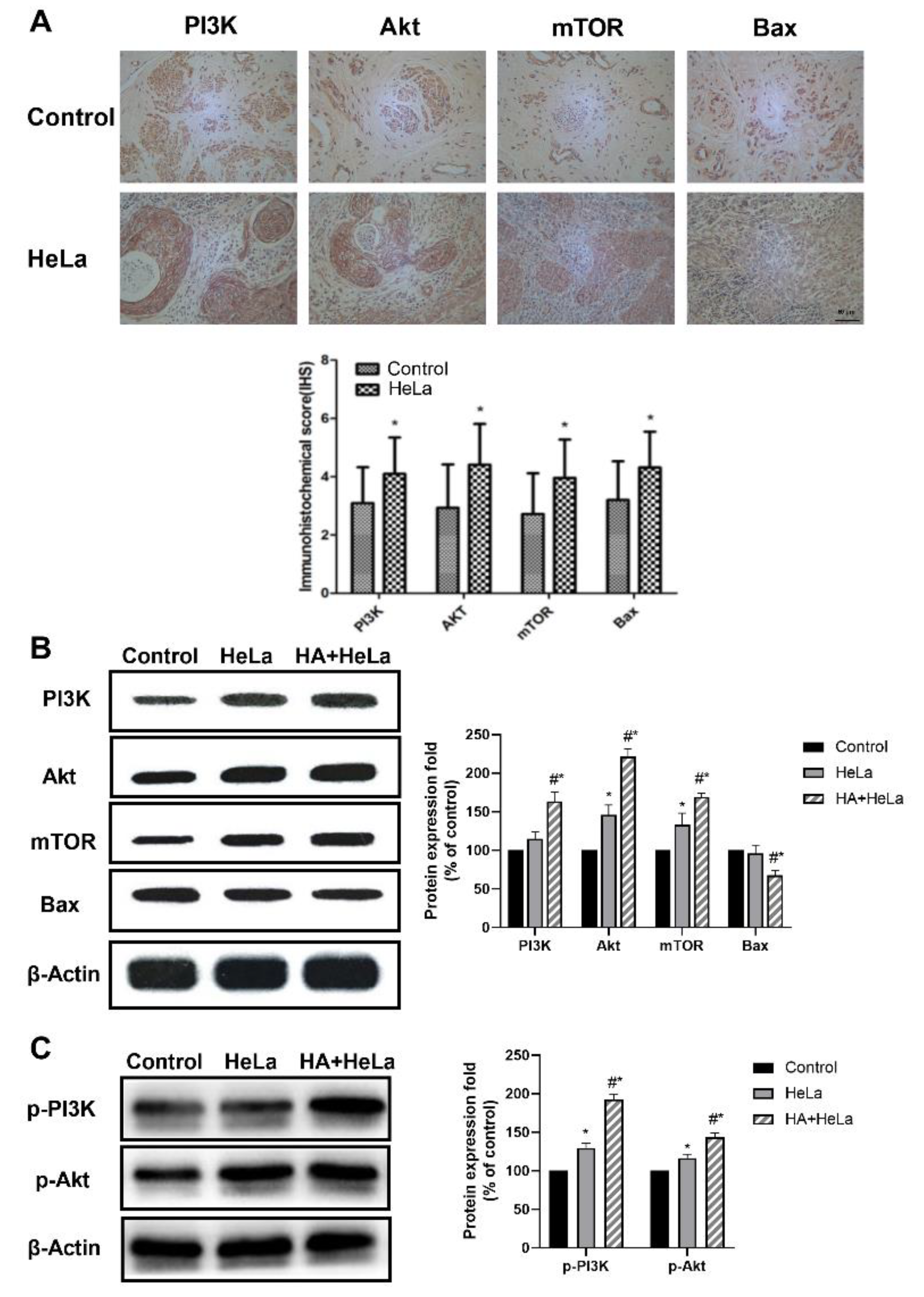
2.3. The PI3K/Akt/mTOR Pathway Is Activated by Histamine
2.4. Inhibition Effects of Apigenin on HeLa Cell Proliferation
2.5. Apigenin Inhibited Cervical Tumor Growth In Vivo by Attenuating the Abnormal ER Signaling Caused by Histamine
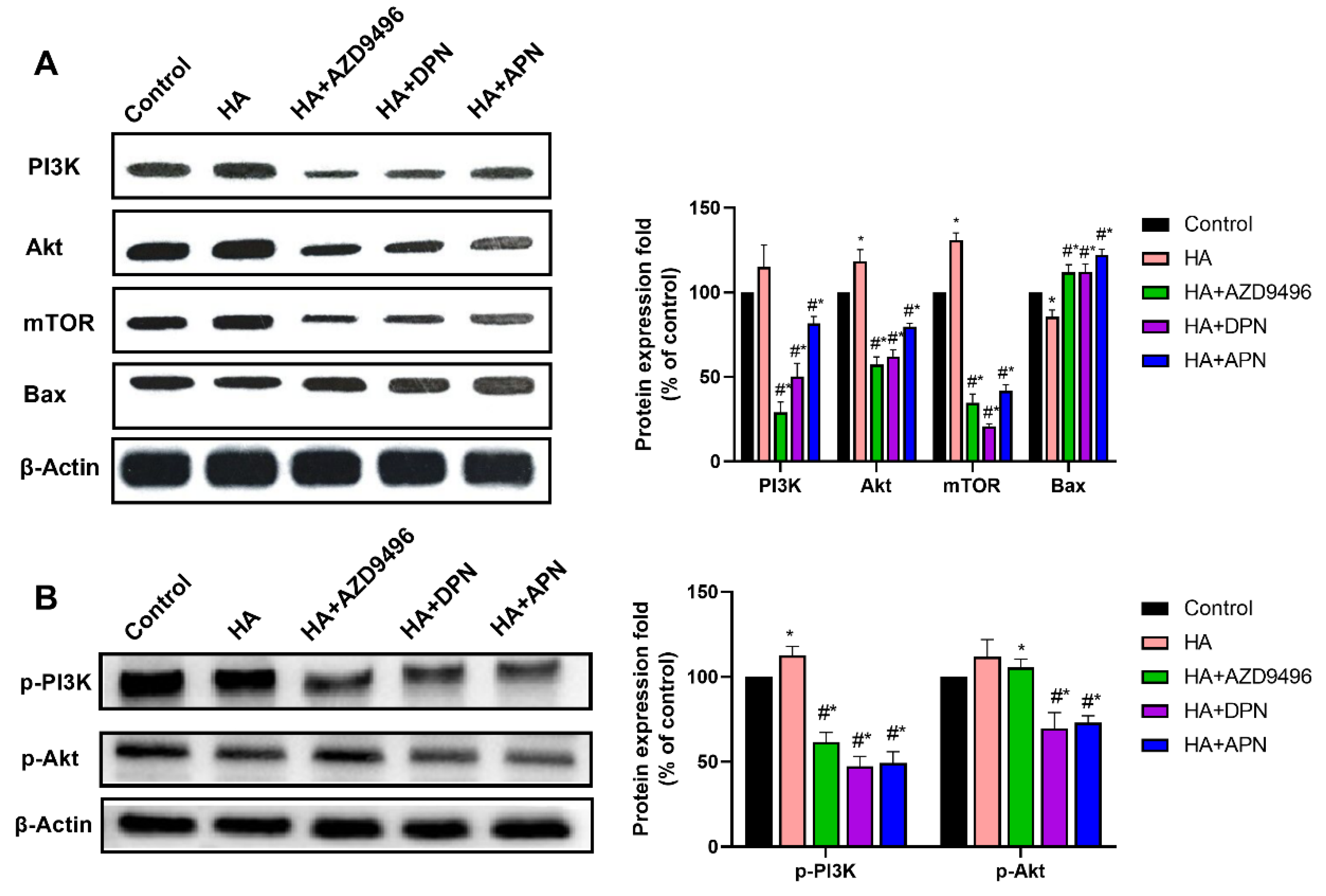
2.6. PI3K/Akt/mTOR Pathway Was Inhibited by the ER Expression Level Change Caused by Apigenin Treatment
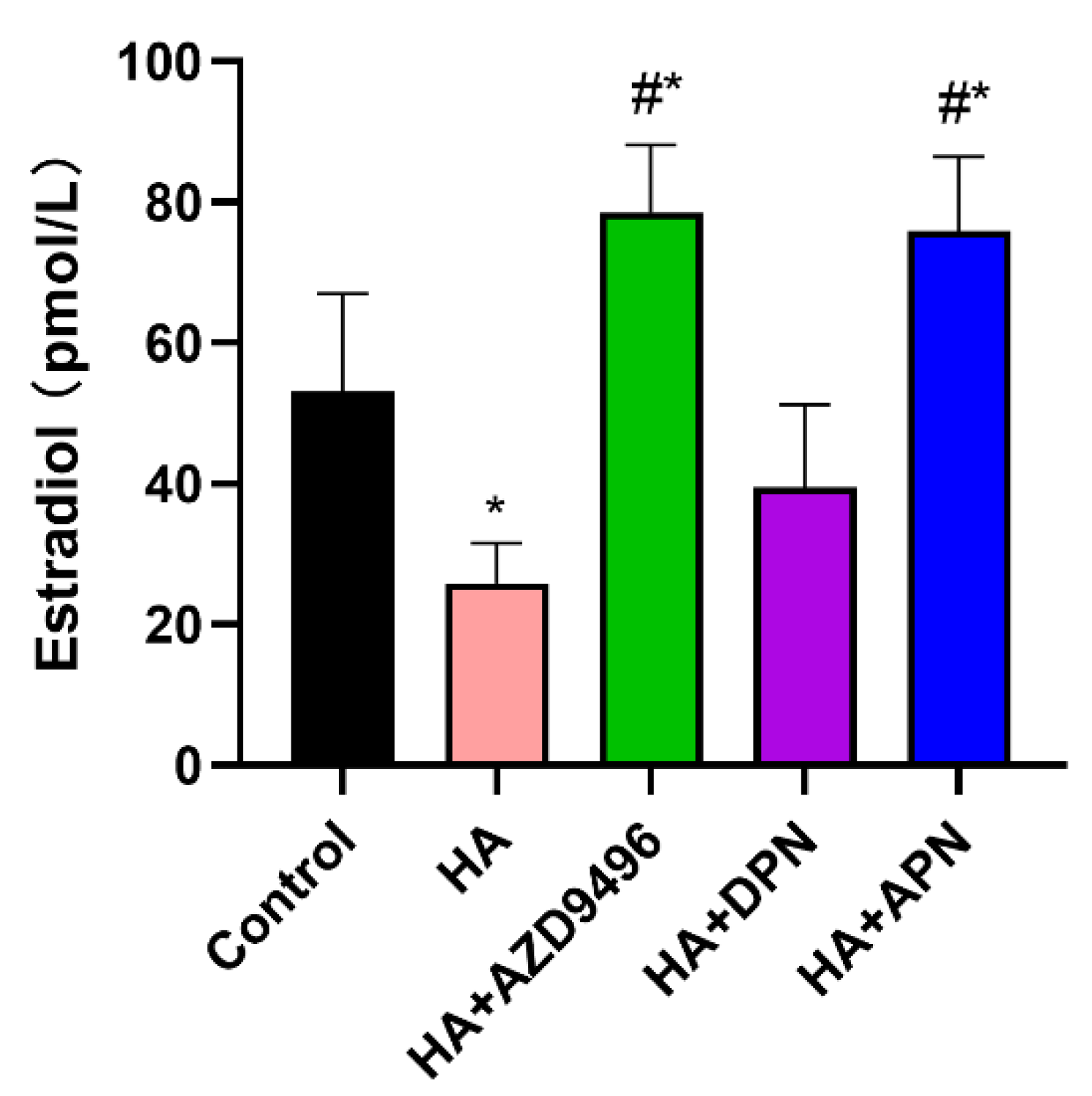
2.7. Apigenin Modulated the Serum Estradiol Level in Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals and Antibodies
4.2. Cell Culture and Treatment
4.3. Determination of Cell Proliferation
4.4. Animal Model
4.5. ELISA Assays for Serum Cytokine Levels and Serum Estradiol Level
4.6. Immunohistochemistry Analysis
4.7. Real-Time PCR
4.8. Western Blot Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.D.; Chen, Z.; Saller, C.; Tarvin, K.; Carvalho, A.L.; Scapulatempo-Neto, C.; Silveira, H.C.; Fregnani, J.H.; Creighton, C.J. Integrated genomic and molecular characterization of cervical cancer. Nature 2017, 543, 378. [Google Scholar]
- Uyar, D.; Rader, J. Genomics of cervical cancer and the role of human papillomavirus pathobiology. Clin. Chem. 2014, 60, 144–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.-Z.; Peng, Y.; Cao, L.-Y.; Chen, Y.-S.; Li, K.; Fu, B.-H. Value of apparent diffusion coefficient (ADC) in assessing radiotherapy and chemotherapy success in cervical cancer. Magn. Reason Imag. 2015, 33, 516–524. [Google Scholar] [CrossRef]
- Small, W., Jr.; Bacon, M.A.; Bajaj, A.; Chuang, L.T.; Fisher, B.J.; Harkenrider, M.M.; Jhingran, A.; Kitchener, H.C.; Mileshkin, L.R.; Viswanathan, A.N.; et al. Cervical cancer: A global health crisis. Cancer 2017, 123, 2404–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishayee, A.; Sethi, G. Bioactive natural products in cancer prevention and therapy: Progress and promise. Semin. Cancer Biol. 2016, 40–41, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Shukla, S.; Gupta, S. Apigenin and cancer chemoprevention: Progress, potential and promise. Int. J. Oncol. 2007, 30, 233–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrott, K.M.; Wiley, C.D.; Desprez, P.-Y.; Campisi, J. Apigenin suppresses the senescence-associated secretory phenotype and paracrine effects on breast cancer cells. Geroscience 2017, 39, 161–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Jin, H.; Pi, J.; Bai, H.; Yang, F.; Wu, C.; Jiang, J.; Cai, J. Apigenin induced apoptosis in esophageal carcinoma cells by destruction membrane structures. Scanning 2016, 38, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Allred, K.F.; Dykes, L.; Allred, C.D.; Awika, J.M. Enhanced action of apigenin and naringenin combination on estrogen receptor activation in non-malignant colonocytes: Implications on sorghum-derived phytoestrogens. Food Funct. 2015, 6, 749–755. [Google Scholar] [CrossRef]
- Sakamoto, T.; Horiguchi, H.; Oguma, E.; Kayama, F. Effects of diverse dietary phytoestrogens on cell growth, cell cycle and apoptosis in estrogen-receptor-positive breast cancer cells. J. Nutr. Biochem. 2010, 21, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Hwang, K.-A.; Choi, K.-C. Treatment with kaempferol suppresses breast cancer cell growth caused by estrogen and triclosan in cellular and xenograft breast cancer models. J. Nutr. Biochem. 2016, 28, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Medina, V.A.; Rivera, E.S. Histamine receptors and cancer pharmacology. Br. J. Pharmacol. 2010, 161, 755–767. [Google Scholar] [CrossRef] [Green Version]
- Rigoni, A.; Colombo, M.; Pucillo, C. The role of mast cells in molding the tumor microenvironment. Cancer Microenviron. 2015, 8, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, L.; Hodges, K.; Meng, F.; Alpini, G.; Francis, H. Histamine and histamine receptor regulation of gastrointestinal cancers. Transl Gastrointest Cancer 2012, 1, 215. [Google Scholar]
- Chen, J.; Hu, X.Y. Inhibition of histamine receptor H3R suppresses prostate cancer growth, invasion and increases apoptosis via the AR pathway. Oncol Lett. 2018, 16, 4921–4928. [Google Scholar] [CrossRef] [Green Version]
- Rae, J.M.; Johnson, M.D.; Scheys, J.O.; Cordero, K.E.; Larios, J.M.; Lippman, M.E. GREB1 is a critical regulator of hormone dependent breast cancer growth. Breast Cancer Res. Treat. 2005, 92, 141–149. [Google Scholar] [CrossRef]
- Judit, R.; Xiaohua, N.; Michael, H.; Wentzel, E.A.; Amirali Hassanzadeh, S.; Chowdhury, W.H.; Kudrolli, T.A.; Srinivasan, Y.; Jun, L.; Ron, R. miR-21: An androgen receptor-regulated microRNA that promotes hormone-dependent and hormone-independent prostate cancer growth. Cancer Res. 2009, 69, 7165. [Google Scholar]
- Fernández-Nogueira, P.; Noguera-Castells, A.; Fuster, G.; Recalde-Percaz, L.; Moragas, N.; López-Plana, A.; Enreig, E.; Jauregui, P.; Carbó, N.; Almendro, V. Histamine receptor 1 inhibition enhances antitumor therapeutic responses through extracellular signal-regulated kinase (ERK) activation in breast cancer. Cancer Lett. 2018, 424, 70–83. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423. [Google Scholar] [CrossRef]
- Rivera, E.S.; Cricco, G.P.; Engel, N.I.; Fitzsimons, C.P.; Martín, G.A.; Bergoc, R.M. Histamine as an autocrine growth factor: An unusual role for a widespread mediator. Semin. Cancer Biol. 2000, 10, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Cricco, G.; Martín, G.; Medina, V.; Núñez, M.; Gutiérrez, A.; Cocca, C.; Bergoc, R.; Rivera, E. Histamine regulates the MAPK pathway via the H 2 receptor in PANC-1 human cells. Inflamm. Res. 2004, 53, S65–S66. [Google Scholar] [CrossRef] [PubMed]
- Davio, C.; Baldi, A.; Mladovan, A.; Cricco, G.; Fitzsimons, C.; Bergoc, R.; Rivera, E. Expression of histamine receptors in different cell lines derived from mammary gland and human breast carcinomas. Inflamm. Res. 1995, 44, S70–S71. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Pi, C.; Wang, G. Inhibition of PI3K/Akt/mTOR pathway by apigenin induces apoptosis and autophagy in hepatocellular carcinoma cells. Biomed. Pharmacother. 2018, 103, 699–707. [Google Scholar] [CrossRef]
- Chiang, L.-C.; Ng, L.T.; Lin, I.C.; Kuo, P.-L.; Lin, C.-C. Anti-proliferative effect of apigenin and its apoptotic induction in human Hep G2 cells. Cancer Lett. 2006, 237, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; Van Der Saag, P.T.; Van Der Burg, B.; Gustafsson, J.-A.K. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor β. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef]
- Long, X.; Fan, M.; Bigsby, R.M.; Nephew, K.P. Apigenin inhibits antiestrogen-resistant breast cancer cell growth through estrogen receptor-α-dependent and estrogen receptor-α-independent mechanisms. Mol. Cancer Ther. 2008, 7, 2096–2108. [Google Scholar] [CrossRef] [Green Version]
- Mak, P.; Leung, Y.-K.; Tang, W.-Y.; Harwood, C.; Ho, S.-M. Apigenin suppresses cancer cell growth through ERβ. Neoplasia 2006, 8, 896–904. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.K.-L.; Leung, T.H.-Y.; Chan, D.W.; Wei, N.; Lau, G.T.-Y.; Liu, S.S.; Siu, M.K.; Ngan, H.Y.-S. Targeting estrogen receptor subtypes (ERa and ERb) with selective ER modulators in ovarian cancer. J. Endocrinol. 2014, 221, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Li, Y.; Wen, Z.; Kong, F.; Guan, X.; Liu, W. microRNA-206 overexpression inhibits cellular proliferation and invasion of estrogen receptor α-positive ovarian cancer cells. Mol. Med. Rep. 2014, 9, 1703–1708. [Google Scholar] [CrossRef] [Green Version]
- Ploch, E. Hormonal replacement therapy in patients after cervical cancer treatment. Gynecol. Oncol. 1987, 26, 169–177. [Google Scholar] [CrossRef]
- Lin, C.-C.; Tsai, Y.-L.; Ho, C.-T.; Teng, S.-C. Determination of the differential estrogenicity of isoflavonoids by E2–ER–ERE-dependent gene expression in recombinant yeast and MCF-7 human breast cancer cells. Food Chem. 2008, 108, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, Q.; Feng, X.; Bo, Y. Effect of evodiagenine mediates photocytotoxicity on human breast cancer cells MDA-MB-231 through inhibition of PI3K/AKT/mTOR and activation of p38 pathways. Fitoterapia 2014, 99, 292–299. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds of apigenin are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Descriptions | Comment |
|---|---|---|
| Control | No HeLa cells injected | Negative control to see the effects of other treatment |
| HeLa | Injected with 2 × 106 HeLa cells | Positive control to mimic cervical cancer |
| HA | Intraperitoneally injected with 1 mg/kg histamine every 3 days | Test group to study the effects of histamine on cervical cancer |
| HA + APG | Intraperitoneally injected with 1 mg/kg histamine and 100 mg/kg apigenin every 3 days | Test group to study the effects of apigenin on cervical cancer |
| HA + AZD9496 | Intraperitoneally injected with 1 mg/kg histamine and 2 mg/kg AZD9496 every 3 days | Test group to study the effects of ERα on cervical cancer |
| HA + DPN | Intraperitoneally injected with 1 mg/kg histamine and 2 mg/kg DPN every 3 days | Test group to study the effects of ERβ on cervical cancer |
| Gene | Primer Sequence (5′–3′) | Size of Product |
|---|---|---|
| β-actin | CTCGCCTTTGCCGATCC | 258 bp |
| GGGGTACTTCAGGGTGAGGA | ||
| ERα | ATGCGCTGCGTCGCCTCTAAC | 78 bp |
| CGCAGGGCAGAAGGCTCAGA | ||
| ERβ | AGCGCGGAGGCTGCGAGAAAT | 56 bp |
| CCTGCTCTTCGCCCTGCAAGTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, E.; Zhang, Y.; Fan, Z.; Cheng, L.; Han, S.; Che, H. Apigenin Inhibits Histamine-Induced Cervical Cancer Tumor Growth by Regulating Estrogen Receptor Expression. Molecules 2020, 25, 1960. https://doi.org/10.3390/molecules25081960
Zhang E, Zhang Y, Fan Z, Cheng L, Han S, Che H. Apigenin Inhibits Histamine-Induced Cervical Cancer Tumor Growth by Regulating Estrogen Receptor Expression. Molecules. 2020; 25(8):1960. https://doi.org/10.3390/molecules25081960
Chicago/Turabian StyleZhang, Erkang, Yani Zhang, Zhuoyan Fan, Lei Cheng, Shiwen Han, and Huilian Che. 2020. "Apigenin Inhibits Histamine-Induced Cervical Cancer Tumor Growth by Regulating Estrogen Receptor Expression" Molecules 25, no. 8: 1960. https://doi.org/10.3390/molecules25081960
APA StyleZhang, E., Zhang, Y., Fan, Z., Cheng, L., Han, S., & Che, H. (2020). Apigenin Inhibits Histamine-Induced Cervical Cancer Tumor Growth by Regulating Estrogen Receptor Expression. Molecules, 25(8), 1960. https://doi.org/10.3390/molecules25081960