
Antiplasmodial and Cytotoxic Flavonoids from Pappea capensis (Eckl. & Zeyh.) Leaves


Abstract
:1. Introduction
2. Results and Discussion
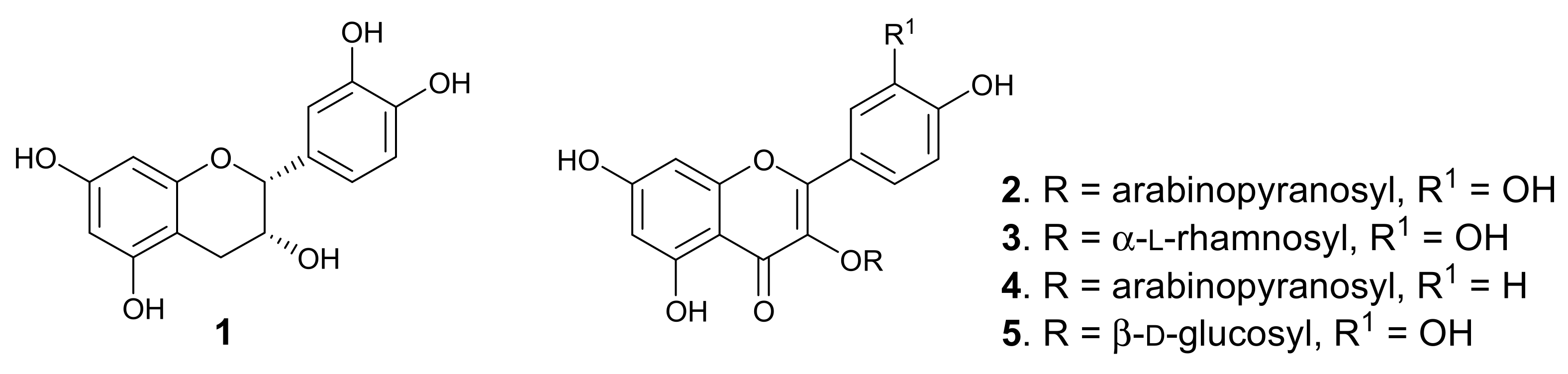
2.1. Chemistry
2.2. Biological Activity
3. Materials and Methods
3.1. General Procedures
3.2. HPLC and HPLC Conditions
3.3. Plant Material and Preparation of Extract for Bioassay
3.3.1. Isolation of Compounds from the Leaves of P. capensis
Spectroscopic Data of Compounds
3.4. Antimalarial Assay
3.4.1. The Parasites
3.4.2. Assessment of In Vitro Antiplasmodial Activity
3.5. In Vitro Cytotoxicity Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- WHO. World Malaria Report 2020; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Snow, R.W.; Sartorius, B.; Kyalo, D.; Maina, J.; Amratia, P.; Mundia, C.W.; Bejon, P.; Noor, A.M. The prevalence of Plasmodium falciparum in sub-Saharan Africa since 1900. Nature. 2017, 550, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwikkard, S.; van Heerden, F.R. Antimalarial activity of plant metabolites. Nat. Prod. Rep. 2002, 19, 675–692. [Google Scholar] [CrossRef] [PubMed]
- Wells, T.N.C. Natural products as starting points for future anti-malarial therapies: Going back to our roots? Malar. J. 2011, 10. [Google Scholar] [CrossRef] [Green Version]
- Coates Palgrave, K.; Drummond, R. Trees of Southern Africa; C. Struik Publishers: Cape Town, South Africa, 2002; p. 652. [Google Scholar]
- Cowling, R.; Proches, S.; Vlok, J.H.J. On the origin of southern African subtropical thicket vegetation. S. Afr. J. Bot. 2005, 71, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Hutchings, A. Zulu Medicinal Plants: An Inventory; University of Natal Press: Pietermaritzburg, South Africa, 1996; p. 189. [Google Scholar]
- Koch, A.; Tamez, P.; Pezzuto, J.; Soejarto, D. Evaluation of plants used for antimalarial treatment by the Maasai of Kenya. J. Ethnopharmacol. 2005, 101, 95–99. [Google Scholar] [CrossRef]
- Asnake, S.; Teklehaymanot, T.; Hymete, A.; Erko, B.; Giday, M. Antimalarial medicinal plants used by Gumuz people of Mandura Woreda, Benishangul-Gumuz regional state, Ethiopia. Indian J. Tradit. Knowl. 2016, 15, 546–552. [Google Scholar]
- Njoroge, G.N.; Bussmann, R.W. Diversity and utilization of antimalarial ethnophytotherapeutic remedies among the Kikuyus (Central Kenya). J. Ethnobiol. Ethnomed. 2006, 2, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bapela, M.J.; Meyer, J.J.M.; Kaiser, M. In vitro antiplasmodial screening of ethnopharmacologically selected South African plant species used for the treatment of malaria. J. Ethnopharmacol. 2014, 156, 370–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokoka, T.A.; Xolani, P.K.; Zimmermann, S.; Hata, Y.; Adams, M.; Kaiser, M.; Moodley, N.; Maharaj, V.; Koorbanally, N.A.; Hamburger, M.; et al. Antiprotozoal screening of 60 South African plants, and the identification of the antitrypanosomal germacranolides schkuhrin I and II. Planta Med. 2013, 79, 1380–1384. [Google Scholar] [CrossRef] [Green Version]
- Mulaudzi, R.B.; Ndhlala, A.R.; Kulkarni, M.G.; Finnie, J.F.; Van Staden, J. Anti-inflammatory and mutagenic evaluation of medicinal plants used by Venda people against venereal and related diseases. J. Ethnopharmacol. 2013, 146, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Pendota, S.C.; Aderogba, M.A.; Moyo, M.; McGaw, L.J.; Mulaudzi, R.B.; Van Staden, J. Antimicrobial, antioxidant and cytotoxicity of isolated compounds from leaves of Pappea capensis. S. Afr. J. Bot. 2017, 108, 272–277. [Google Scholar] [CrossRef]
- Tajuddeen, N.; Sallau, M.S.; Musa, A.M.; Yahaya, S.M.; Habila, J.D.; Ismail, A.M. A novel antimicrobial flavonoid from the stem bark of Commiphora pedunculata (Kotschy & Peyr.) Engl. Nat. Prod. Res. 2016, 30, 1109–1115. [Google Scholar] [PubMed]
- Da Silva Sa, F.A.; De Paula, J.A.M.; Dos Santos, P.A.; De Almeida Ribeiro Oliveira, L.; De Almeida Ribeiro Oliveira, G.; Liao, L.M.; De Paula, J.R.; Do Rosario Rodrigues Silva, M. Phytochemical analysis and antimicrobial activity of Myrcia tomentosa (Aubl.) DC. leaves. Molecules 2017, 22, 1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajuddeen, N.; Swart, T.; Hoppe, H.C.; van Heerden, F.R. Antiplasmodial and cytotoxic activities of extract and compounds from Ozoroa obovata (Oliv.) R. & A. Fern. var. obovata. Chem. Biodivers. 2021. [Google Scholar] [CrossRef]
- Vasange, M.; Liu, B.L.; Welch, C.J.; Rolfsen, W.C.; Bohlin, L. The flavonoid constituents of two Polypodium species (Calaguala) and their effect on the elastase release in human neutrophils. Planta Med. 1997, 63, 511–517. [Google Scholar] [CrossRef]
- Lee, S.; Park, H.-S.; Notsu, Y.; Ban, H.S.; Kim, Y.P.; Ishihara, K.; Hirasawa, N.; Jung, S.H.; Lee, Y.S.; Lim, S.S.; et al. Effects of hyperin, isoquercitrin and quercetin on lipopolysaccharide-induced nitrite production in rat peritoneal macrophages. Phytother. Res. 2008, 22, 1552–1556. [Google Scholar] [CrossRef] [PubMed]
- Umadevi, I.; Daniel, M. Chemosystematics of the Sapindaceae. Feddes Repert. 1991, 102, 607–612. [Google Scholar] [CrossRef]
- Silva, F.L.; Moreno, P.R.H.; Braz-Filho, R.; Tavares, J.F.; Barbosa-Filho, J.M. Chemical constituents of Cardiospermum corindum L. and their distribution in Sapindaceae. Biochem. Syst. Ecol. 2014, 57, 137–140. [Google Scholar] [CrossRef] [Green Version]
- Lima Cardoso, C.A.; Coelho, R.G.; Honda, N.K.; Pott, A.; Pavan, F.R.; Fujimura Leite, C.Q. Phenolic compounds and antioxidant, antimicrobial and antimycobacterial activities of Serjania erecta Radlk. (Sapindaceae). Braz. J. Pharm. Sci. 2013, 49, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Tajuddeen, N.; Van Heerden, F.R. Antiplasmodial natural products: An update. Malar. J. 2019, 18, 404. [Google Scholar] [CrossRef] [Green Version]
- Houel, E.; Nardella, F.; Jullian, V.; Valentin, A.; Vonthron-Senecheau, C.; Villa, P.; Obrecht, A.; Kaiser, M.; Bourreau, E.; Odonne, G.; et al. Wayanin and guaijaverin, two active metabolites found in a Psidium acutangulum Mart. ex DC (syn. P. persoonii McVaugh) (Myrtaceae) antimalarial decoction from the Wayana Amerindians. J. Ethnopharmacol. 2016, 187, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, D.; Fuehrer, H.-P.; Starzengrueber, P.; Swoboda, P.; Khan, W.A.; Reismann, J.A.B.; Mueller, M.S.K.; Chiba, P.; Noedl, H. Antiplasmodial activity of flavonol quercetin and its analogues in Plasmodium falciparum: Evidence from clinical isolates in Bangladesh and standardized parasite clones. Parasitol. Res. 2012, 110, 2289–2295. [Google Scholar] [CrossRef]
- Lehane, A.M.; Saliba, K.J. Common dietary flavonoids inhibit the growth of the intraerythrocytic malaria parasite. BMC Res Notes. 2008, 1, 26. [Google Scholar] [CrossRef] [Green Version]
- Atilaw, Y.; Muiva-Mutisya, L.; Ndakala, A.; Akala, H.M.; Yeda, R.; Wu, Y.J.; Coghi, P.; Wong, V.K.W.; Erdelyi, M.; Yenesew, A. Four Prenylflavone Derivatives with Antiplasmodial Activities from the Stem of Tephrosia purpurea subsp. leptostachya. Molecules 2017, 22, 1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezenyi, I.C.; Salawu, O.A.; Kulkarni, R.; Emeje, M. Antiplasmodial activity-aided isolation and identification of quercetin-4′-methyl ether in Chromolaena odorata leaf fraction with high activity against chloroquine-resistant Plasmodium falciparum. Parasitol. Res. 2014, 113, 4415–4422. [Google Scholar] [CrossRef]
- Kirmizibekmez, H.; Calis, I.; Perozzo, R.; Brun, R.; Donmez, A.A.; Linden, A.; Rudi, P.; Tasdemir, D. Inhibiting activities of the secondary metabolites of Phlomis brunneogaleata against parasitic protozoa and plasmodial enoyl-ACP reductase, a crucial enzyme in fatty acid biosynthesis. Planta Med. 2004, 70, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef]
- Makler, M.T.; Ries, J.M.; Williams, J.A.; Bancroft, J.E.; Piper, R.C.; Gibbins, B.L.; Hinrichs, D.J. Parasite lactate-dehydrogenase as an assay for plasmodium-falciparum drug sensitivity. Am. J. Trop. Med. Hyg. 1993, 48, 739–741. [Google Scholar] [CrossRef]


{kind=link}
| Compound | Viability% ± SD (50 µg/mL) | Viability% ± SD (10 µg/mL) | ||
|---|---|---|---|---|
| 3D7 | HeLa | 3D7 | HeLa | |
| P. capensis | 12.1 ± 0.2 | 1.9 ± 0.2 | 93.1 ± 3.8 | 71.4 ± 6.1 |
| 2 | 19.1 ± 0.5 | 2.8 ± 0.1 | 68.4 ± 3.4 | 72.4 ± 5.4 |
| 3 | 16.2 ± 2.2 | 1.4 ± 0.1 | 66.8 ± 2.6 | 71.3 ± 4.5 |
| 4 | 18.1 ± 1.0 | 3.1 ± 0.1 | 83.1 ± 3.5 | 70.9 ± 3.7 |
| 5 | 18.4 ± 2.9 | 1.9 ± 0.6 | 58.7 ± 1.9 | 64.8 ± 2.2 |
| Chloroquine a | - | - | <10 ± 2.1 | - |
| Emetine b | - | - | - | <5 ± 1.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tajuddeen, N.; Swart, T.; Hoppe, H.C.; van Heerden, F.R. Antiplasmodial and Cytotoxic Flavonoids from Pappea capensis (Eckl. & Zeyh.) Leaves. Molecules 2021, 26, 3875. https://doi.org/10.3390/molecules26133875
Tajuddeen N, Swart T, Hoppe HC, van Heerden FR. Antiplasmodial and Cytotoxic Flavonoids from Pappea capensis (Eckl. & Zeyh.) Leaves. Molecules. 2021; 26(13):3875. https://doi.org/10.3390/molecules26133875
Chicago/Turabian StyleTajuddeen, Nasir, Tarryn Swart, Heinrich C. Hoppe, and Fanie R. van Heerden. 2021. "Antiplasmodial and Cytotoxic Flavonoids from Pappea capensis (Eckl. & Zeyh.) Leaves" Molecules 26, no. 13: 3875. https://doi.org/10.3390/molecules26133875
APA StyleTajuddeen, N., Swart, T., Hoppe, H. C., & van Heerden, F. R. (2021). Antiplasmodial and Cytotoxic Flavonoids from Pappea capensis (Eckl. & Zeyh.) Leaves. Molecules, 26(13), 3875. https://doi.org/10.3390/molecules26133875