
Aloe djiboutiensis: Antioxidant Activity, Molecular Networking-Based Approach and In Vivo Toxicity of This Endemic Species in Djibouti

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Antioxidant Activity
2.2. Total Polyphenol and Total Flavonoid Contents
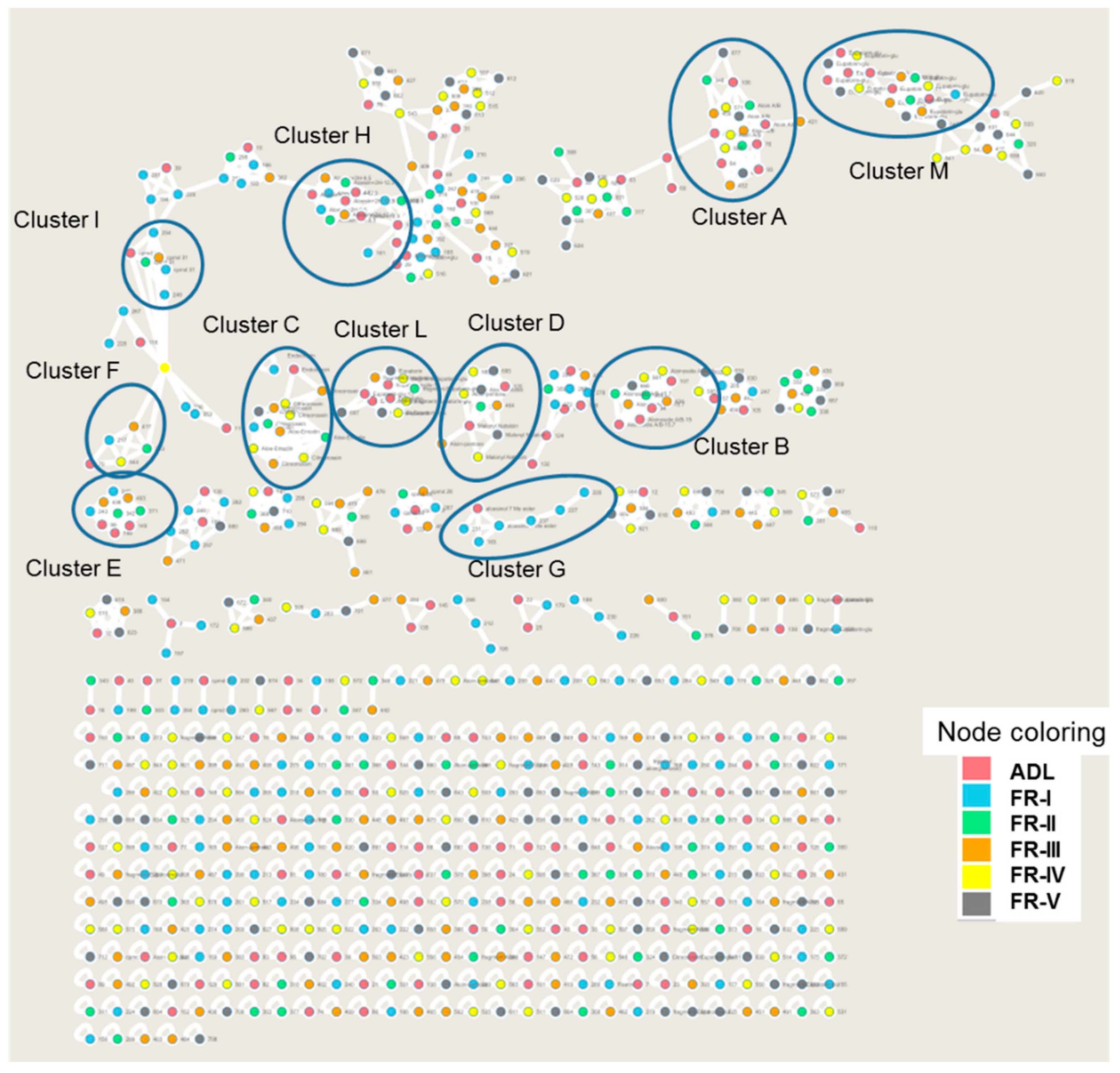
2.3. Identification of Specialized Metabolites Using LC-MS/MS Molecular Networking Based-Approach
2.4. In Vitro Toxicity
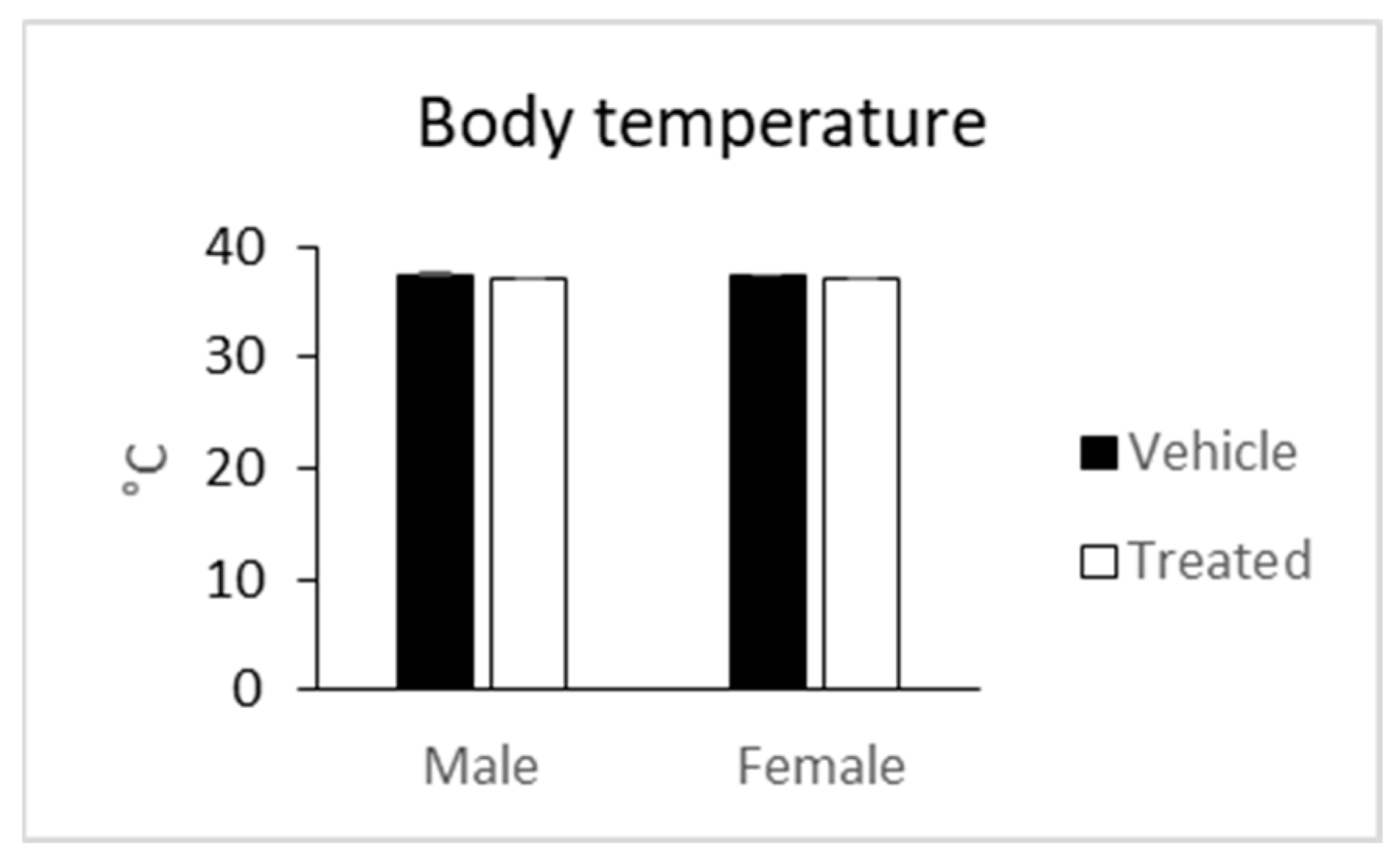
2.5. In Vivo Observations
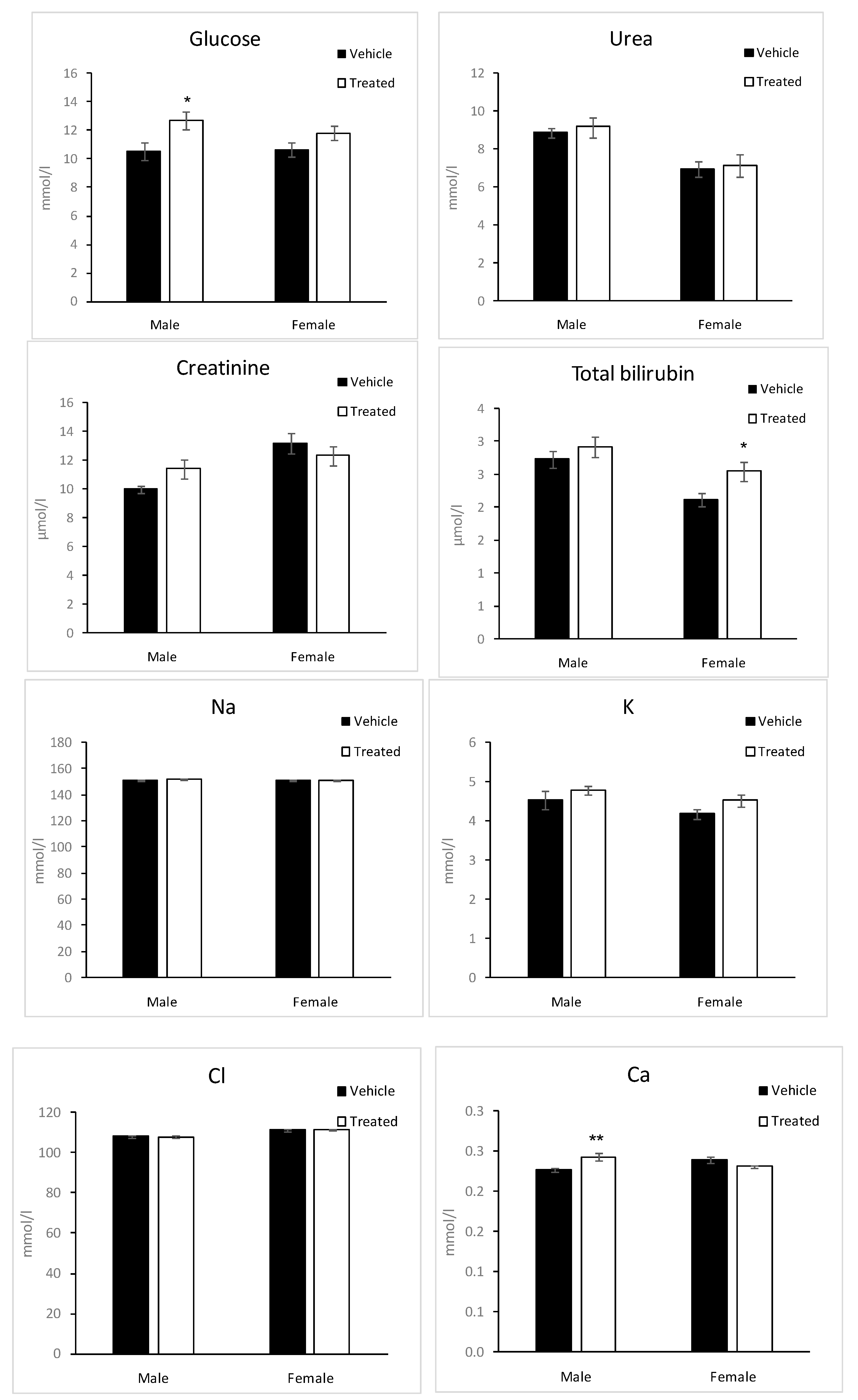
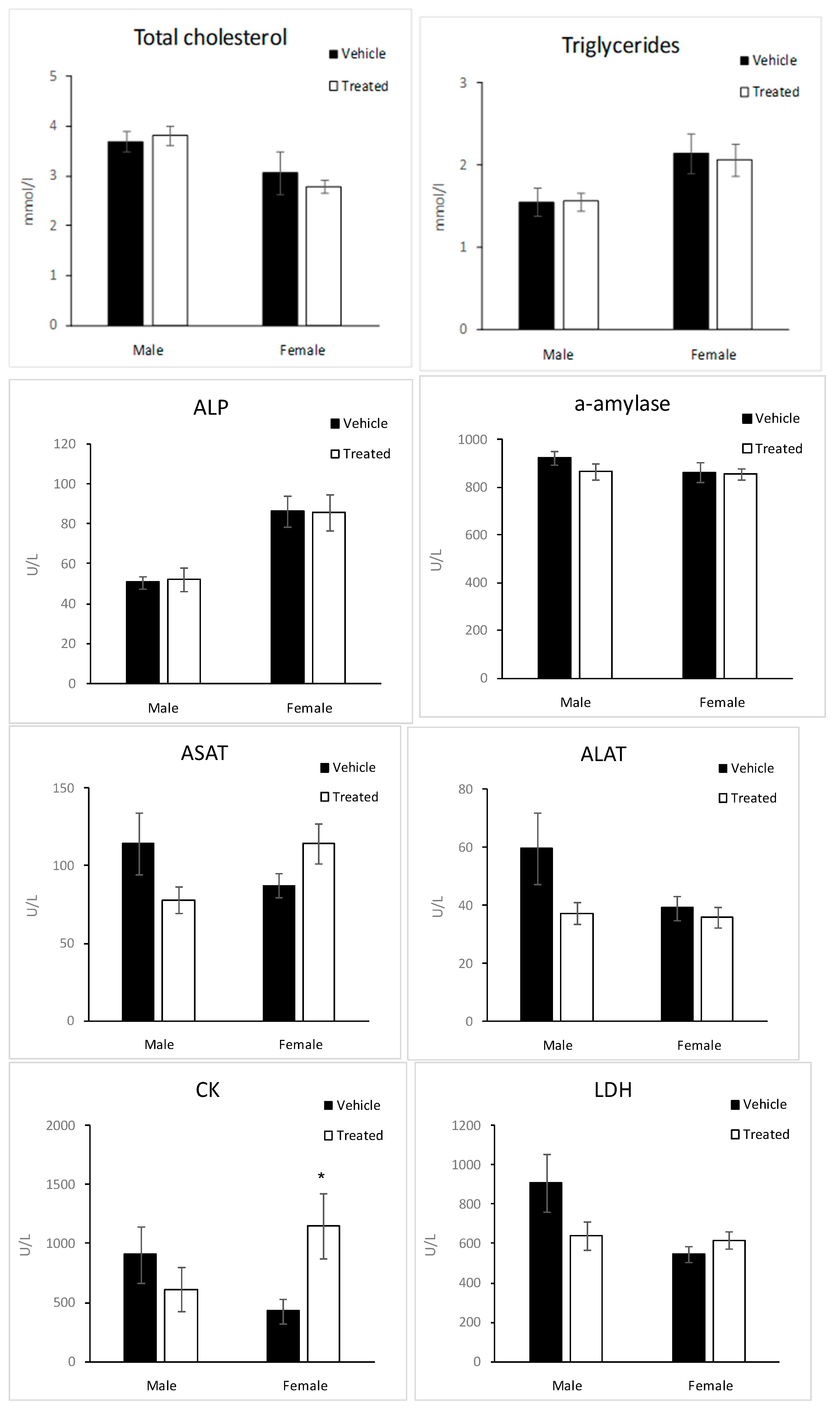
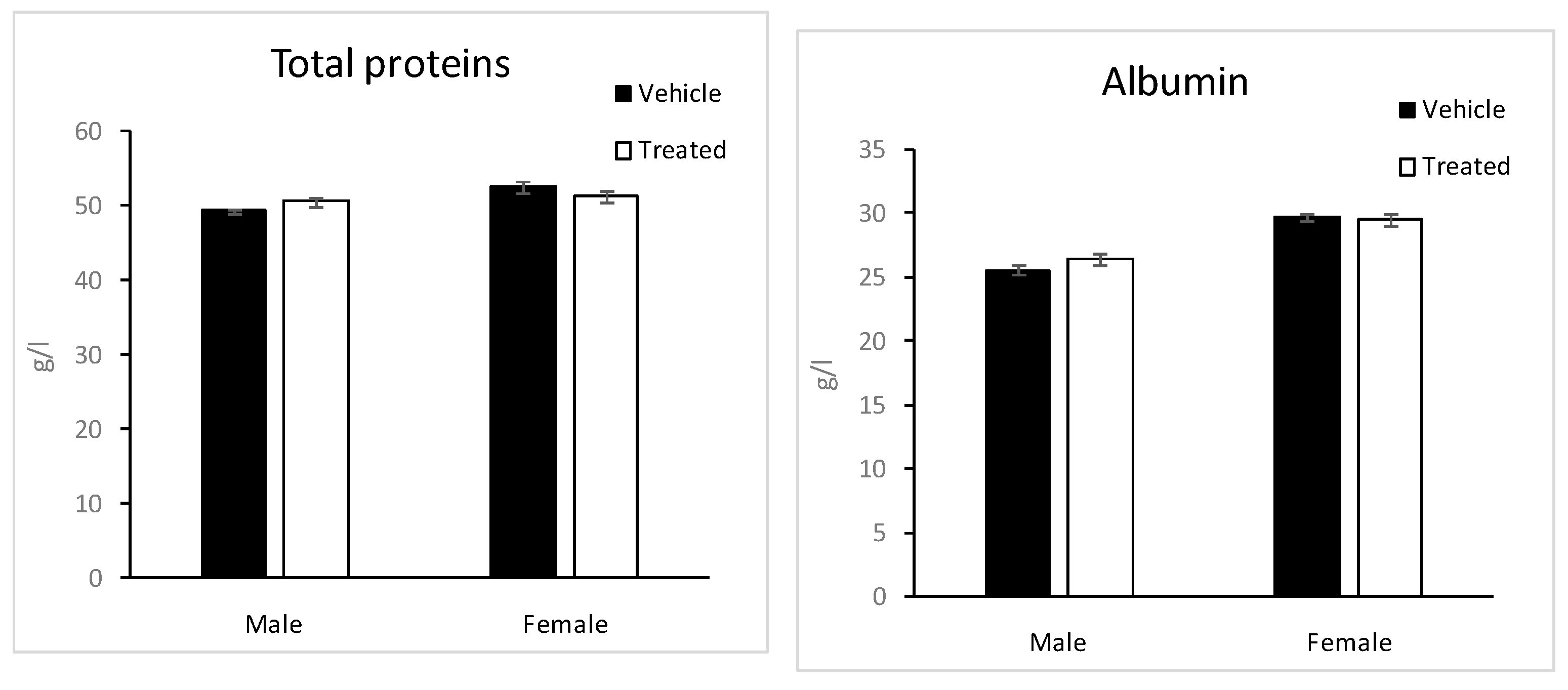
2.6. Biochemistry Parameters
2.7. Blood Hematology
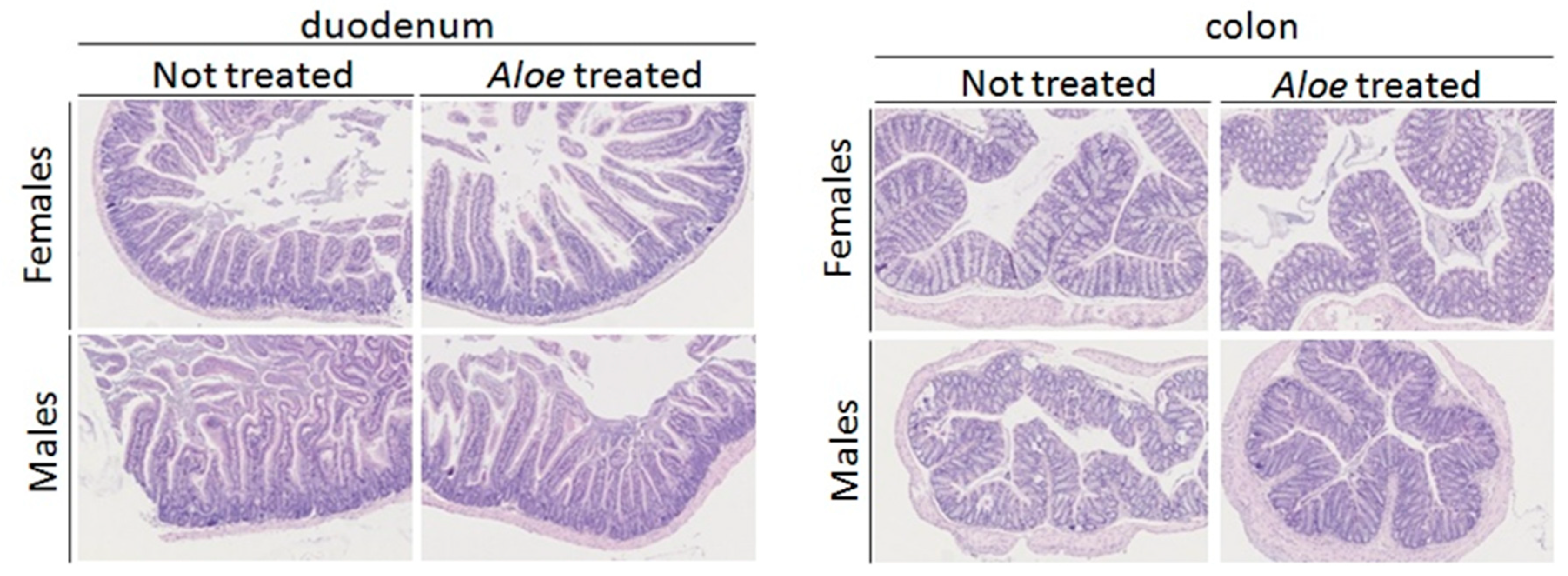
2.8. Histology
3. Materials and Methods
3.1. Plant Material and Extract Preparation
3.2. Chemicals
3.3. Antioxidant Activity Tests
3.3.1. DPPH Radical-Scavenging Test
3.3.2. ABTS Radical-Scavenging Test
3.3.3. FRAP Assay
3.4. Determination of Phenolic Content (PC) and Flavonoid Content (FC)
3.5. LC-MS/MS Analysis, Creation of Molecular Networking and NMR Apparatus
3.6. In-Vitro Toxicity
3.7. In Vivo Toxicity
3.7.1. Animal and Ethics
3.7.2. Study Design
3.7.3. Dysmorphological Screen
3.7.4. Body Temperature
3.7.5. Blood Analysis
3.7.6. Histology
3.7.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Salehi, B.; Albayrak, S.; Antolak, H.; Kręgiel, D.; Pawlikowska, E.; Sharifi-Rad, M.; Uprety, Y.; Tsouh Fokou, P.; Yousef, Z.; Amiruddin Zakaria, Z.; et al. Aloe Genus Plants: From Farm to Food Applications and Phytopharmacotherapy. IJMS 2018, 19, 2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Mei, N. Aloe vera: A Review of Toxicity and Adverse Clinical Effects. J. Environ. Sci. Health Part C 2016, 34, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Crosswhite, F.; Crosswhite, C. Aloe vera, Plant Symbolism and the Threshing Floor. Deserts Plants 1984, 6, 43–50. [Google Scholar]
- Eshun, K.; He, Q. Aloe vera: A Valuable Ingredient for the Food, Pharmaceutical and Cosmetic Industries—A Review. Crit. Rev. Food Sci. Nutr. 2004, 44, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Yadav, A.; Yadav, M.; Yadav, J.P. Effect of Climate Change on Phytochemical Diversity, Total Phenolic Content and in Vitro Antioxidant Activity of Aloe vera (L.) Burm.f. BMC Res. Notes 2017, 10, 60. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Singh, A.K.; Gupta, A.; Bishayee, A.; Pandey, A.K. Therapeutic Potential of Aloe vera—A Miracle Gift of Nature. Phytomedicine 2019, 60, 152996. [Google Scholar] [CrossRef]
- Ni, Y.; Turner, D.; Yates, K.M.; Tizard, I. Isolation and Characterization of Structural Components of Aloe vera L. Leaf Pulp. Int. Immunopharmacol. 2004, 4, 1745–1755. [Google Scholar] [CrossRef]
- El Sayed, A.M.; Ezzat, S.M.; El Naggar, M.M.; El Hawary, S.S. In Vivo Diabetic Wound Healing Effect and HPLC–DAD–ESI–MS/MS Profiling of the Methanol Extracts of Eight Aloe Species. Rev. Bras. de Farmacogn. 2016, 26, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Teradaira, R.; Shinzato, M.; Beppu, H.; Fujita, K. Antigastric Ulcer Effects in Rats OfAloe Arborescens Miller Var.Natalensis Berger Extract. Phytother. Res. 1993, 7, S34–S36. [Google Scholar] [CrossRef]
- Yagi, A.; Kanbara, T.; Morinobu, N. Inhibition of Mushroom-Tyrosinase by Aloe Extract. Planta Med. 1987, 53, 515–517. [Google Scholar] [CrossRef]
- Hutter, J.A.; Salman, M.; Stavinoha, W.B.; Satsangi, N.; Williams, R.F.; Streeper, R.T.; Weintraub, S.T. Antiinflammatory C-Glucosyl Chromone from Aloe Barbadensis. J. Nat. Prod. 1996, 59, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Grosse, Y.; Loomis, D.; Lauby-Secretan, B.; Ghissassi, F.E.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Baan, R.; Mattock, H.; Straif, K. Carcinogenicity of Some Drugs and Herbal Products. Lancet Oncol. 2013, 14, 807–808. [Google Scholar] [CrossRef]
- Shao, A.; Broadmeadow, A.; Goddard, G.; Bejar, E.; Frankos, V. Safety of Purified Decolorized (Low Anthraquinone) Whole Leaf Aloe vera (L) Burm. f. Juice in a 3-Month Drinking Water Toxicity Study in F344 Rats. Food Chem. Toxicol. 2013, 57, 21–31. [Google Scholar] [CrossRef]
- Mccoy, T. Aloe djiboutiensis and Aloe Ericahenriettae Two New Species from Djibouti: And the Mystery of Aloe Eumassawana’s Natural Habitat Solved. Cactus Succul. J. 2007, 79, 269–273. [Google Scholar] [CrossRef]
- Abdoul-Latif, F.; Merito, A.; Elmi, A. Plantes Médicinales de la Republique de Djibouti; Editions Universitaires Europeennes EUE: Chisinau, Republic of Moldova, 2018; Volume 1. [Google Scholar]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frum, Y.; Viljoen, A.M. In Vitro 5-Lipoxygenase and Anti-Oxidant Activities of South African Medicinal Plants Commonly Used Topically for Skin Diseases. Skin Pharmacol. Physiol. 2006, 19, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sazhina, N.N.; Lapshin, P.V.; Zagoskina, N.V.; Misin, V.M. Comparative Study of Antioxidant Properties of Extracts of Various Aloe Species. Russ. J. Bioorg. Chem. 2016, 42, 735–740. [Google Scholar] [CrossRef]
- López, A.; de Tangil, M.; Vega-Orellana, O.; Ramírez, A.; Rico, M. Phenolic Constituents, Antioxidant and Preliminary Antimycoplasmic Activities of Leaf Skin and Flowers of Aloe vera (L.) Burm. f. (Syn. A. Barbadensis Mill.) from the Canary Islands (Spain). Molecules 2013, 18, 4942–4954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, D.K.; Saini, M.R. Evaluation of Radioprotective Efficacy and Possible Mechanism of Action of Aloe Gel. Environ. Toxicol. Pharmacol. 2011, 31, 427–435. [Google Scholar] [CrossRef]
- Giannakoudakis, D.A.; Hosseini-Bandegharaei, A.; Tsafrakidou, P.; Triantafyllidis, K.S.; Kornaros, M.; Anastopoulos, I. Aloe vera Waste Biomass-Based Adsorbents for the Removal of Aquatic Pollutants: A Review. J. Environ. Manag. 2018, 227, 354–364. [Google Scholar] [CrossRef]
- Yen, G. Antioxidant Activity of Anthraquinones and Anthrone. Food Chem. 2000, 70, 437–441. [Google Scholar] [CrossRef]
- Hęś, M.; Dziedzic, K.; Górecka, D.; Jędrusek-Golińska, A.; Gujska, E. Aloe vera (L.) Webb.: Natural Sources of Antioxidants—A Review. Plant Foods Hum. Nutr. 2019, 74, 255–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Grace, O.M.; Kokubun, T.; Veitch, N.C.; Simmonds, M.S.J. Characterisation of a Nataloin Derivative from Aloe Ellenbeckii, a Maculate Species from East Africa. S. Afr. J. Bot. 2008, 74, 761–763. [Google Scholar] [CrossRef]
- Abouelela, M.E.; Assaf, H.K.; Abdelhamid, R.A.; Elkhyat, E.S.; Sayed, A.M.; Oszako, T.; Belbahri, L.; El Zowalaty, A.E.; Abdelkader, M.S.A. Identification of Potential SARS-CoV-2 Main Protease and Spike Protein Inhibitors from the Genus Aloe: An In Silico Study for Drug Development. Molecules 2021, 26, 1767. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Parshad, B.; Manchanda, P.; Sharma, S.K. Chromones and Their Derivatives as Radical Scavengers: A Remedy for Cell Impairment. Curr. Top. Med. Chem. 2014, 14, 2552–2575. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Hua, Y. Concentration-Dependence of Prooxidant and Antioxidant Effects of Aloin and Aloe-Emodin on DNA. Food Chem. 2005, 91, 413–418. [Google Scholar] [CrossRef]
- Cock, I. Problems of Reproducibility and Efficacy of Bioassays Using Crude Extracts, with Reference to Aloe vera. Pharmacogn. Commun. 2011, 1, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Teo, S.; Stirling, D.; Thomas, S.; Hoberman, A.; Kiorpes, A.; Khetani, V. A 90-Day Oral Gavage Toxicity Study of d-Methylphenidate and d,l-Methylphenidate in Sprague–Dawley Rats. Toxicology 2002, 179, 183–196. [Google Scholar] [CrossRef]
- Teka, T.; Bisrat, D.; Yeshak, M.; Asres, K. Antimalarial Activity of the Chemical Constituents of the Leaf Latex of Aloe Pulcherrima Gilbert and Sebsebe. Molecules 2016, 21, 1415. [Google Scholar] [CrossRef]
- Wang, B.; Feng, W.; Wang, M.; Wang, T.; Gu, Y.; Zhu, M.; Ouyang, H.; Shi, J.; Zhang, F.; Zhao, Y.; et al. Acute Toxicological Impact of Nano- and Submicro-Scaled Zinc Oxide Powder on Healthy Adult Mice. J. Nanopart. Res. 2008, 10, 263–276. [Google Scholar] [CrossRef]
- Loots, D.T.; van der Westhuizen, F.H.; Botes, L. Aloe Ferox Leaf Gel Phytochemical Content, Antioxidant Capacity, and Possible Health Benefits. J. Agric. Food Chem. 2007, 55, 6891–6896. [Google Scholar] [CrossRef] [PubMed]
- Yakubu, M.T.; Akanji, M.A.; Oladiji, A.T. Alterations in Serum Lipid Profile of Male Rats by Oral Administration of Aqueous Extract of Fadogia Agrestis Stem. Res. J. Med. Plant 2008, 2, 66–73. [Google Scholar]
- Nair, A.; Jacob, S. A Simple Practice Guide for Dose Conversion between Animals and Human. J. Basic Clin. Pharma 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Ban, X.; He, J.; Tong, J.; Tian, J.; Wang, Y. Hepatoprotective and Antioxidant Activity of Ethanolic Extracts of Edible Lotus (Nelumbo Nucifera Gaertn.) Leaves. Food Chem. 2010, 120, 873–878. [Google Scholar] [CrossRef]
- Elmi, A.; Spina, R.; Risler, A.; Philippot, S.; Mérito, A.; Duval, R.E.; Abdoul-latif, F.M.; Laurain-Mattar, D. Evaluation of Antioxidant and Antibacterial Activities, Cytotoxicity of Acacia Seyal Del Bark Extracts and Isolated Compounds. Molecules 2020, 25, 2392. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Ke, H.; He, J.; Ban, X.; Zeng, H.; Wang, Y. Extracts of Halenia Elliptica Exhibit Antioxidant Properties in Vitro and in Vivo. Food Chem. Toxicol. 2011, 49, 185–190. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmi, A.; Spina, R.; Abdoul-Latif, F.; Yagi, S.; Fontanay, S.; Risler, A.; Duval, R.E.; Laurain-Mattar, D. Rapid Screening for Bioactive Natural Compounds in Indigofera Caerulea Rox Fruits. Ind. Crop. Prod. 2018, 125, 123–130. [Google Scholar] [CrossRef]
- Mohimani, H.; Gurevich, A.; Shlemov, A.; Mikheenko, A.; Korobeynikov, A.; Cao, L.; Shcherbin, E.; Nothias, L.-F.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of Microbial Metabolites through Database Search of Mass Spectra. Nat. Commun. 2018, 9, 4035. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Lasarow, R.M.; Isseroff, R.R.; Gomez, E.C. Quantitative In Vitro Assessment of Phototoxicity by a Fibroblast-Neutral Red Assay. J. Investig. Dermatol. 1992, 98, 725–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chia, R.; Achilli, F.; Festing, M.F.W.; Fisher, E.M.C. The Origins and Uses of Mouse Outbred Stocks. Nat. Genet. 2005, 37, 1181–1186. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, H.; Schughart, K.; Wolf, E.; Balling, R.; de Angelis, M.H. Screening for Dysmorphological Abnormalities-a Powerful Tool to Isolate New Mouse Mutants. Mamm. Genome 2000, 11, 528–530. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antioxidant Activity | Phenolic Content (PC) | Flavonoid Content (FC) | |||
|---|---|---|---|---|---|
| Sample | DPPH; ABTS IC50 (µg/mL) | FRAP (mg EVC/g DW) | mg GAE/g DW | mg QE/g DW | |
| ADL | >1000 | 600 ± 1.8 | 1.50 ± 0.12 | 83.01 ± 0.8 | 13.12 ± 0.11 |
| ADM | >1000 | 632 ± 3.2 | 1.68 ± 0.20 | 46.4 ± 0.32 | 4.25 ± 0.1 |
| Vitamin C | 220 ± 3.1 | 50 ± 3.3 | - | - | - |
| Trolox | 130 ± 7.1 | 50 ± 2.7 | - | - | - |
| Compound Label | Name | Retention Time in Minutes | Chemical Formula [M + H]+ | Mesured m/z (Da) [M + H]+ | Error (ppm) | MS/MS Fragmentation | Cluster Number | Presence in the Samples |
|---|---|---|---|---|---|---|---|---|
| Anthraquinones Derivatives | ||||||||
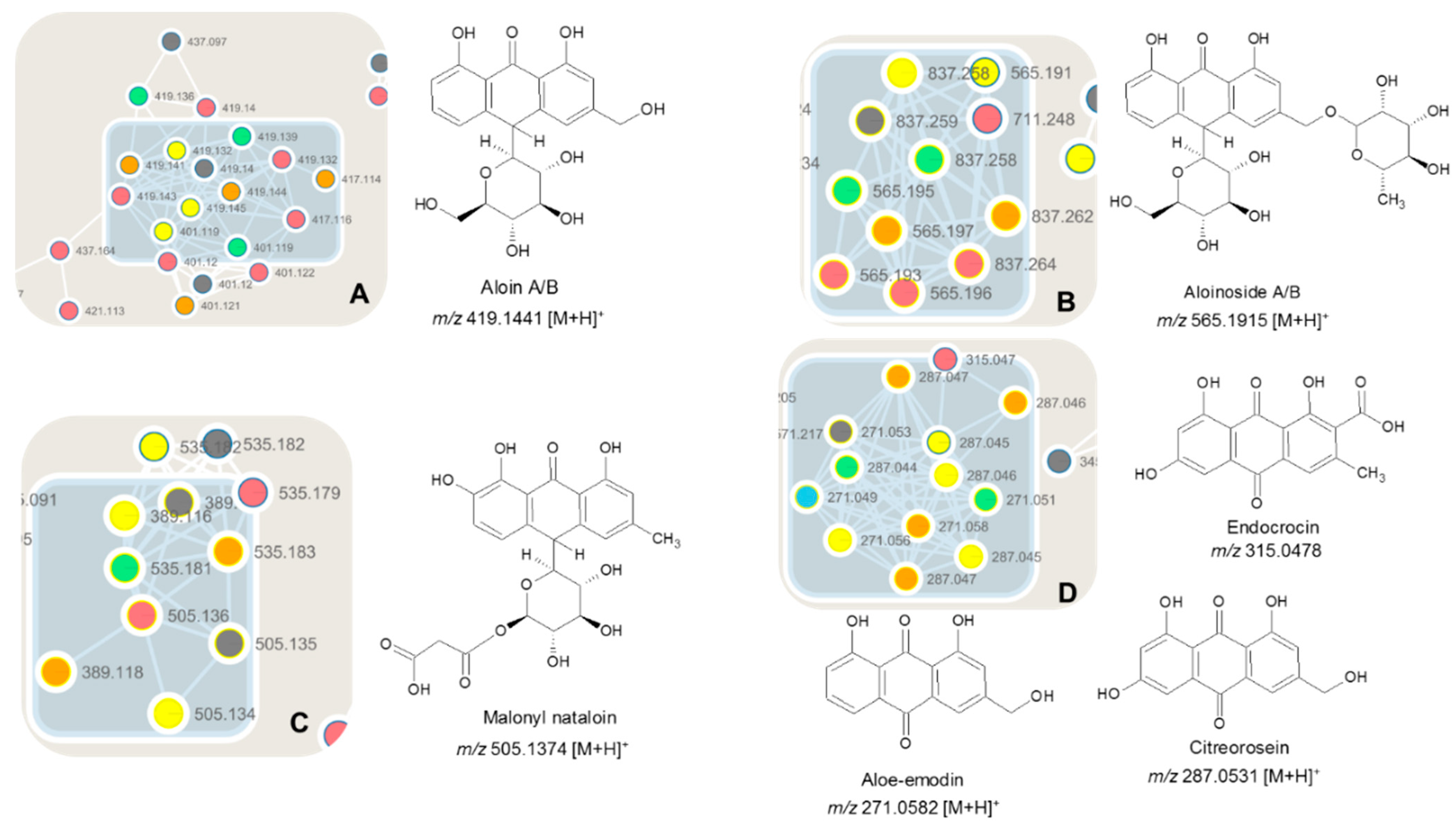
| Cmpds 1A and 1B DT | Aloin A/B | 14.7/15.1 | [C21H20O9 + H]+ | 419.1441 | 0.4 | 257.0796 [M-glycosyl + H]+ 239.0768 [M-glycosyl-H2O + H]+ 211.0785 [M-glycosyl-H2O-CO + H]+ | A | ADL, FR-II FR-III, FR-IV, FR-V |
| Cmpds 2A and 2B D+ | Aloinoside A/B | 15.0/15.7 | [C27H32O13 + H]+ | 565.1915 | 6.4 | 419.1416 [M-rhamnosyl + H]+ 239.0730 [M-rhamnosyl-glycosyl + H]+ 211.0737 [M-rhamnosyl-glycosyl-CO + H]+ | B | ADL FR-II, FR-III |
| Cmpd 3 M | Malonyl nataloin | 15.6 | [C24H24O12 + H] | 505.1374 | 6.6 | 239.0697 [M-mal-glycosyl + H]+ | C | ADL FR-IV, FR-V |
| Cmpd 4 | Aloin-pentose | 16.9 | [C20H20O8 + H]+ | 389.1213 | 4.6 | 239.0676 [M-pentosyl-H2O + H]+ | C | ADL FR-II, FR-III FR-IV, FR-V |
| Cmpd 5 | Endocrocin | 13.4 | [C16H10O7 + H]+ | 315.0478 | 6.9 | 297.0369 [M-H2O + H]+ 269.0442 [M-H2O-CO2 + H]+ 241.048 [M-H2O-2CO2 + H]+ | D | ADL FR-I, FR-II, FR-III, FR-IV, FR-V |
| Cmpd 6 DT | Aloemodin | 19.1 | [C15H10O5 + H]+ | 271.0582 | 7.0 | 253.0477 [M-H2O + H]+ 241.0465 [M-CH2O + H]+ 225.0526 [M-H2O-CO + H]+ | D | ADL FR-I, FR-II, FR-III, FR-IV, FR-V |
| Cmpd 7 | Citreorosein | 17.4 | [C15H10O6 + H]+ | 287.0531 | 6.7 | 269.0454 [M-H2O + H]+ 241.0487 [M-H2O-CO + H]+ 213.0537 [M-H2O-2CO + H]+ 185.0578 [M-H2O-3CO + H]+ 157.0654 [M-H2O-4CO + H]+ | D | ADL FR-IV, FR-V |
| Chromones Derivatives | ||||||||
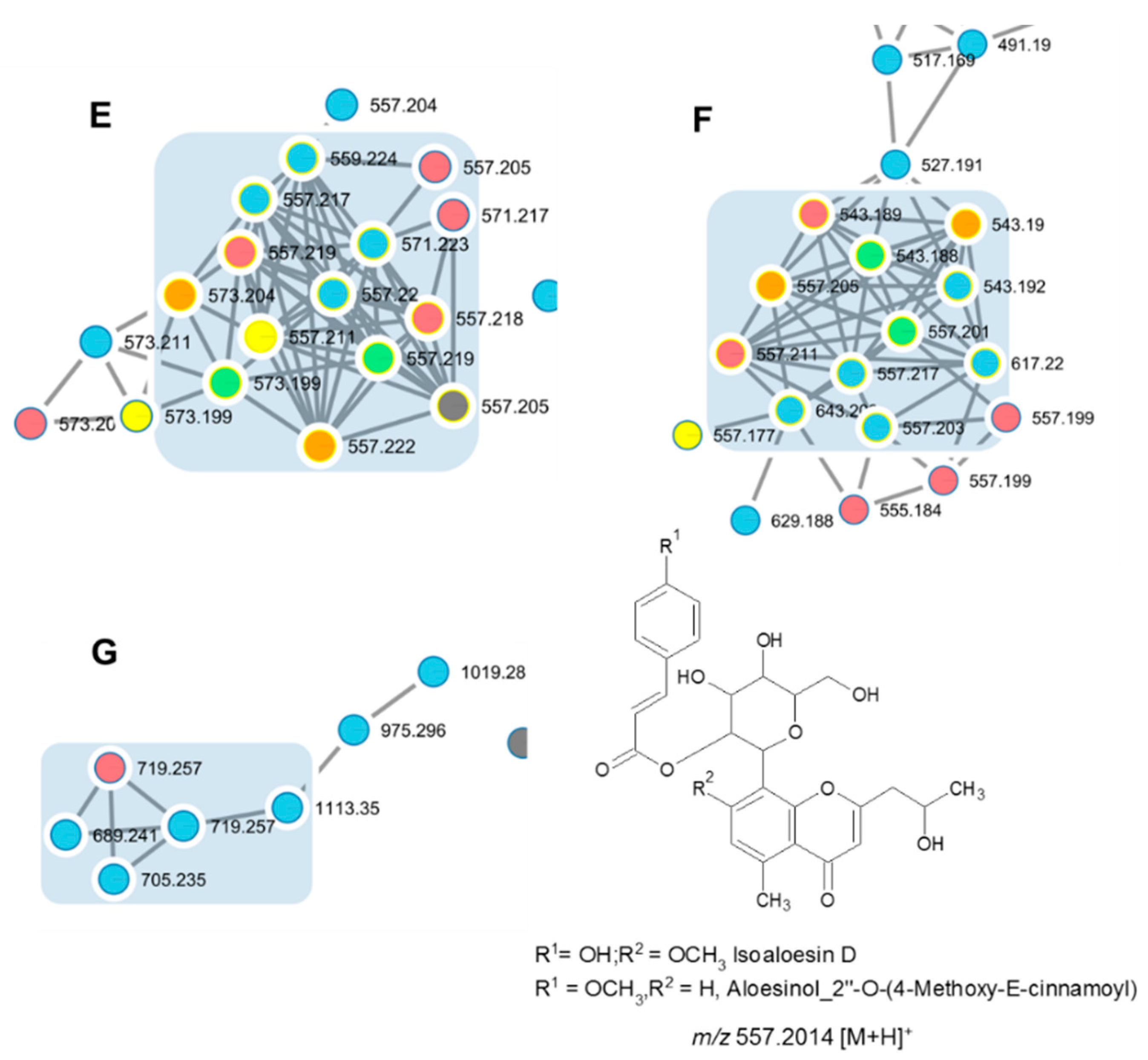
| Cmpd 8 D+ | Isoaloeresin D or Aloesinol_7-Me_ether,_2″-O-(4-hydroxy-E-cinnamoyl) | 15.4 | [C29H32O11 + H]+ | 557.1998 | 9.5 | 513.1783 [M-CH2CHOH + H]+ 349.1291 [M-CH2CHOH-coumaric + H]+ 217.0876 [M-CH2CHOH-coumaric-pentosyl + H]+ 187.0727 [M-CH2CHOH-coumaric-glycosyl + H]+ 437.1589 [M-part of sugar + H]+ 393.1374 [M-part of sugar-CH2CO + H]+ 247.0963 [M-part of sugar-CH2CO-(coumaric-H2O) + H]+ 147.0485 [coumaric-H2O + H]+ | E | ADL FR-I, FR-II; FR-III, FR-IV, FR-V |
| Cmpds 9A and 9B D+ | Aloesinol_2″-O-(4-Methoxy-cinnamoyl) | 16.0/17.0 | [C29H32O11 + H]+ | 557.2014 | 0.7 | 513.1783 [M-CH2CHOH + H]+ 335.1101 [M-CH2CHOH-MeO-cinnamoyl + H]+ 203.0672 [M-CH2CHOH-MeO-cinnamoyl-pentosyl + H]+ 161.059 (MeO-cinnamoyl)+ | F | FR-I |
| Cmpds 10A and 10B D+ | Isoloeresin-D + glycosyl | 12.6/13.2 | [C35H42O16 + H]+ | 719.2570 | 4.5 | 557.2033 [M-glycosyl + H]+ 393.1326 [M-2 glycosyl + H]+ 247.0962 [M-2 glycosyl-(coumaric-H2O) + H]+ 147.0427 [Coumaric acid-H2O + H]+ | G | ADL FR-I |
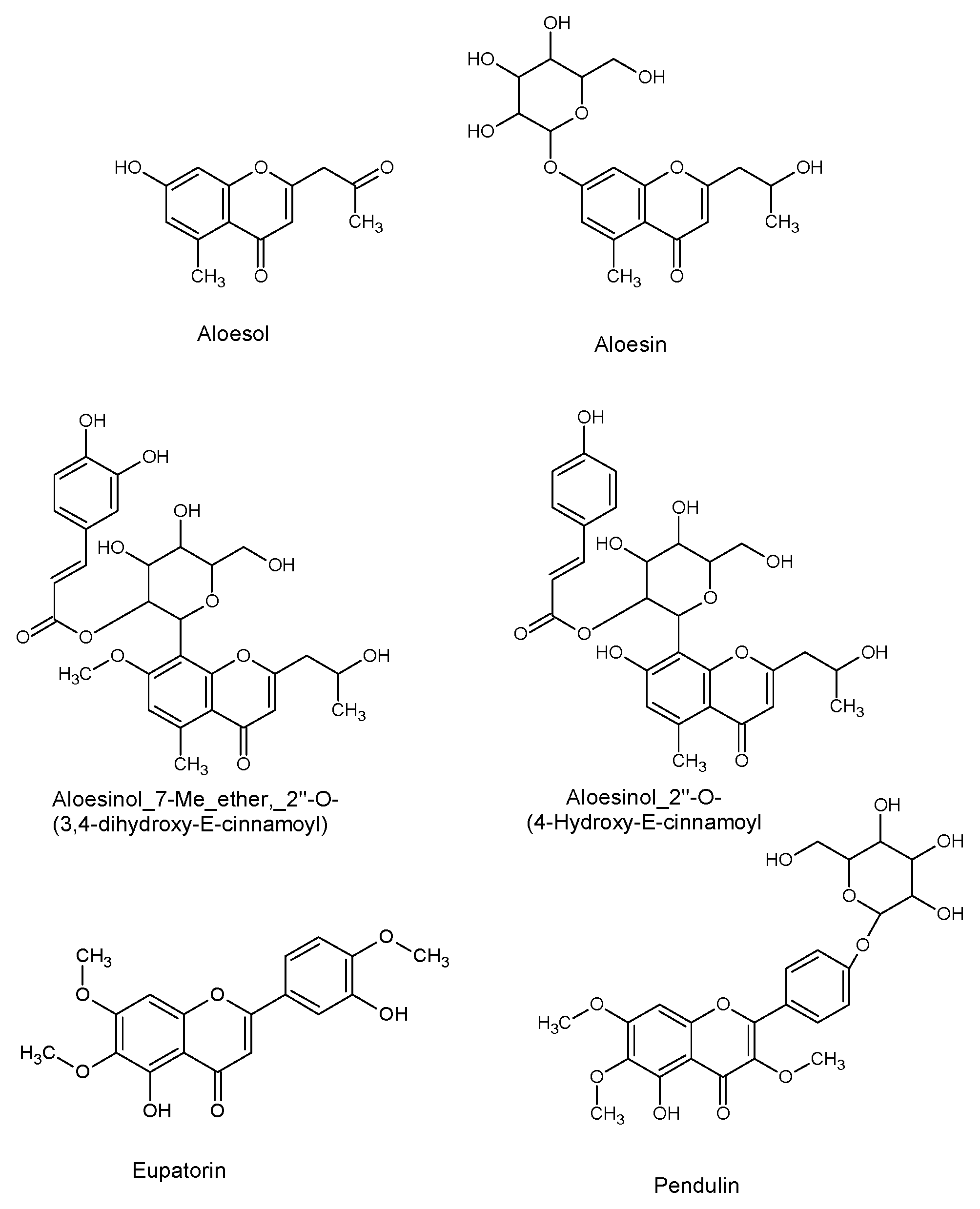
| Cmpd 11 | Aloesol | 13.5 | [C13H14O4 + H]+ | 235.0951 | 6.9 | 191.0684 [M-C2H4O + H]+ 176.0466 [M-C2H4O-CH3 + H]+ | No cluster | FR-III |
| Cmpd 12 | Aloesin | 12.4 | [C19H22O9 + H]+ | 395.1339 | 0.6 | 233.0786 [M-glycosyl + H]+ 215.068 [M-glycosyl-H2O + H]+ 203.0666 [M-glycosyl-CH2O + H]+ | H | ADL |
| Cmpd 13 | Aloesin–Glycoside | 9.6 | [C25H32O14 + H]+ | 557.1874 | 1.6 | 395.1321 [M-glycosyl + H]+ 233.0780 [M-2 glycosyl + H]+ | H | ADL FR-I |
| Cmpd 14 D+ | Aloesinol_7-Me_ether,_2″-O-(3,4-dihydroxy-E-cinnamoyl) | 13.8 | [C29H32O12 + H]+ | 573.1982 | 2.8 | 529.1748 [M-CH2CHOH + H]+ 367.1345 [M-CH2CHOH-(caffeic-H2O) + H]+ 205.0817 [M-CH2CHOH-(caffeic-H2O) + H]+ 409.1270 [M-deoxyhexose + H]+ 247.0948 [M-deoxyhexose-(caffeic-H2O) + H]+ 163.0378 [caffeic-H2O + H]+ | E | ADL FR-I, FR-II, FR-III, FR-IV, FR-V |
| Cmpd 15 D+ | Aloesinol_2″-O-(4-Hydroxy-E-cinnamoyl) | 13.4 | [C28H30O11 + H]+ | 543.1908 | 8.6 | 499.1630 [M-CH2CHOH + H]+ 335.1095 [M-CH2CHOH-coumaric + H]+ 203.0659 [M-CH2CHOH-coumaric-pentosyl + H]+ 397.1467 [M-(coumaric-H2O) + H]+ 233.0781 [M-(coumaric-H2O)-deoxyhexose + H]+ 379.1258 [M-deoxyhexose + H]+ 147.0485 [coumaric-H2O + H]+ | I | ADL FR-I, FR-II, FR-III |
| Flavonols/Chromones Isomers | ||||||||
| Cmpd 16 | Eupatorin or aglycon of pendulin | 16.4 | [C18H16O7 + H]+ | 345.0953 | 4.5 | 285.0768 [M-H2O-CH2CO + H]+ 267.0615 [M-H2O-CH2CO-H2O + H]+ | L | ADL FR-IV, FR-V |
| Cmpd 17 | Eupatorin-glycosyde or Pendulin | 13.6 | [C24H26O12 + H]+ | 507.1505 | 1.5 | 345.0959 [M-glycosyl + H]+ 327.0858 [M-glycosyl-H2O + H]+ 285.0768 [M-glycosyl-H2O-CH2CO + H]+ | M | ADL FR-I, FR-II, FR-III, FR-IV, FR-V |
| Parameters | Dosage (g/kg/day) | ||||
|---|---|---|---|---|---|
| Male | Female | ||||
| Unit | 0 | 0.15 | 0 | 0.15 | |
| WBC | ×103 cells/µL | 5.60 | 7.07 | 6.54 | 5.79 |
| RBC | ×106 cells/µL | 8.65 | 9.01 | 9.35 | 9.56 |
| HGB | g/dL | 12.8 | 13.4 | 14.4 | 14.1 |
| HCT | % | 45.6 | 47.6 | 49.8 | 49.0 |
| MCV ** | fL | 52.8 | 52.9 | 53.3 | 51.3 |
| MCH * | pg | 14.8 | 14.9 | 15.4 | 14.8 |
| MCHC | g/dL | 28.0 | 28.2 | 28.9 | 28.9 |
| NEUTRO | % | 17.6 | 19.8 | 13.4 | 18.8 |
| LYMPHO | % | 76.2 | 74.0 | 80.0 | 74.6 |
| MONO | % | 3.1 | 2.1 | 1.5 | 1.9 |
| EOSINO | % | 2.6 | 3.6 | 4.7 | 4.5 |
| LUC | % | 0.4 | 0.4 | 0.3 | 0.1 |
| BASO | % | 0.1 | 0.1 | 0.1 | 0.1 |
| PLT | ×103 cells/µL | 1232 | 1307 | 1090 | 1236 |
| MPV | fL | 4.2 | 4.3 | 4.5 | 4.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmi, A.; Mohamed Abdoul-Latif, F.; Spina, R.; Dupire, F.; Philippot, S.; Marie-France, C.; Jacobs, H.; Laurain-Mattar, D. Aloe djiboutiensis: Antioxidant Activity, Molecular Networking-Based Approach and In Vivo Toxicity of This Endemic Species in Djibouti. Molecules 2021, 26, 3046. https://doi.org/10.3390/molecules26103046
Elmi A, Mohamed Abdoul-Latif F, Spina R, Dupire F, Philippot S, Marie-France C, Jacobs H, Laurain-Mattar D. Aloe djiboutiensis: Antioxidant Activity, Molecular Networking-Based Approach and In Vivo Toxicity of This Endemic Species in Djibouti. Molecules. 2021; 26(10):3046. https://doi.org/10.3390/molecules26103046
Chicago/Turabian StyleElmi, Abdirahman, Fatouma Mohamed Abdoul-Latif, Rosella Spina, François Dupire, Stéphanie Philippot, Champy Marie-France, Hugues Jacobs, and Dominique Laurain-Mattar. 2021. "Aloe djiboutiensis: Antioxidant Activity, Molecular Networking-Based Approach and In Vivo Toxicity of This Endemic Species in Djibouti" Molecules 26, no. 10: 3046. https://doi.org/10.3390/molecules26103046
APA StyleElmi, A., Mohamed Abdoul-Latif, F., Spina, R., Dupire, F., Philippot, S., Marie-France, C., Jacobs, H., & Laurain-Mattar, D. (2021). Aloe djiboutiensis: Antioxidant Activity, Molecular Networking-Based Approach and In Vivo Toxicity of This Endemic Species in Djibouti. Molecules, 26(10), 3046. https://doi.org/10.3390/molecules26103046