
Overview of Inositol and Inositol Phosphates on Chemoprevention of Colitis-Induced Carcinogenesis


{kind=link}
Abstract
:1. Introduction
2. Nature and Sources of Inositol Compounds
3. Biosynthesis and Metabolism of Inositol and Its Phosphates
4. Potential Nutritional Deficiency of Inositol and Inositol Phosphates
5. Chemopreventive Effects of Inositol and InsP6 on Cancer—Colitis-Induced Carcinogenesis as a Model
5.1. Effects of Inositol and InsP6 on Inhibiting Dextran Sulfate Sodium (DSS)-Induced Ulcerative Colitis (UC) and Carcinogenesis in Rodents
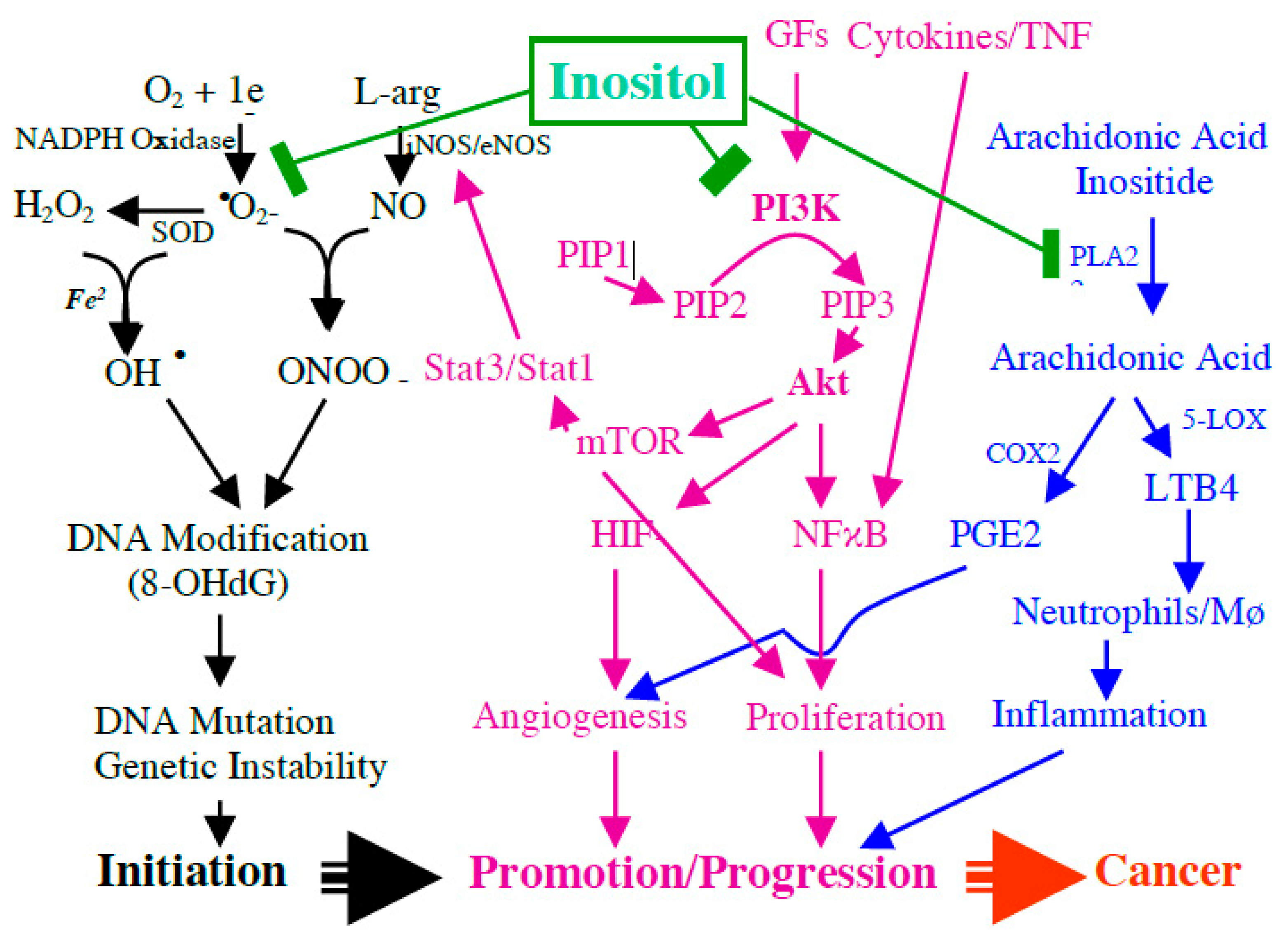
5.2. Potential Mechanism of Inositol and Its Phosphates against Inflammation and Carcinogenesis
5.2.1. PI3K Signaling and Gut Epithelial Progenitor Cell
5.2.2. Antioxidant
5.2.3. Effects of Inositol on the Mucosal and Anti-Tumor Immunity
5.2.4. Effects of Inositol and InsP6 on Cell Survival, Proliferation, and Differentiation
5.2.5. Effects of Inositol and InsP6 on DNA Damage Repair
5.3. Pharmacokinetics of InsP6
6. Summary, Conclusions, and Future Direction
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bizzarri, M.; Fuso, A.; DiNicola, S.; Cucina, A.; Bevilacqua, A. Pharmacodynamics and pharmacokinetics of inositol(s) in health and disease. Expert Opin. Drug Metab. Toxicol. 2016, 12, 1181–1196. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, M.; DiNicola, S.; Bevilacqua, A.; Cucina, A. Broad Spectrum Anticancer Activity of Myo-Inositol and Inositol Hexakisphosphate. Int. J. Endocrinol. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Shamsuddin, A.M. Metabolism and cellular functions of IP6: A review. Anticancer Res. 2000, 19, 3733–3736. [Google Scholar]
- Clements, R.S.; Darnell, B. Myo-inositol content of common foods: Development of a high-myo-inositol diet. Am. J. Clin. Nutr. 1980, 33, 1954–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, K.; Vucenik, I.; Shamsuddin, A.M. [3H]Phytic Acid (Inositol Hexaphosphate) Is Absorbed and Distributed to Various Tissues in Rats. J. Nutr. 1993, 123, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, F., Jr. D-myoinositol 1-phosphate as product of cyclization of glucose 6-phosphate and sub-strate for a specific phosphatase in rat testis. J. Biol. Chem. 1967, 242, 1375–1382. [Google Scholar]
- Eisenberg, F.; Parthasarathy, R. Measurement of biosynthesis of myo-inositol from glucose 6-phosphate. Methods Enzymol. 1987, 141, 127–143. [Google Scholar] [CrossRef]
- Falkenburger, B.H.; Jensen, J.B.; Dickson, E.J.; Suh, B.-C.; Hille, B. SYMPOSIUM REVIEW: Phosphoinositides: Lipid regulators of membrane proteins. J. Physiol. 2010, 588, 3179–3185. [Google Scholar] [CrossRef]
- Clements, R.S., Jr.; Reynertson, R. Myoinositol metabolism in diabetes mellitus. Effect of insulin treatment. Diabetes 1977, 26, 215–221. [Google Scholar] [CrossRef]
- Jimenez, M.A.; Scarino, M.L.; Vignolini, F.; Mengheri, E. Evidence that Polyunsaturated Lecithin Induces a Reduction in Plasma Cholesterol Level and Favorable Changes in Lipoprotein Composition in Hypercholesterolemic Rats. J. Nutr. 1990, 120, 659–667. [Google Scholar] [CrossRef]
- Irvine, R.; Schell, M.J. Back in the water: The return of the inositol phosphates. Nat. Rev. Mol. Cell Biol. 2001, 2, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Graf, E.; Eaton, J.W. Dietary suppression of colonic cancer fiber or phytate? Cancer 1985, 56, 717–718. [Google Scholar] [CrossRef]
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53, S330–S375. [Google Scholar] [CrossRef]
- Wilson, M.S.C.; Livermore, T.M.; Saiardi, A. Inositol pyrophosphates: Between signalling and metabolism. Biochem. J. 2013, 452, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Takasuga, S.; Sasaki, J.; Kofuji, S.; Eguchi, S.; Yamazaki, M.; Suzuki, A. Mammalian phosphoinositide kinases and phosphatases. Prog. Lipid Res. 2009, 48, 307–343. [Google Scholar] [CrossRef] [PubMed]
- Gardocki, M.E.; Jani, N.; Lopes, J.M. Phosphatidylinositol biosynthesis: Biochemistry and regulation. Biochim. et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2005, 1735, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, L.K.; Seelan, R.S.; Tobias, C.; Casanova, M.F.; Parthasarathy, R.N. Mammalian inositol 3-phosphate synthase: Its role in the biosynthesis of brain inositol and its clinical use as a psychoactive agent. Subcell Biochem. 2006, 39, 293–314. [Google Scholar] [PubMed]
- Raboy, V.; Gerbasi, P. Genetics of myo-Inositol Phosphate Synthesis and Accumulation. Macromol. Protein Complexes III Struct. Funct. 1996, 26, 257–285. [Google Scholar] [CrossRef]
- Chakraborty, A.; Kim, S.; Snyder, S.H. Inositol Pyrophosphates as Mammalian Cell Signals. Sci. Signal. 2011, 4, re1. [Google Scholar] [CrossRef] [Green Version]
- Bennett, M.; Onnebo, S.M.N.; Azevedo, C.; Saiardi, A. Inositol pyrophosphates: Metabolism and signaling. Cell. Mol. Life Sci. 2006, 63, 552–564. [Google Scholar] [CrossRef]
- Saiardi, A.; Bhandari, R.; Resnick, A.C.; Snowman, A.M.; Snyder, S.H. Phosphorylation of proteins by inositol pyrophosphates. Science 2004, 306, 2101–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.; Goretsky, T.; Managlia, E.; Dirisina, R.; Singh, A.P.; Brown, J.B.; May, R.; Yang, G.-Y.; Ragheb, J.W.; Evers, B.M.; et al. Phosphoinositide 3-kinase signaling mediates beta-catenin activation in intestinal epithe-lial stem and progenitor cells in colitis. Gastroenterology 2010, 139, 869–881.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, M.J.; Irvine, R.F. Inositol phosphates and cell signalling. Nat. Cell Biol. 1989, 341, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Putney, J.W. Formation and actions of calcium-mobilizing messenger, inositol 1,4,5-trisphosphate. Am. J. Physiol. 1987, 252, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Streb, H.; Schulz, I. Regulation of cytosolic free Ca2+ concentration in acinar cells of rat pancreas. Am. J. Physiol. 1983, 245, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Streb, H.; Irvine, R.F.; Berridge, M.J.; Schulz, I. Release of Ca2+ from a nonmitochondrial intracellular store in pancreatic acinar cells by inositol-1,4,5-trisphosphate. Nat. Cell Biol. 1983, 306, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Shears, S.B. Diphosphoinositol polyphosphates: Metabolic messengers? Mol. Pharmacol. 2009, 76, 236–252. [Google Scholar] [CrossRef] [Green Version]
- Sasakawa, N.; Sharif, M.; Hanley, M.R. Metabolism and biological activities of inositol pen-takisphosphate and inositol hexakisphosphate. Biochem. Pharmacol. 1995, 50, 137–146. [Google Scholar] [CrossRef]
- Shears, S.B. Assessing the omnipotence of inositol hexakisphosphate. Cell. Signal. 2001, 13, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Barker, C.J.; Illies, C.; Gaboardi, G.C.; Berggren, P.-O. Inositol pyrophosphates: Structure, enzymology and function. Cell. Mol. Life Sci. 2009, 66, 3851–3871. [Google Scholar] [CrossRef]
- DiNicola, S.; Minini, M.; Unfer, V.; Verna, R.; Cucina, A.; Bizzarri, M. Nutritional and Acquired Deficiencies in Inositol Bioavailability. Correlations with Metabolic Disorders. Int. J. Mol. Sci. 2017, 18, 2187. [Google Scholar] [CrossRef] [PubMed]
- Laster, J.; Frame, L.A. Beyond the Calories-Is the Problem in the Processing? Curr. Treat. Options Gastroenterol. 2019, 17, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Chagas, P.; Chiva-Blanch, G. Diet and Cardiovascular Disease: Effects of Foods and Nutrients in Classical and Emerging Cardiovascular Risk Factors. Curr. Med. Chem. 2019, 26, 3639–3651. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Morenga, L.T. Carbohydrate quality and human health: A series of systematic reviews and me-ta-analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef] [Green Version]
- Arsenault, J.E.; Brown, K.H. Zinc intake of US preschool children exceeds new dietary reference in-takes. Am. J. Clin. Nutr. 2003, 78, 1011–1017. [Google Scholar] [CrossRef] [Green Version]
- Gibson, R.S.; Vanderkooy, P.D.S.; Thompson, L. Dietary phytate x calcium/zinc millimolar ratios and zinc nutriture in some Ontario preschool children. Biol. Trace Elem. Res. 1991, 30, 87–94. [Google Scholar] [CrossRef]
- Vitolins, M.Z.; Quandt, S.A.; Case, L.D.; Bell, R.A.; Arcury, T.A.; McDonald, J. Vitamin and mineral supplement use by older rural adults. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2000, 55, M613–M617. [Google Scholar] [CrossRef] [Green Version]
- Harland, B.F.; Smith, S.A.; Howard, M.P.; Ellis, R.; Smith, J.C. Nutritional status and phytate:zinc and phytate x calcium:zinc dietary molar ratios of lacto-ovo vegetarian Trappist monks: 10 years later. J. Am. Diet. Assoc. 1988, 88, 1562–1566. [Google Scholar]
- Phelps, D.L.; Ward, R.M.; Williams, R.L.; Nolen, T.L.; Watterberg, K.L.; Oh, W.; Goedecke, M.; Ehrenkranz, R.A.; Fennell, T.; Poindexter, B.B.; et al. Safety and pharmacokinetics of multiple dose myo-inositol in preterm infants. Pediatr. Res. 2016, 80, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Vucenik, I.; Shamsuddin, A.M. Cancer Inhibition by Inositol Hexaphosphate (IP6) and Inositol: From Laboratory to Clinic. J. Nutr. 2003, 133, 3778S–3784S. [Google Scholar] [CrossRef]
- Vucenik, I.; Yang, G.Y.; Shamsuddin, A.M. Inositol hexaphosphate and inositol inhibit DMBA-induced rat mammary cancer. Carcinogenesis 1995, 16, 1055–1058. [Google Scholar] [CrossRef] [PubMed]
- Wattenberg, L.W.; Estensen, R.D. Studies of chemopreventive effects of myo-inositol on ben-zo[a]pyrene-induced neoplasia of the lung and forestomach of female A/J mice. Carcinogenesis 1993, 14, 1975–1977. [Google Scholar]
- Ullah, A.; Shamsuddin, A.M. Dose-dependent inhibition of large intestinal cancer by inositol hexa-phosphate in F344 rats. Carcinogenesis 1990, 11, 2219–2222. [Google Scholar] [CrossRef] [PubMed]
- Shamsuddin, A.M.; Ullah, A.; Chakravarthy, A.K. Inositol and inositol hexaphosphate suppress cell proliferation and tumor formation in CD-1 mice. Carcinogenesis 1989, 10, 1461–1463. [Google Scholar] [CrossRef] [PubMed]
- Shamsuddin, A.M.; Elsayed, A.M.; Ullah, A. Suppression of large intestinal cancer in F344 rats by inositol hexaphosphate. Carcinogenesis 1988, 9, 577–580. [Google Scholar] [CrossRef]
- Yang, G.-Y.; Taboada, S.; Liao, J. Inflammatory Bowel Disease: A Model of Chronic Inflammation-Induced Cancer. Methods Mol. Biol. 2009, 511, 193–233. [Google Scholar] [CrossRef]
- Liao, J.; Seril, D.N.; Yang, A.L.; Lu, G.G.; Yang, G.-Y. Inhibition of chronic ulcerative colitis associated adenocarcinoma development in mice by inositol compounds. Carcinogenesis 2006, 28, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Seril, D.N.; Liao, J.; West, A.B.; Yang, G.-Y. High-iron diet: Foe or feat in ulcerative colitis and ulcerative colitis-associated carcin-ogenesis. J. Clin. Gastroenterol. 2006, 40, 391–397. [Google Scholar] [CrossRef]
- Seril, D.N.; Liao, J.; Yang, G.-Y.; Yang, C.S. Oxidative stress and ulcerative colitis-associated carcinogenesis: Studies in humans and animal models. Carcinogenesis 2003, 24, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Seril, D.N.; Liao, J.; Ho, K.-L.K.; Yang, C.S.; Yang, G.-Y. Inhibition of chronic ulcerative colitis-associated colorectal adenocarcinoma devel-opment in a murine model by N-acetylcysteine. Carcinogenesis 2002, 23, 993–1001. [Google Scholar] [CrossRef] [Green Version]
- Seril, D.N.; Liao, J.; Ho, K.-L.K.; Warsi, A.; Yang, C.S.; Yang, G.-Y. Dietary Iron Supplementation Enhances DSS-Induced Colitis and Associated Colorectal Carcinoma Development in Mice. Dig. Dis. Sci. 2002, 47, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.-Y.; Taboada, S.; Liao, J. Induced Nitric Oxide Synthase as a Major Player in the Oncogenic Transformation of Inflamed Tissue. Bioinform. MicroRNA Res. 2009, 512, 119–156. [Google Scholar] [CrossRef]
- Bradford, E.M.; Thompson, C.A.; Goretsky, T.; Yang, G.-Y.; Rodriguez, L.M.; Li, L.; Barrett, T.A. Myo-inositol reduces beta-catenin activation in colitis. World J. Gastroenterol. 2017, 23, 5115–5126. [Google Scholar] [CrossRef] [PubMed]
- Graf, E.; Empson, K.L.; Eaton, J.W. Phytic acid. A natural antioxidant. J. Biol. Chem. 1987, 262, 11647–11650. [Google Scholar] [PubMed]
- Zajdel, A.; Wilczok, A.; Węglarz, L.; Dzierżewicz, Z. Phytic Acid Inhibits Lipid PeroxidationIn Vitro. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Tucci, M.; Benghuzzi, H.; Cason, Z.; Hughes, J. The effects of inositol hexaphosphate on the inflammatory response in transformed RAW 264.7 macrophages. Biomed. Sci. Instrum. 2000, 36, 21–26. [Google Scholar]
- Eggleton, P.; Penhallow, J.; Crawford, N. Priming action of inositol hexakisphosphate (InsP6) on the stimulated respiratory burst in human neutrophils. Biochim. et Biophys. Acta (BBA) Bioenerg. 1991, 1094, 309–316. [Google Scholar] [CrossRef]
- Eggleton, P. Effect of IP6 on human neutrophil cytokine production and cell morphology. Anticancer Res. 2000, 19, 3711–3715. [Google Scholar]
- Wawszczyk, J.; Orchel, A.; Kapral, M.; Węglarz, L. Role of protein tyrosine kinase in the effect of IP6 on IL-8 secretion in intestinal epithelial cells. Acta Pol. Pharm. Drug Res. 2013, 70, 79–86. [Google Scholar]
- Wawszczyk, J.; Orchel, A.; Kapral, M.; Hollek, A.; Weglarz, L. Phytic acid down-regulates IL-8 secretion from colonic epithelial cells by influ-encing mitogen-activated protein kinase signaling pathway. Acta Pol. Pharm. 2012, 69, 1276–1282. [Google Scholar]
- Weglarz, L.; Wawszczyk, J.; Orchel, A.; Jaworska-Kik, M.; Dzierzewicz, Z. Phytic acid modulates in vitro IL-8 and IL-6 release from colonic epithelial cells stimulated with LPS and IL-1beta. Dig. Dis. Sci. 2007, 52, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Baten, A.; Ullah, A.; Tomazic, V.J.; Shamsuddin, A.M. Inositol-phosphate-induced enhancement of natural killer cell activity correlates with tumor suppression. Carcinogenesis 1989, 10, 1595–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z. Inositol hexaphosphate-induced enhancement of natural killer cell activity correlates with suppression of colon carcinogenesis in rats. World J. Gastroenterol. 2005, 11, 5044–5046. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Y.; Shamsuddin, A.M. IP6-induced growth inhibition and differentiation of HT-29 human colon cancer cells: Involvement of intracellular inositol phosphates. Anticancer Res. 1995, 15, 2479–2487. [Google Scholar]
- Yoon, M.R.; Nam, S.H.; Kang, M.Y. Antioxidative and Antimutagenic Activities of 70% Ethanolic Extracts from Four Fungal Mycelia-Fermented Specialty Rices. J. Clin. Biochem. Nutr. 2008, 43, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Hanakahi, L.A.; Bartlet-Jones, M.; Chappell, C.; Pappin, D.; West, S.C. Binding of Inositol Phosphate to DNA-PK and Stimulation of Double-Strand Break Repair. Cell 2000, 102, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Lieber, M.R. Binding of Inositol Hexakisphosphate (IP6) to Ku but Not to DNA-PKcs. J. Biol. Chem. 2002, 277, 10756–10759. [Google Scholar] [CrossRef] [Green Version]
- Byrum, J.; Jordan, S.; Safrany, S.T.; Rodgers, W. Visualization of inositol phosphate-dependent mobility of Ku: Depletion of the DNA-PK cofactor InsP6 inhibits Ku mobility. Nucleic Acids Res. 2004, 32, 2776–2784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, J.C.; Salerno, B.; Hanakahi, L.A. Evidence for an inositol hexakisphosphate-dependent role for Ku in mammalian nonhomologous end joining that is independent of its role in the DNA-dependent protein kinase. Nucleic Acids Res. 2008, 36, 5713–5726. [Google Scholar] [CrossRef]
- Shen, X.; Xiao, H.; Ranallo, R.; Wu, W.-H.; Wu, C. Modulation of ATP-dependent chromatin-remodeling complexes by inositol polyphos-phates. Science 2003, 299, 112–114. [Google Scholar] [CrossRef] [Green Version]
- Steger, D.J.; Haswell, E.S.; Miller, A.L.; Wente, S.R.; O’Shea, E.K. Regulation of chromatin remodeling by inositol polyphosphates. Science 2003, 299, 114–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Ma, W.Y.; Hecht, S.S.; Dong, Z. Inositol hexaphosphate inhibits cell transformation and activator protein 1 activation by targeting phosphatidylinositol-3’ kinase. Cancer Res. 1997, 57, 2873–2878. [Google Scholar] [PubMed]
- Eiseman, J.L.; Lan, J.; Guo, J.; Joseph, E.; Vucenik, I. Pharmacokinetics and tissue distribution of inositol hexaphosphate in C.B17 SCID mice bearing human breast cancer xenografts. Metabolism 2011, 60, 1465–1474. [Google Scholar] [CrossRef]
- March, J.; Simonet, B.; Grases, F. Determination of phytic acid by gas chromatography–mass spectroscopy: Application to biological samples. J. Chromatogr. B Biomed. Sci. Appl. 2001, 757, 247–255. [Google Scholar] [CrossRef]
- Grases, F.; Simonet, B.M.; Vucenik, I.; Prieto, R.M.; Costa-Bauzá, A.; March, J.G.; Shamsuddin, A.M. Absorption and excretion of orally administered inositol hexaphosphate (IP6or phytate) in humans. BioFactors 2001, 15, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Bathena, S.P.R.; Huang, J.; Epstein, A.A.; Gendelman, H.E.; Boska, M.D.; Alnouti, Y. Rapid and reliable quantitation of amino acids and myo-inositol in mouse brain by high performance liquid chromatography and tandem mass spectrometry. J. Chromatogr. B 2012, 893-894, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grases, F.; Simonet, B.M.; Vucenik, I.; Perelló, J.; Prieto, R.M.; Shamsuddin, A.M. Effects of exogenous inositol hexakisphosphate (InsP6) on the levels of InsP6 and of inositol trisphosphate (InsP3) in malignant cells, tissues and biological fluids. Life Sci. 2002, 71, 1535–1546. [Google Scholar] [CrossRef]
- Frieler, R.A.; Mitteness, D.J.; Golovko, M.Y.; Gienger, H.M.; Rosenberger, T.A. Quantitative determination of free glycerol and myo-inositol from plasma and tis-sue by high-performance liquid chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 3667–3672. [Google Scholar] [CrossRef] [Green Version]
- Kindt, E.; Shum, Y.; Badura, L.; Snyder, P.J.; Brant, A.; Fountain, S.; Szekely-Klepser, G. Development and validation of an LC/MS/MS procedure for the quantification of en-dogenous myo-inositol concentrations in rat brain tissue homogenates. Anal. Chem. 2004, 76, 4901–4908. [Google Scholar] [CrossRef]
- Yamakoshi, M.; Takahashi, M.; Kouzuma, T.; Imamura, S.; Tsuboi, I.; Kawazu, S.; Yamagata, F.; Tominaga, M.; Noritake, M. Determination of urinary myo-inositol concentration by an improved enzymatic cycling method using myo-inositol dehydrogenase from Flavobacterium sp. Clin. Chim. Acta 2003, 328, 163–171. [Google Scholar] [CrossRef]
- Kouzuma, T.; Takahashi, M.; Endoh, T.; Kaneko, R.; Ura, N.; Shimamoto, K.; Watanabe, N. An enzymatic cycling method for the measurement of myo-inositol in biological samples. Clin. Chim. Acta 2001, 312, 143–151. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weinberg, S.E.; Sun, L.Y.; Yang, A.L.; Liao, J.; Yang, G.Y. Overview of Inositol and Inositol Phosphates on Chemoprevention of Colitis-Induced Carcinogenesis. Molecules 2021, 26, 31. https://doi.org/10.3390/molecules26010031
Weinberg SE, Sun LY, Yang AL, Liao J, Yang GY. Overview of Inositol and Inositol Phosphates on Chemoprevention of Colitis-Induced Carcinogenesis. Molecules. 2021; 26(1):31. https://doi.org/10.3390/molecules26010031
Chicago/Turabian StyleWeinberg, Samuel E., Le Yu Sun, Allison L. Yang, Jie Liao, and Guang Yu Yang. 2021. "Overview of Inositol and Inositol Phosphates on Chemoprevention of Colitis-Induced Carcinogenesis" Molecules 26, no. 1: 31. https://doi.org/10.3390/molecules26010031
APA StyleWeinberg, S. E., Sun, L. Y., Yang, A. L., Liao, J., & Yang, G. Y. (2021). Overview of Inositol and Inositol Phosphates on Chemoprevention of Colitis-Induced Carcinogenesis. Molecules, 26(1), 31. https://doi.org/10.3390/molecules26010031