
Piperlongumine Analogs Promote A549 Cell Apoptosis through Enhancing ROS Generation


Abstract
:1. Introduction
2. Results
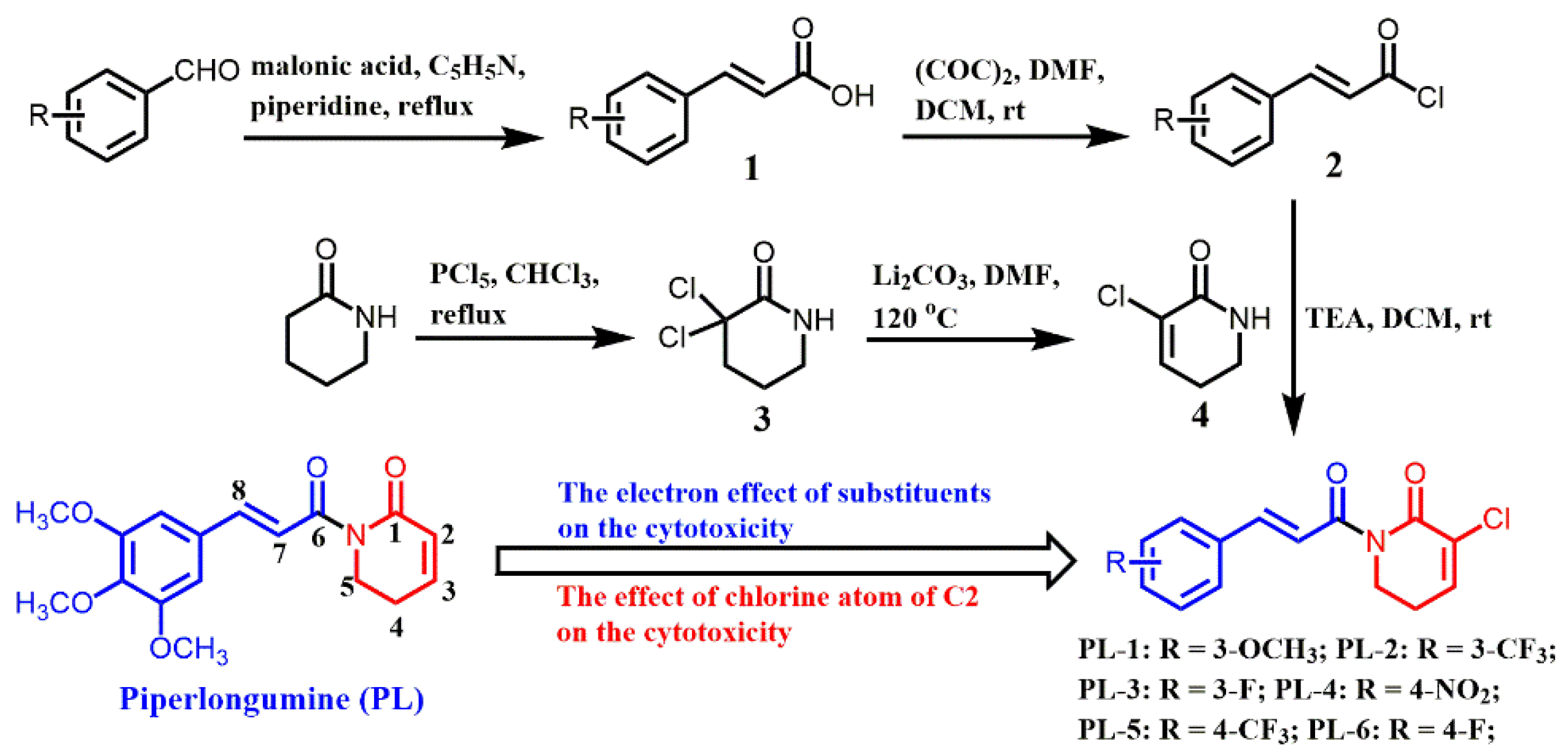
2.1. Chemistry
2.2. Assessment of In Vitro Cytotoxicity
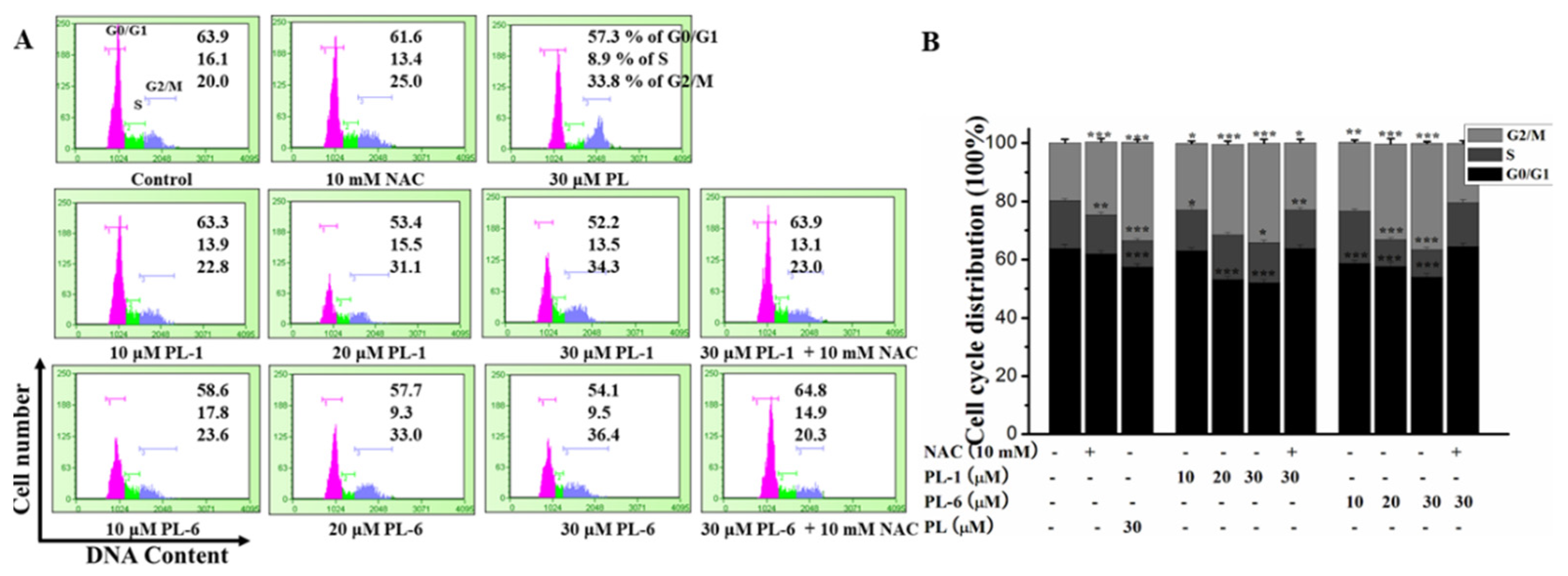
2.3. Piperlongumine (PL) Analogs Triggered Cell Cycle Arrest
2.4. The Apoptotic Effects of PL Analogs against the A549 Cell
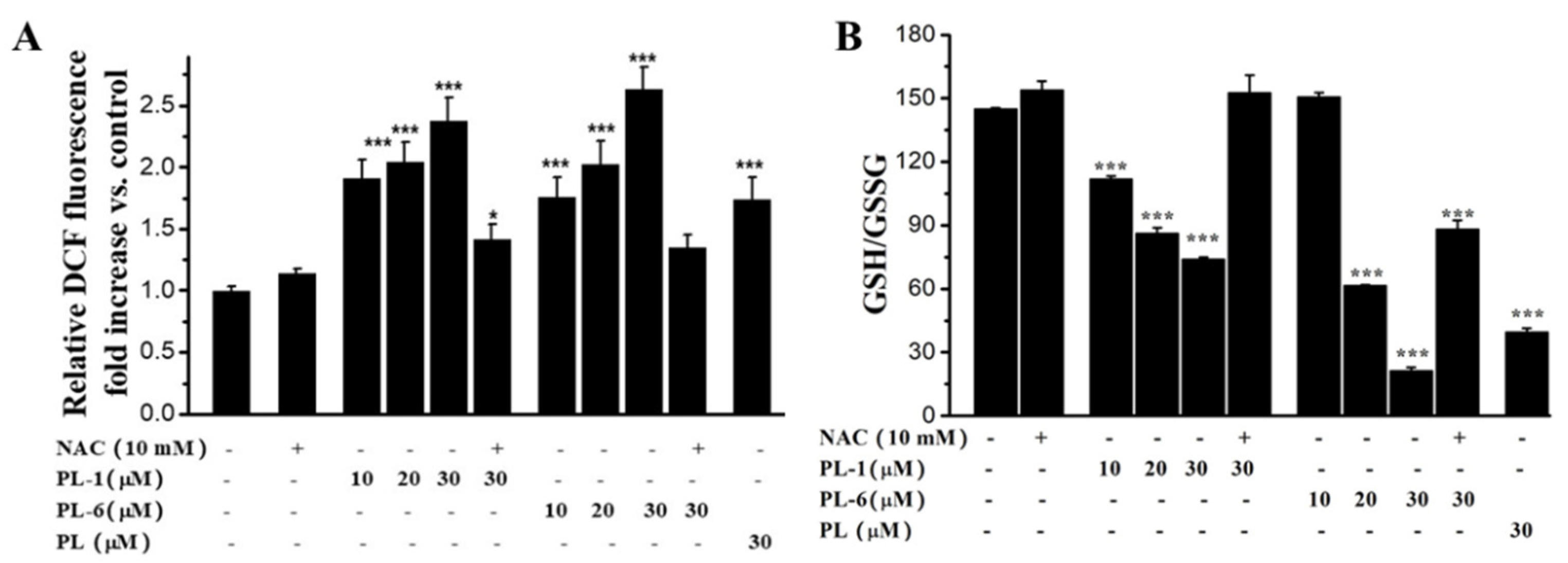
2.5. PL Analogs Induced Reactive Oxygen Species (ROS) Generation and Redox Imbalance
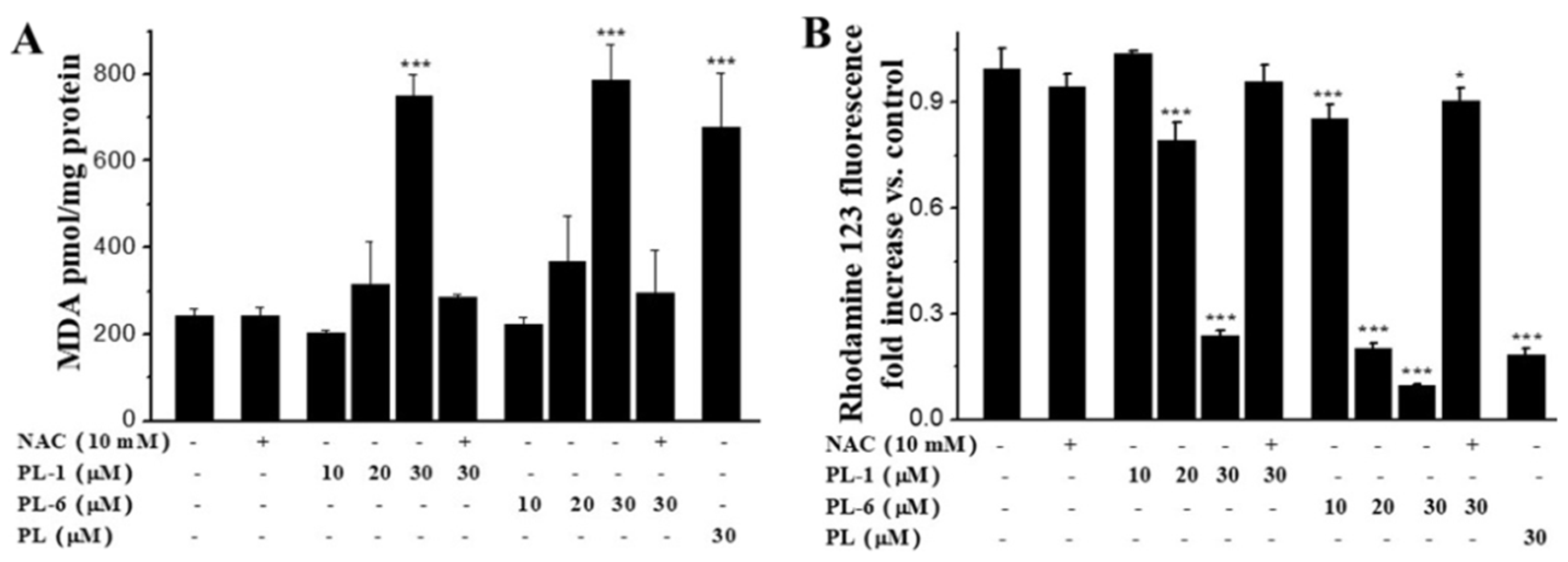
2.6. PL Analogs Induced Lipid Peroxidation and the Loss of MMP (Mitochondrial Membrane Potential)
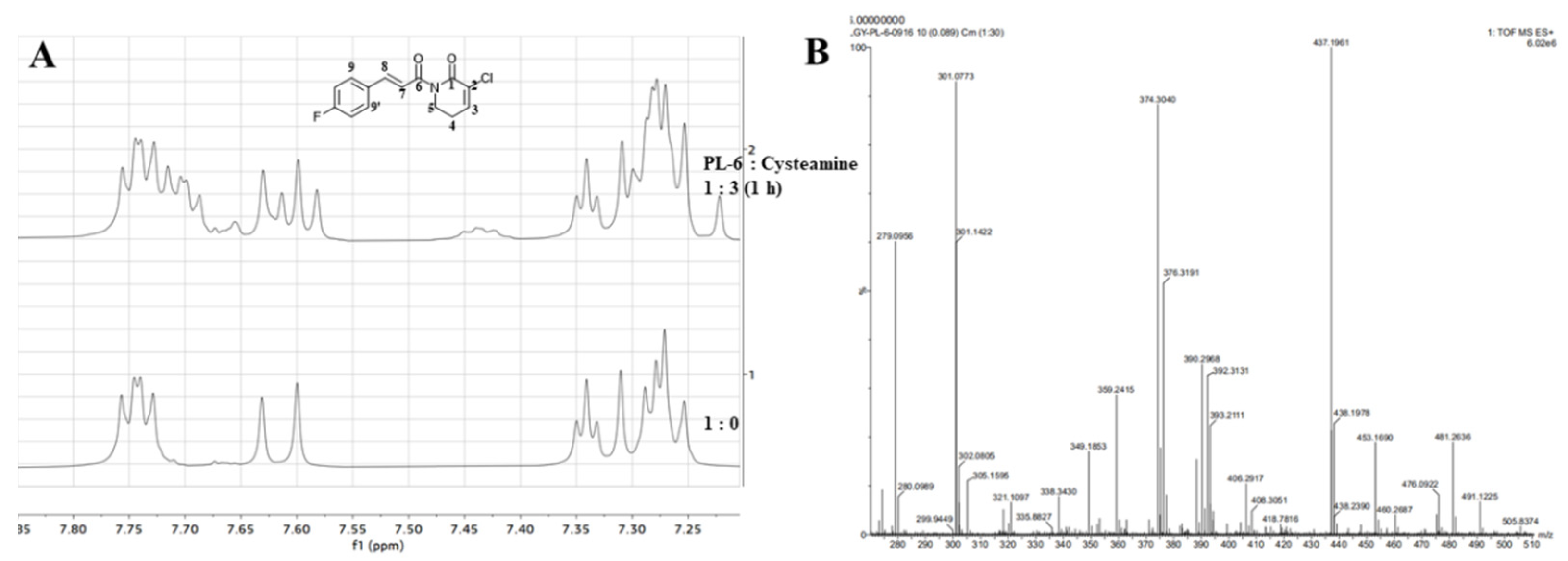
2.7. The Nucleophilic Addition Reaction of PL Analogs and Cysteamine
2.8. The Inhibition Activity of PL-6 on TrxR
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Synthesis of the PL Analogs
4.2.1. The Preparation of Cinnamoyl Chloride (Intermediate 2)
4.2.2. The Preparation of 3-Chloro-5,6-dihydropyridin-2(1H)-one (Intermediate 4)
4.3. Cytotoxicity Assay
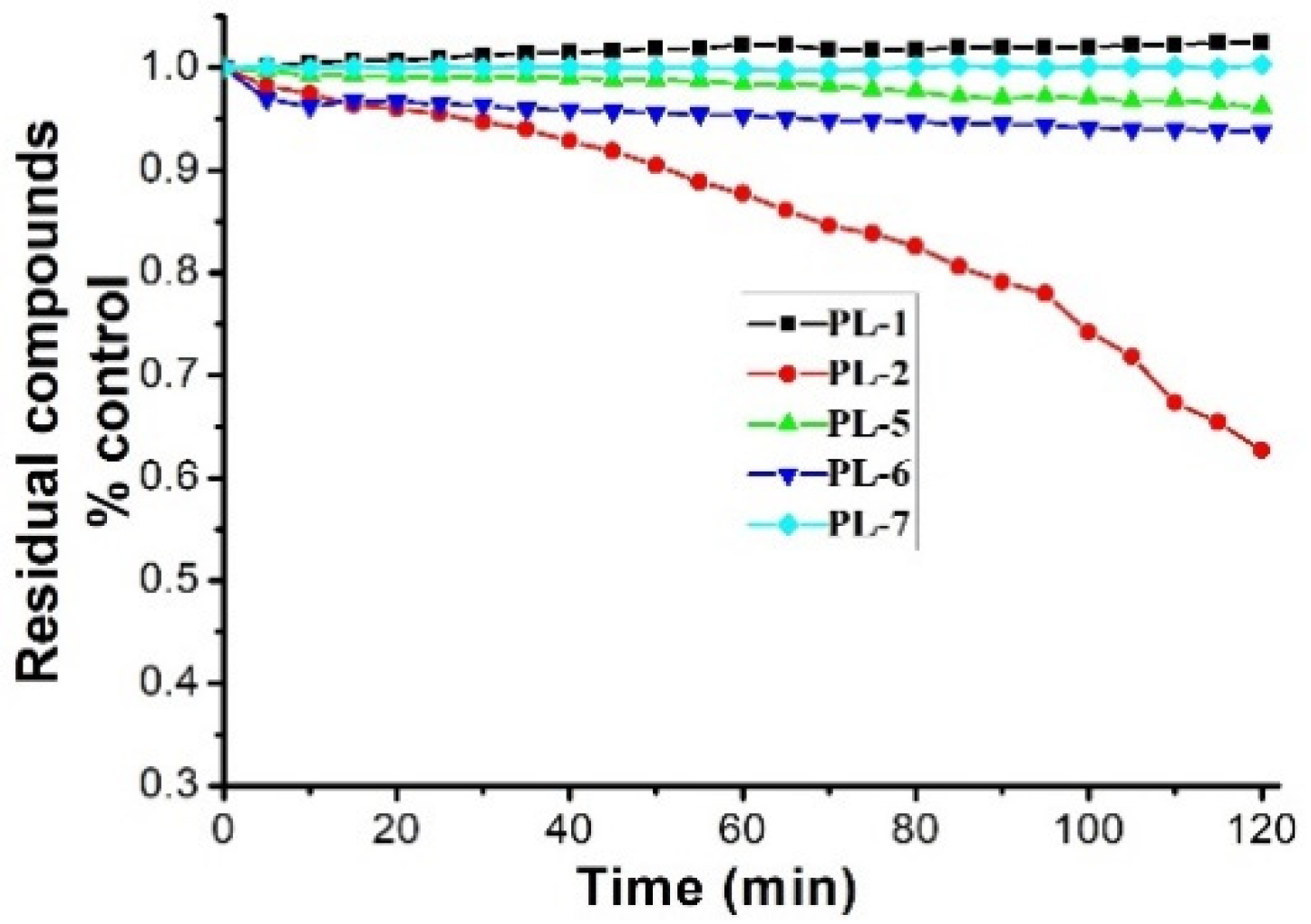
4.4. Stability Test
4.5. Cell Cycle Analysis
4.6. Cancer Cells Apoptosis Assay
4.7. ROS Detection
4.8. The Evaluation of Intracellular Redox Balance Levels
4.9. Determination of Lipid Peroxidation
4.10. Measurement of Mitochondrial Membrane Potential
4.11. Electrophilicity Assessment by a Kinetic Thiol Assay
4.12. Determination of the Reaction Site by Nuclear Magnetic Resonance (1H-NMR) and High-Resolution Mass Spectrometry (HRMS)
4.13. Western Blot Analysis
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Burgos, R.A.; Seguel, K.; Perez, M.; Meneses, A.; Ortega, M.; Guarda, M.I.; Loaiza, A.; Hancke, J.L. Andrographolide inhibits IFN-γ and IL-2 cytokine production and protects against cell apoptosis. Planta Med. 2005, 71, 429–434. [Google Scholar] [CrossRef]
- Fleck, J.D.; Betti, A.H.; Silva, F.P.D.; Troian, E.A.; Olivaro, C.; Ferreira, F.; Verza, S.G. Saponins from Quillaja saponaria and Quillaja brasiliensis: Particular Chemical Characteristics and Biological Activities. Molecules 2019, 24, 171. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Cao, Y.X.; Li, G.Y.; Lei, H.; Attiogbe, M.K.I.; Yao, J.C.; Yang, X.Y.; Liu, Y.J.; Hei, Y.Y.; Zhang, H. F10, a new camptothecin derivative, was identified as a new orally-bioavailable, potent antitumor agent. Eur. J. Med. Chem. 2020, 202, 112528. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Ren, J.; Wang, Y.; Su, J.Y.; Zhu, Y.M.; Chen, C.G.; Long, W.G.; Jiang, Q.; Li, J. Synergistic killing effect of paclitaxel and honokiol in non-small cell lung cancer cells through paraptosis induction. Cell. Oncol. 2020, 44, 135–150. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Krishnan, A.; Vohora, D. A systematic review on Piper longum L.: Bridging traditional knowledge and pharmacological evidence for future translational research. J. Ethnopharmacol. 2020, 247, 112255. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Qu, P.C.; Wang, Q.S.; Zhang, L.; Liu, H.L.; Luo, R.; Chen, X.Q.; Ba, Y.Y.; Wu, X.; Yang, H. Neuroprotective effects of alkaloids from Piper longum in a MPTP induced mouse model of Parkinson’s disease. Pharm. Biol. 2015, 53, 1516–1524. [Google Scholar] [CrossRef]
- Sucher, N.J.; Carles, M.C. A pharmacological basis of herbal medicines for epilepsy. Epilepsy Behav. 2015, 52, 308–318. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, H.; Corbet, C.; Mey, S.; Law, K.; Gevaert, T.; Feron, O.; Ridder, M.D. Piperlongumine increases sensitivity of colorectal cancer cells to radiation: Involvement of ROS production via dual inhibition of glutathione andthioredoxin systems. Cancer Lett. 2019, 450, 42–52. [Google Scholar] [CrossRef]
- Zhou, J.F.; Huang, Z.X.; Ni, X.; Lv, C. Piperlongumine induces apoptosis and G2/M phase arrest in human osteosarcoma cells by regulating ROS/PI3K/Akt pathway. Toxicol. In Vitro 2020, 65, 104775. [Google Scholar] [CrossRef]
- Zheng, J.; Son, D.J.; Gu, S.M.; Woo, J.R.; Ham, Y.W.; Lee, H.P.; Kim, W.J.; Jung, J.K.; Hong, J.T. Piperlongumine inhibits lung tumor growth via inhibition of nuclearfactor kappa B signaling pathway. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef]
- Thongsom, S.; Suginta, W.; Lee, K.J.; Choe, H.; Talabnin, C. Piperlongumine induces G2/M phase arrest and apoptosis in cholangiocarcinoma cells through the ROS-JNK-ERK signaling pathway. Apoptosis 2017, 22, 1473–1484. [Google Scholar] [CrossRef]
- Raj, L.; Ide, T.; Gurkar, A.U.; Foley, M.; Schenone, M.; Li, X.; Tolliday, N.J.; Golub, T.R.; Carr, S.A.; Shamji, A.F.; et al. Selective killing of cancer cells by a small molecule targeting the stress response to ROS. Nature 2011, 475, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free Radic. Biol. Med. 2017, 104, 144–164. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Q.; Yu, M.X.; Hao, C.X.; Yang, W.H. Shikonin induces tumor apoptosis in glioma cells via endoplasmic reticulum stress, and Bax/Bak mediated mitochondrial outer membrane permeability. J. Ethnopharmacol. 2020, 263, 113059. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.G.; Lu, J.; Holmgren, A. Thioredoxin reductase is irreversibly modified by curcumin a novel molecular mechanism for its anticancer activity. J. Biol. Chem. 2005, 280, 25284–25290. [Google Scholar] [CrossRef] [Green Version]
- Dai, F.; Liu, G.Y.; Li, Y.; Yan, W.J.; Wang, Q.; Yang, J.; Lu, D.L.; Ding, D.J.; Lin, D.; Zhou, B. Insights into the importance for designing curcumin-inspired anticancer agents by a prooxidant strategy: The case of diarylpentanoids. Free Radic. Biol. Med. 2015, 85, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.J.; Wang, Q.; Yuan, C.H.; Wang, F.; Ji, Y.; Dai, F.; Jin, X.L.; Zhou, B. Designing piperlongumine-directed anticancer agents by an electrophilicity-based prooxidant strategy: A mechanistic investigation. Free Radic. Biol. Med. 2016, 97, 109–123. [Google Scholar] [CrossRef]
- Adamsa, D.J.; Daia, M.; Pellegrinoa, G.; Wagnera, B.K.; Stema, A.M.; Shamjia, A.F.; Schreibera, S.L. Synthesis, cellular evaluation and mechanism of action of piperlongumine analogs. Proc. Natl. Acad. Sci. USA 2012, 109, 15115–15120. [Google Scholar] [CrossRef] [Green Version]
- Hagmann, W.K. The many roles for fluorine in medicinal chemistry. J. Med. Chem. 2008, 51, 4359–4369. [Google Scholar] [CrossRef]
- Liang, X.J.; Guan, Y.P.; Zhang, B.C.; Liang, J.; Wang, B.C.; Li, Y.; Wang, J. Severe immune-related pneumonitis with PD-1 inhibitor after progression on previous PDL1 inhibitor in small cell lung cancer: A case report and review of literature. Front Oncol. 2019, 9, 1437. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.B.; Hu, L.; Yang, Z.J.; Sun, Y.; Hoffman, R.M.; Yi, Z. Aurora-A/NF-ĸB signaling is associated with radio-resistance in human lung adenocarcinoma. Anticancer. Res. 2019, 3911, 5991–5998. [Google Scholar] [CrossRef]
- Qin, L.M.; Zhong, M.L.; Adah, D.; Qin, L.; Chen, X.P.; Ma, C.Q.; Fu, Q.; Zhu, X.P.; Li, Z.L.; Wang, N.N.; et al. A novel tumour suppressor lncRNA F630028O10Rik inhibits lung cancer angiogenesis by regulating miR-223-3p. J. Cell Mol. Med. 2020, 24, 3549–3559. [Google Scholar] [CrossRef]
- Song, Y.X.; Zhou, B.; Du, X.Y.; Wang, Y.; Zhang, J.; Ai, Y.Q.; Xia, Z.J.; Zhao, G.F. Folic acid (FA)-conjugated mesoporous silica nanoparticles combined with MRP-1 siRNA improves the suppressive effects of myricetin on non-small cell lung cancer (NSCLC). Biomed. Pharmacother. 2020, 125, 109561. [Google Scholar] [CrossRef]
- Kong, Q.; Beel, J.A.; Lillehei, K.O. A threshold concept for cancer therapy. Med. Hypotheses 2000, 55, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Hatai, T.; Matsuzawa AInoshita, S.; Mochida, Y.; Kuroda, T.; Sakamaki, K.; Kuida, K.; Yonehara, S.; Ichijo, H.; Takeda, K. Execution of apoptosis signal-regulating kinase 1 (ASK1)-induced apoptosis by the mitochondria-dependent caspase activation. J. Biol. Chem. 2000, 275, 26576–26581. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Mu, W.W.; Liu, G.Y. Synthesis and evaluation of the anticancer activity of bischalcone analogs in human lung carcinoma (A549) cell line. Eur. J. Pharmacol. 2020, 888, 173396. [Google Scholar] [CrossRef]
- Liu, G.Y.; Zhai, Q.; Chen, J.Z.; Zhang, Z.Q.; Yang, J. 2,2′-Fluorine mono-carbonyl curcumin induce reactive oxygen species-Mediated apoptosis in Human lung cancer NCI-H460 cells. Eur. J. Pharmacol. 2016, 786, 161–168. [Google Scholar] [CrossRef]
- Liu, G.Y.; Sun, Y.Z.; Zhou, N.; Du, X.M.; Yang, J.; Guo, S.J. 3,3′-OH curcumin causes apoptosis in HepG2 cells through ROS-mediated pathway. Eur. J. Med. Chem. 2016, 112, 157–163. [Google Scholar] [CrossRef]
- Shi, H.L.; Li, Y.; Ren, X.R.; Zhang, Y.H.; Yang, Z.; Qi, C.Z. A novel quinazoline-based analog induces G2/M cell cycle arrest and apoptosis in human A549 lung cancer cells via a ROS-dependent mechanism. Biochem. Biophys. Res. Commun. 2017, 486, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Y.; Di, Z.M.; Cao, Q.; Xu, W.S.; Bi, S.X.; Yu, J.S.; Shen, Y.J.; Yu, Y.Q.; Shen, Y.X.; Feng, L.J. Xanthatin induces glioma cell apoptosis and inhibits tumor growth via activating endoplasmic reticulum stress-dependent CHOP pathway. Acta Pharmacol. Sin. 2020, 41, 404–414. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comps. | IC50/(µM) | Comps. | IC50/(µM) | ||
|---|---|---|---|---|---|
| A549 | SK-OV3 | A549 | SK-OV3 | ||
 | 4.3 ± 0.2 | 12.6 ± 1.4 | 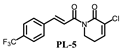 | 8.5 ± 0.3 | 15.4 ± 0.8 |
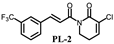 | 7.73 ± 0.1 | 23.2 ± 1.7 |  | 3.3 ± 0.2 | 18.5± 1.4 |
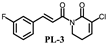 | >200 | >200 | 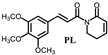 | 7.9 ± 0.8 | 19.4 ± 1.7 |
 | 19.6 ± 1.9 | 34.8 ± 2.8 | |||
| Comps. | k2 (M−1 s−1) | Comps. | k2 (M−1 s−1) |
|---|---|---|---|
| PL-1 | 5.70 ± 0.24 | PL-6 | 8.96 ± 0.04 |
| PL | 2.26 ± 0.04 |
| Signals Assigned | Chemical Shift (ppm) | Coupled Splitting | Coupling Constants (J/Hz) | Time (h) | Integral |
|---|---|---|---|---|---|
| H-9 H-9′ | 7.731 | dd | 8.5 | 0 | 2.00 |
| 7.658–7.759 | m | / | 1 | 2.00 | |
| H-3 | 7.332 | t | 4.5 | 0 | 1.00 |
| 7.332 | t | 4.5 | 1 | 0.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, A.-L.; Mu, W.-W.; Li, Y.-M.; Sun, Y.-L.; Li, P.-X.; Liu, R.-M.; Yang, J.; Liu, G.-Y. Piperlongumine Analogs Promote A549 Cell Apoptosis through Enhancing ROS Generation. Molecules 2021, 26, 3243. https://doi.org/10.3390/molecules26113243
Sun A-L, Mu W-W, Li Y-M, Sun Y-L, Li P-X, Liu R-M, Yang J, Liu G-Y. Piperlongumine Analogs Promote A549 Cell Apoptosis through Enhancing ROS Generation. Molecules. 2021; 26(11):3243. https://doi.org/10.3390/molecules26113243
Chicago/Turabian StyleSun, Ai-Ling, Wen-Wen Mu, Yan-Mo Li, Ya-Lei Sun, Peng-Xiao Li, Ren-Min Liu, Jie Yang, and Guo-Yun Liu. 2021. "Piperlongumine Analogs Promote A549 Cell Apoptosis through Enhancing ROS Generation" Molecules 26, no. 11: 3243. https://doi.org/10.3390/molecules26113243