
Interstrain Variability of Human Vaginal Lactobacillus crispatus for Metabolism of Biogenic Amines and Antimicrobial Activity against Urogenital Pathogens

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
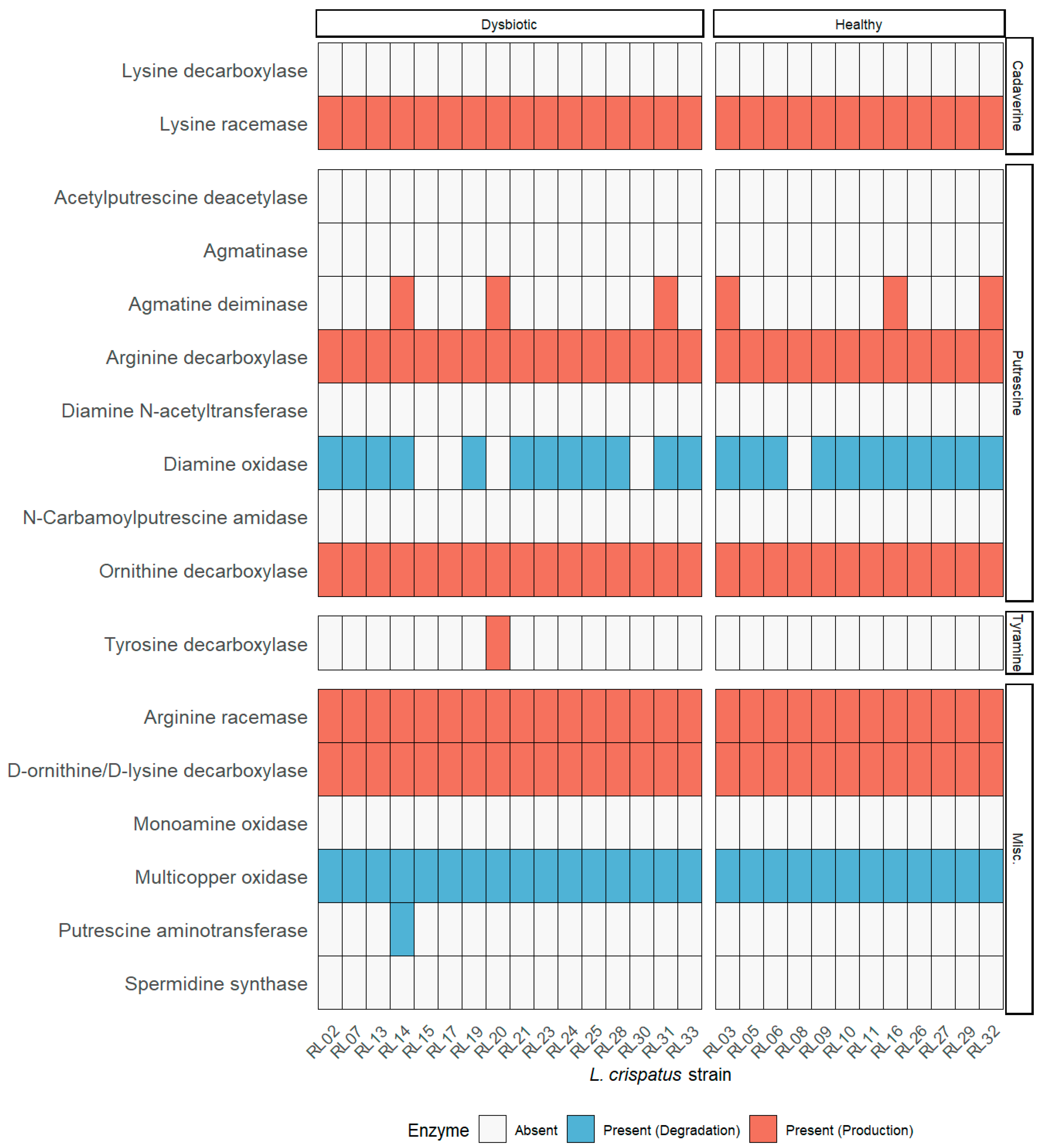
2.1. Untargeted Metabolomics and Functional Genomics
2.2. Targeted Metabolomics
2.3. Biogenic Amines Pathway Analysis
2.4. Antibacterial Activity
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains Used in This Study
4.2. Phylogenetic and Functional Genomics Analyses
4.3. LC-MS Protocol
4.4. Metabolite Identification
4.5. Biogenic Amines Pathway Analysis
4.6. Agar Well Diffusion Assays
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petrova, M.I.; Lievens, E.; Malik, S.; Imholz, N.; Lebeer, S. Lactobacillus species as biomarkers and agents that can promote various aspects of vaginal health. Front. Physiol. 2015, 6, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Barrons, R.; Tasone, D. Use of Lactobacillus probiotics for bacterial genitourinary infections in women: A review. Clin. Ther. 2008, 30, 453–468. [Google Scholar] [CrossRef]
- Schlabritz-Loutsevitch, N.; Gygax, S.E.; Dick, E.; Smith, W.L.; Snider, C.; Hubbard, G.; Ventolini, G. Vaginal dysbiosis from an evolutionary perspective. Sci. Rep. 2016, 6, 26817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillan, A.; Rulisa, S.; Sumarah, M.; Macklaim, J.M.; Renaud, J.; Bisanz, J.E.; Gloor, G.B.; Reid, G. A multi-platform metabolomics approach identifies highly specific biomarkers of bacterial diversity in the vagina of pregnant and non-pregnant women. Sci. Rep. 2015, 5, 14174. [Google Scholar] [CrossRef] [PubMed]
- Reid, G. Is bacterial vaginosis a disease? Appl. Microbiol. Biotechnol. 2018, 102, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.R.; Wierzbicki, M.R.; French, A.L.; Morris, S.; Newmann, S.; Reno, H.; Green, L.; Miller, S.; Powell, J.; Parks, T.; et al. Randomized trial of Lactin-V to prevent recurrence of bacterial vaginosis. N. Engl. J. Med. 2020, 382, 1906–1915. [Google Scholar] [CrossRef]
- Andreu, A.; Stapleton, A.E.; Fennell, C.L.; Hillier, S.L.; Stamm, W.E. Hemagglutination, adherence, and surface properties of vaginal Lactobacillus species. J. Infect. Dis. 1995, 171, 1237–1243. [Google Scholar] [CrossRef]
- Kim, J.-W.; Rajagopal, S.N. Antibacterial activities of Lactobacillus crispatus ATCC 33820 and Lactobacillus gasseri ATCC 33323. J. Microbiol. 2001, 39, 146–148. [Google Scholar]
- Nelson, T.M.; Borgogna, J.; Brotman, R.M.; Ravel, J.; Walk, S.T.; Yeoman, C.J. Vaginal biogenic amines: Biomarkers of bacterial vaginosis or precursors to vaginal dysbiosis? Front. Physiol. 2015, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puebla-Barragan, S.; Renaud, J.; Sumarah, M.; Reid, G. Malodorous biogenic amines in Escherichia coli-caused urinary tract infections in women—A metabolomics approach. Sci. Rep. 2020, 10, 9703. [Google Scholar] [CrossRef] [PubMed]
- Borgogna, J.; Shardell, M.D.; Grace, S.G.; Santori, E.K.; Americus, B.; Li, Z.; Ulanov, A.; Forney, L.; Nelson, T.M.; Brotman, R.M.; et al. Biogenic amines increase the odds of bacterial vaginosis and affect the growth of and lactic acid production by vaginal Lactobacillus spp. Appl. Environ. Microbiol. 2021, 87, e03068-20. [Google Scholar] [CrossRef]
- van der Veer, C.; Hertzberger, R.Y.; Bruisten, S.M.; Tytgat, H.L.P.; Swanenburg, J.; de Kat Angelino-Bart, A.; Schuren, F.; Molenaar, D.; Reid, G.; de Vries, H.; et al. Comparative genomics of human Lactobacillus crispatus isolates reveals genes for glycosylation and glycogen degradation: Implications for in vivo dominance of the vaginal microbiota. Microbiome 2019, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.I.; Matos, D.; San Romão, M.V.; Barreto Crespo, M.T. Dual role for the tyrosine decarboxylation pathway in Enterococcus faecium E17: Response to an acid challenge and generation of a proton motive force. Appl. Environ. Microbiol. 2009, 75, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, L.M.; Sammel, M.D.; Appleby, D.H.; Elovitz, M.A.; Baldwin, D.A.; Jeffcoat, M.K.; Macones, G.A.; Parry, S. Evidence of a gene-environment interaction that predisposes to spontaneous preterm birth: A role for asymptomatic bacterial vaginosis and DNA variants in genes that control the inflammatory response. Am. J. Obstet. Gynecol. 2010, 202, 386.e1–386.e6. [Google Scholar] [CrossRef] [PubMed]
- Elli, M.; Zink, R.; Rytz, A.; Reniero, R.; Morelli, L. Iron requirement of Lactobacillus spp. in completely chemically defined growth media. J. Appl. Microbiol. 2000, 88, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Pištěková, H.; Jančová, P.; Berčíková, L.; Buňka, F.; Sokolová, I.; Šopík, T.; Maršálková, K.; de Amaral, O.M.R.P.; Buňková, L. Application of qPCR for multicopper oxidase gene (MCO) in biogenic amines degradation by Lactobacillus casei. Food Microbiol. 2020, 91, 103550. [Google Scholar] [CrossRef]
- Li, B.; Wang, Y.; Xue, L.; Lu, S. Heterologous expression and application of multicopper oxidases from Enterococcus spp. for degradation of biogenic amines. Protein Pept. Lett. 2021, 28, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Borges, S.; Silva, J.; Teixeira, P. The role of lactobacilli and probiotics in maintaining vaginal health. Arch. Gynecol. Obstet. 2014, 289, 479–489. [Google Scholar] [CrossRef]
- Zamfir, M.; Callewaert, R.; Cornea, P.C.; Savu, L.; Vatafu, I.; De Vuyst, L. Purification and characterization of a bacteriocin produced by Lactobacillus acidophilus IBB 801. J. Appl. Microbiol. 1999, 87, 923–931. [Google Scholar] [CrossRef]
- McGroarty, J.A.; Reid, G. Detection of a Lactobacillus substance that inhibits Escherichia coli. Can. J. Microbiol. 1988, 34, 974–978. [Google Scholar] [CrossRef]
- Antonio, M.A.D.; Meyn, L.A.; Murray, P.J.; Busse, B.; Hillier, S.L. Vaginal colonization by probiotic Lactobacillus crispatus CTV-05 is decreased by sexual activity and endogenous lactobacilli. J. Infect. Dis. 2009, 199, 1506–1513. [Google Scholar] [CrossRef] [Green Version]
- Geshnizgani, A.M.; Onderdonk, A.B. Defined medium simulating genital tract secretions for growth of vaginal microflora. J. Clin. Microbiol. 1992, 30, 1323–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsen, T.; Nes, I.F.; Holo, H. Enterolysin A, a cell wall-degrading bacteriocin from Enterococcus faecalis LMG 2333. Appl. Environ. Microbiol. 2003, 69, 2975–2984. [Google Scholar] [CrossRef] [Green Version]
- Fremaux, C.; Klaenhammer, T.R. Helveticin J, a large heat-labile bacteriocin from Lactobacillus helveticus. In Bacteriocins of Lactic Acid Bacteria; De Vuyst, L., Vandamme, E., Eds.; Springer: Boston, MA, USA, 1994. [Google Scholar]
- Jiang, S.; Cai, L.; Lv, L.; Li, L. Pediococcus pentosaceus, a future additive or probiotic candidate. Microb. Cell Fact. 2021, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Agudelo Higuita, N.I.; Huycke, M.M. Enterococcal disease, epidemiology, and implications for treatment. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankarn, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Mysorekar, I.U.; Hultgren, S.J. Mechanisms of uropathogenic Escherichia coli persistence and eradication from the urinary tract. Proc. Natl. Acad. Sci. USA 2006, 103, 14170–14175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anukam, K.C.; Reid, G. Effects of metronidazole on growth of Gardnerella vaginalis ATCC 14018, probiotic Lactobacillus rhamnosus GR-1 and vaginal isolate Lactobacillus plantarum KCA. Microb. Ecol. Health Dis. 2008, 20, 48–52. [Google Scholar] [CrossRef]
- Atassi, F.; Brassart, D.; Grob, P.; Graf, F.; Servin, A.L. In vitro antibacterial activity of Lactobacillus helveticus strain KS300 against diarrhoeagenic, uropathogenic and vaginosis-associated bacteria. J. Appl. Microbiol. 2006, 101, 647–654. [Google Scholar] [CrossRef]
- Panthee, S.; Hamamoto, H.; Ishijima, S.A.; Paudel, A.; Sekimizu, K. Utilization of hybrid assembly approach to determine the genome of an opportunistic pathogenic fungus, Candida albicans TIMM 1768. Genome Biol. Evol. 2018, 10, 2017–2022. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyanhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T. Ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; von Mering, C.; Bork, P. Fast genome-wide functional annotation through orthology assignment by eggNOG-mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.5-7. 2020. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 15 May 2021).
- Petrova, M.I.; Macklaim, J.M.; Wuyts, S.; Verhoeven, T.; Vanderleyden, J.; Gloor, G.B.; Lebeer, S.; Reid, G. Comparative genomic and phenotypic analysis of the vaginal probiotic Lactobacillus rhamnosus GR-1. Front. Microbiol. 2018, 9, 1278. [Google Scholar] [CrossRef]
- Wuyts, S.; Wittouck, S.; De Boeck, I.; Allonsius, C.N.; Pasolli, E.; Segata, N.; Lebeer, S. Large-scale phylogenomics of the Lactobacillus casei group highlights taxonomic inconsistencies and reveals novel clade-associated features. mSystems 2017, 2, e00061-17. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Tautenhahn, R.; Böttcher, C.; Neumann, S. Highly sensitive feature detection for high resolution LC/MS. BMC Bioinform. 2008, 9, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, J.T.; Marcotte, E.M. Chromatographic alignment of ESI-LC-MS proteomics data sets by ordered bijective interpolated warping. Anal. Chem. 2006, 78, 6140–6152. [Google Scholar] [CrossRef]
- McMillan, A.; Renaud, J.B.; Gloor, G.B.; Reid, G.; Sumarah, M.W. Post-acquisition filtering of salt cluster artefacts for LC-MS based human metabolomic studies. J. Cheminform. 2016, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Holder, I.A.; Boyce, S.T. Agar well diffusion assay testing of bacterial susceptibility to various antimicrobials in concentrations non-toxic for human cells in culture. Burns 1994, 20, 426–429. [Google Scholar] [CrossRef]
- Tahara, T.; Oshimura, M.; Umezawa, C.; Kanatani, K. Isolation, partial characterization, and mode of action of Acidocin J1132, a two-component bacteriocin produced by Lactobacillus acidophilus JCM 1132. Appl. Environ. Microbiol. 1996, 62, 892–897. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests; R Package Version 0.7.0. 2021. Available online: https://cran.r-project.org/web/packages/rstatix/index.html (accessed on 15 May 2021).
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means; R Package Version 1.6.0. 2021. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 15 May 2021).
- Ogle, D.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis; R Package Version 0.8.32. 2021. Available online: https://cran.r-project.org/web/packages/FSA/index.html (accessed on 15 May 2021).
- Ahlmann-Eltze, C. Ggsignif: Significance Brackets for “Ggplot2”; R Package Version 0.6.1. 2021. Available online: https://cran.r-project.org/web/packages/ggsignif/ggsignif.pdf (accessed on 15 May 2021).
- Zeileis, A. Econometric Computing with HC and HAC Covariance Matrix Estimators. J. Stat. Softw. 2004, 11. Available online: https://cran.r-project.org/web/packages/sandwich/vignettes/sandwich.pdf (accessed on 15 May 2021). [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Group | Metabolomics Analysis | Genomics Analysis | Antimicrobial Test | Strain | Group | Metabolomics Analysis | Genomics Analysis | Antimicrobial Test |
|---|---|---|---|---|---|---|---|---|---|
| RL02 | DVM | yes | yes | yes | RL06 | LVM | no | yes | no |
| RL03 | LVM | yes | yes | yes | RL07 | DVM | no | yes | no |
| RL05 | LVM | yes | yes | yes | RL08 | LVM | no | yes | no |
| RL09 | LVM | yes | yes | yes | RL11 | LVM | no | yes | no |
| RL10 | LVM | yes | yes | yes | RL15 | DVM | no | yes | no |
| RL13 | DVM | yes | yes | yes | RL19 | DVM | no | yes | no |
| RL14 | DVM | yes | yes | yes | RL20 | DVM | no | yes | no |
| RL16 | LVM | yes | yes | yes | RL21 | DVM | no | yes | no |
| RL17 | DVM | yes | yes | yes | RL24 | DVM | no | yes | no |
| RL23 | DVM | yes | yes | yes | RL26 | LVM | no | yes | no |
| RL25 | DVM | yes | yes | yes | RL30 | DVM | no | yes | no |
| RL27 | LVM | yes | yes | yes | RL31 | DVM | no | yes | no |
| RL28 | DVM | yes | yes | yes | RL32 | LVM | no | yes | no |
| RL29 | LVM | yes | yes | yes | RL33 | DVM | no | yes | yes |
| Strains Added to Original Set | |||||||||
| RL01 | LVM | yes | no | yes | |||||
| RL04 | LVM | yes | no | yes | |||||
| RL12 | LVM | yes | no | yes | |||||
| RL22 | LVM | yes | no | yes | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puebla-Barragan, S.; Watson, E.; van der Veer, C.; Chmiel, J.A.; Carr, C.; Burton, J.P.; Sumarah, M.; Kort, R.; Reid, G. Interstrain Variability of Human Vaginal Lactobacillus crispatus for Metabolism of Biogenic Amines and Antimicrobial Activity against Urogenital Pathogens. Molecules 2021, 26, 4538. https://doi.org/10.3390/molecules26154538
Puebla-Barragan S, Watson E, van der Veer C, Chmiel JA, Carr C, Burton JP, Sumarah M, Kort R, Reid G. Interstrain Variability of Human Vaginal Lactobacillus crispatus for Metabolism of Biogenic Amines and Antimicrobial Activity against Urogenital Pathogens. Molecules. 2021; 26(15):4538. https://doi.org/10.3390/molecules26154538
Chicago/Turabian StylePuebla-Barragan, Scarlett, Emiley Watson, Charlotte van der Veer, John A. Chmiel, Charles Carr, Jeremy P. Burton, Mark Sumarah, Remco Kort, and Gregor Reid. 2021. "Interstrain Variability of Human Vaginal Lactobacillus crispatus for Metabolism of Biogenic Amines and Antimicrobial Activity against Urogenital Pathogens" Molecules 26, no. 15: 4538. https://doi.org/10.3390/molecules26154538
APA StylePuebla-Barragan, S., Watson, E., van der Veer, C., Chmiel, J. A., Carr, C., Burton, J. P., Sumarah, M., Kort, R., & Reid, G. (2021). Interstrain Variability of Human Vaginal Lactobacillus crispatus for Metabolism of Biogenic Amines and Antimicrobial Activity against Urogenital Pathogens. Molecules, 26(15), 4538. https://doi.org/10.3390/molecules26154538