
Anti-Cancer Effects of Zotarolimus Combined with 5-Fluorouracil Treatment in HCT-116 Colorectal Cancer-Bearing BALB/c Nude Mice

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
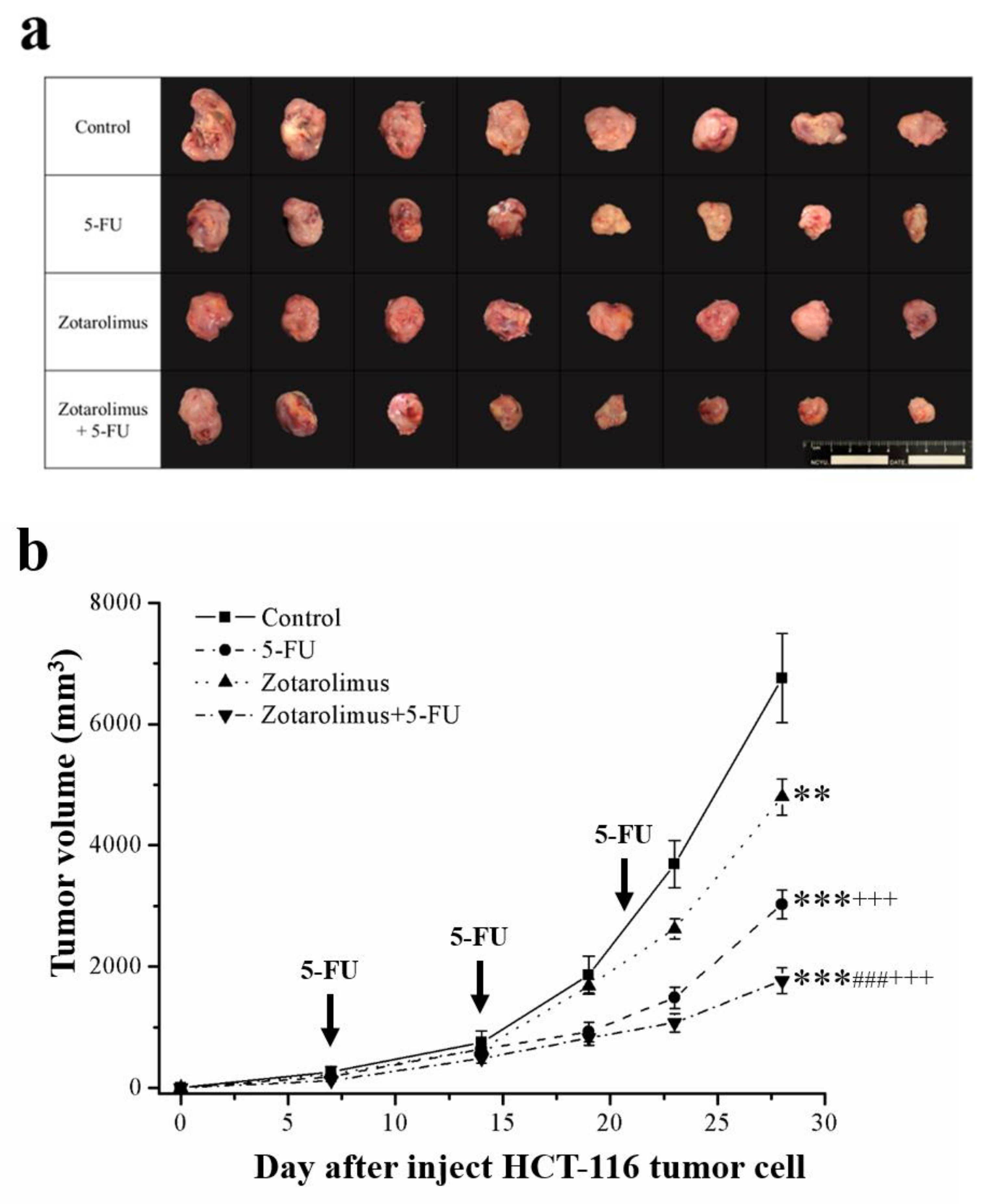
2.1. Zotarolimus Retards Tumor Growth
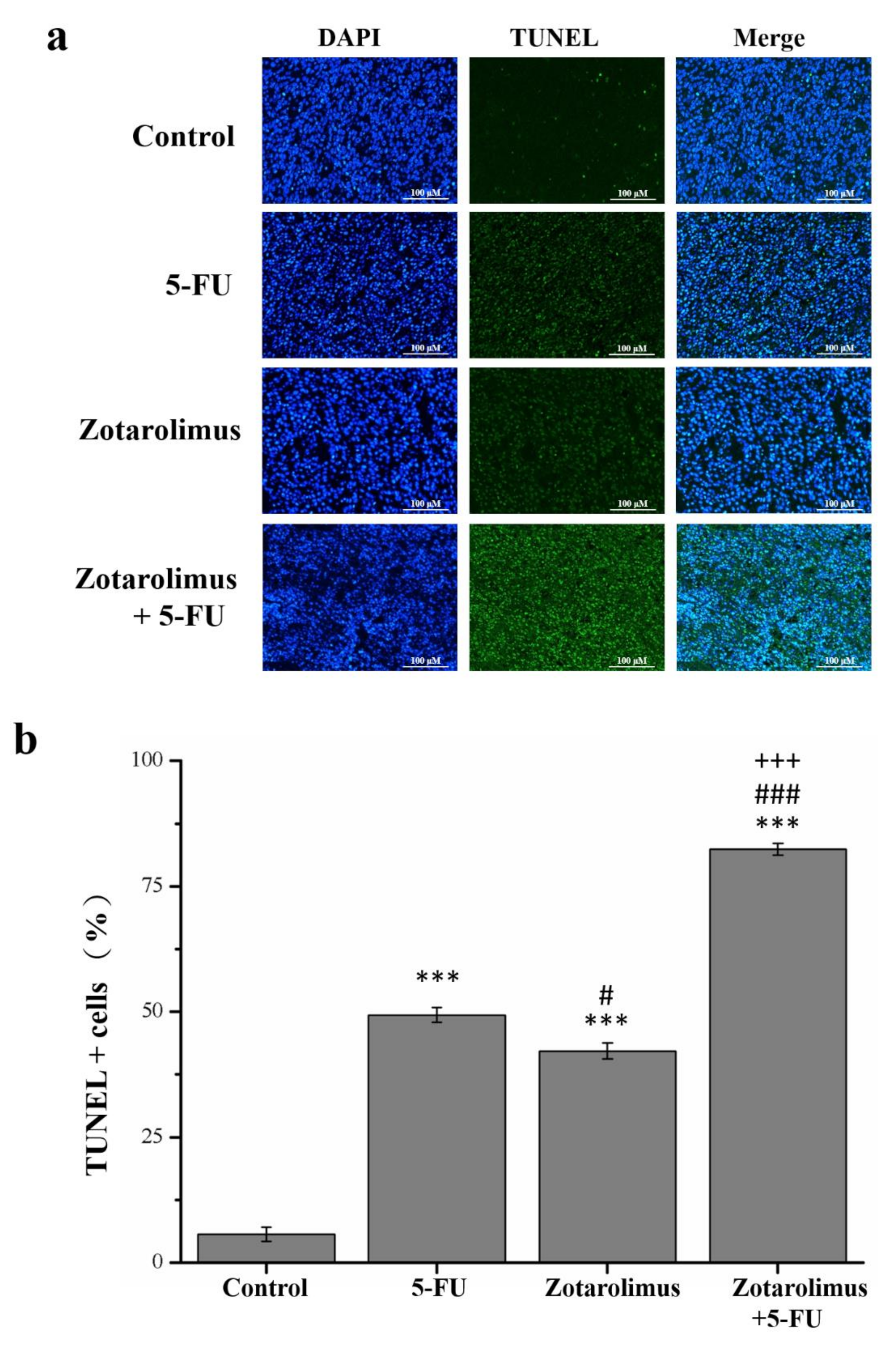
2.2. Zotarolimus Increases Number of TUNEL-Positive Cells
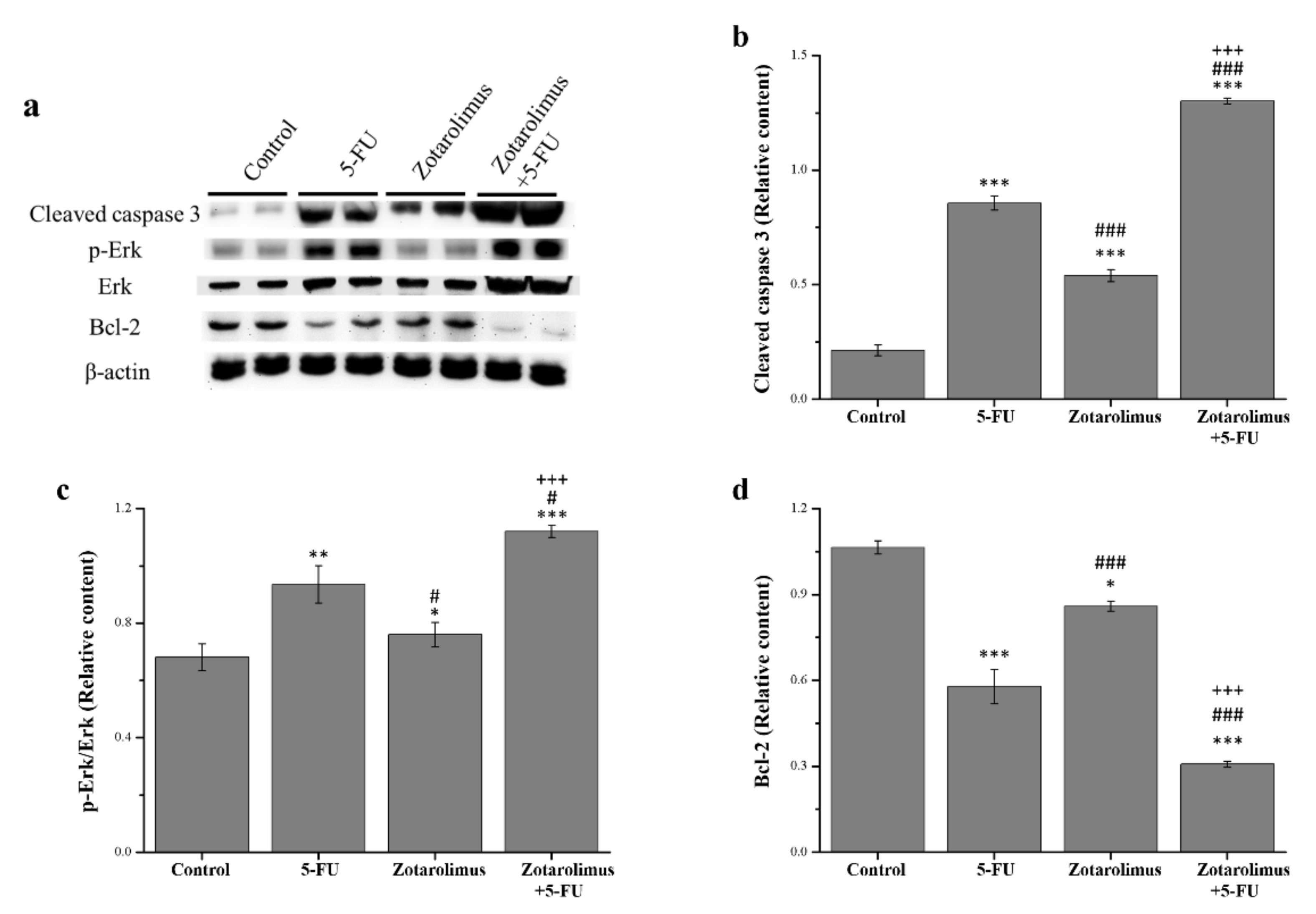
2.3. Zotarolimus Increases Apoptosis-Related Protein Expression
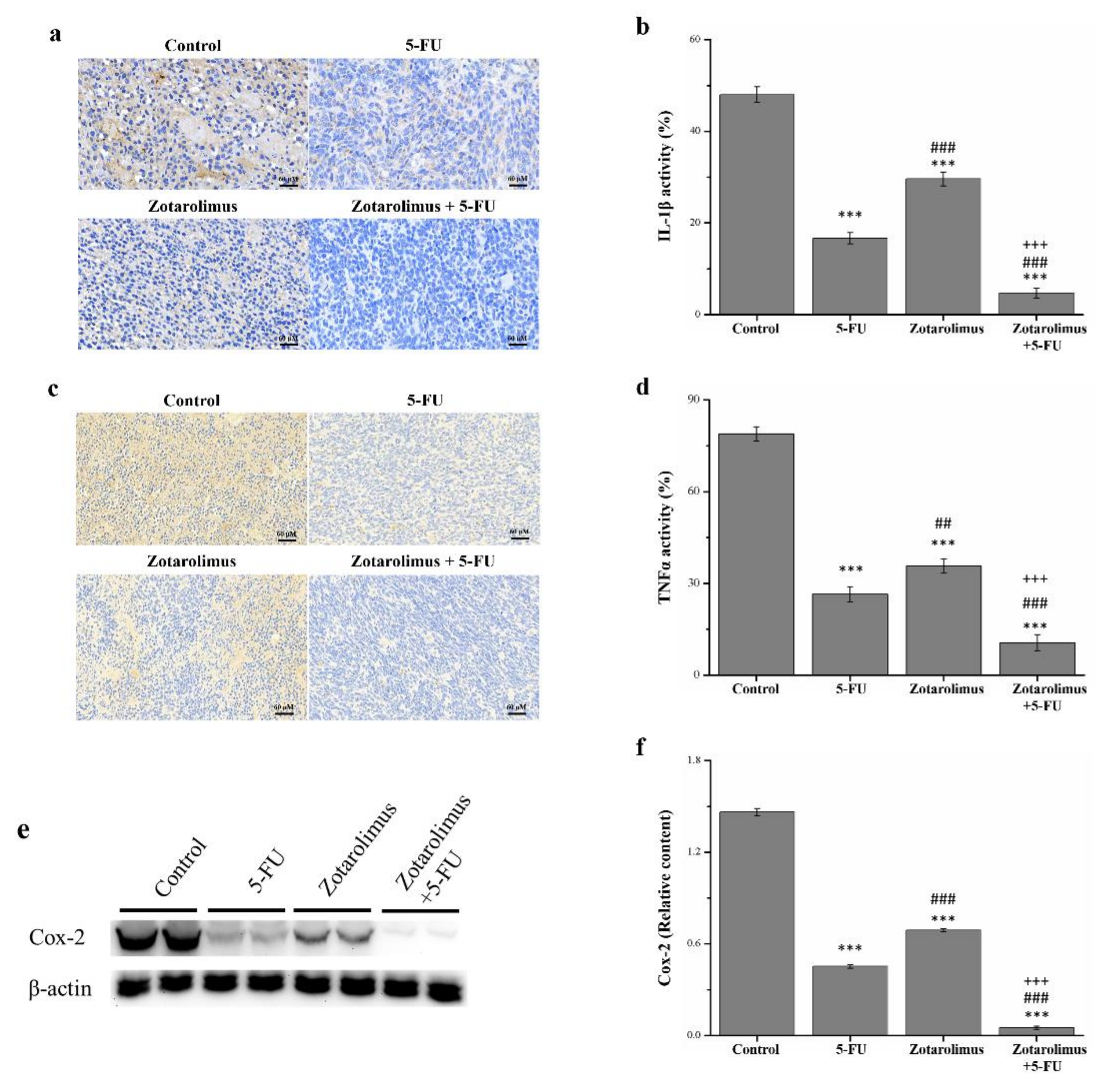
2.4. Zotarolimus Inhibits Production of Inflammation-Related Factors
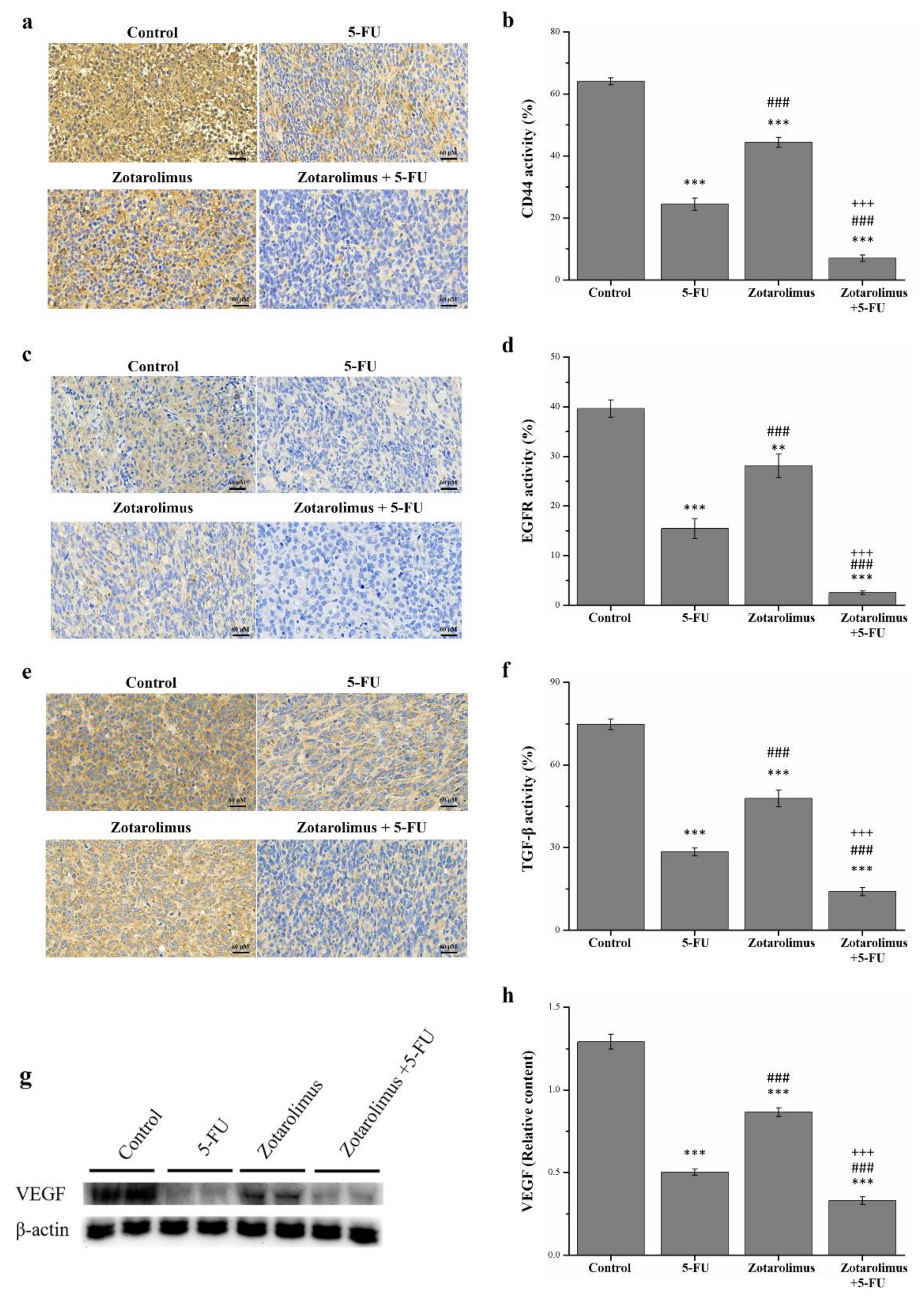
2.5. Zotarolimus Inhibits Metastasis-Related Factors
3. Discussion
4. Materials and Methods
4.1. Animal and Cell Lines
4.2. Tumor Inoculation and Treatment
4.3. Clinical Observations and Histopathological Analysis
4.4. Western Blotting Assay
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Mármol, I.; Sánchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal Carcinoma: A General Overview and Future Perspectives in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manchun, S.; Dass, C.R.; Cheewatanakornkool, K.; Sriamornsak, P. Enhanced anti-tumor effect of pH-responsive dextrin nanogels delivering doxorubicin on colorectal cancer. Carbohydr. Polym. 2015, 126, 222–230. [Google Scholar] [CrossRef]
- Cho, J.; Lee, J.; Bang, H.; Kim, S.T.; Park, S.H.; An, J.Y.; Choi, M.G.; Lee, J.H.; Sohn, T.S.; Bae, J.M.; et al. Programmed cell death-ligand 1 expression predicts survival in patients with gastric carcinoma with microsatellite instability. Oncotarget 2017, 8, 13320. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.P.; Sutton, P.A.; Winiarski, B.K.; Fenwick, S.W.; Malik, H.Z.; Vimalachandran, D.; Tweedle, E.M.; Costello, E.; Palmer, D.H.; Park, B.K.; et al. From mice to men: Murine models of colorectal cancer for use in translational research. Crit. Rev. Oncol. Hematol. 2016, 98, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Argov, M.; Kashi, R.; Peer, D.; Margalit, R. Treatment of resistant human colon cancer xenografts by a fluoxetine–doxorubicin combination enhances therapeutic responses comparable to an aggressive bevacizumab regimen. Cancer Lett. 2009, 274, 118–125. [Google Scholar] [CrossRef]
- Chiu, T.Y.; Chang, G.R.; Chen, W.Y.; Chao, T.H.; Mao, F.C.H. The anti-cancer effects of resveratrol combined with 5-fluorouracil treatment in balb/c mice bearing CT-26 Cells. J. Soc. Colon Rectal Surg. 2016, 27, 65–73. [Google Scholar]
- Chao, T.H.; Chang, G.R.; Chen, W.Y.; Chen, P.L.; Mao, F.C. The synergistic effect of rapamycin combined with 5-fluorouracil in BALB/cByJNarl mice bearing CT-26 tumor cells. Anticancer Res. 2014, 34, 3329–3335. [Google Scholar]
- Chiang, G.G.; Abraham, R.T. Targeting the mTOR signaling network in cancer. Trends. Mol. Med. 2007, 13, 433–442. [Google Scholar] [CrossRef]
- Dancey, J.E. Therapeutic targets: MTOR and related pathways. Cancer Biol. Ther. 2006, 5, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Sabatini, D.M. mTOR signaling in growth, metabolism, and disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, G.R.; Chiu, Y.S.; Wu, Y.Y.; Lin, Y.C.; Hou, P.H.; Mao, F.C. Rapamycin impairs HPD-induced beneficial effects on glucose homeostasis. Br. J. Pharmacol. 2015, 172, 3793–3804. [Google Scholar] [CrossRef] [Green Version]
- Unni, N.; Arteaga, C.L. Is dual mTORC1 and mTORC2 therapeutic blockade clinically feasible in cancer? JAMA Oncol. 2019, 5, 1564–1565. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Tao, T.; Li, H.; Zhu, X. mTOR signaling pathway and mTOR inhibitors in cancer: Progress and challenges. Cell Biosci. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Calne, R.Y.; Lim, S.; Samaan, A.; Collier, D.S.J.; Pollard, S.G.; White, D.J.G.; Thiru, S. Rapamycin for immunosuppression in organ allografting. Lancet 1989, 334, 227. [Google Scholar] [CrossRef]
- Morice, M.C.; Serruys, P.W.; Sousa, J.E.; Fajadet, J.; Ban Hayashi, E.; Perin, M.; Molnar, F. A randomized comparison of a sirolimus-eluting stent with a standard stent for coronary revascularization. N. Engl. J. Med. 2002, 346, 1773–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, G.R.; Chiu, Y.S.; Wu, Y.Y.; Chen, W.Y.; Liao, J.W.; Chao, T.H.; Mao, F.C. Rapamycin protects against high fat diet-induced obesity in C57BL/6J mice. J. Pharmacol. Sci. 2009, 109, 496–5032009. [Google Scholar] [CrossRef] [PubMed]
- Aliper, A.; Jellen, L.; Cortese, F.; Artemov, A.; Karpinsky-Semper, D.; Moskalev, A.; Swick, A.G.; Zhavoronkov, A. Towards natural mimetics of metformin and rapamycin. Aging 2017, 9, 2245–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, G.R.; Wu, Y.Y.; Chiu, Y.S.; Chen, W.Y.; Liao, J.W.; Hsu, H.M.; Chao, T.H.; Hung, S.W.; Mao, F.C. Long-term administration of rapamycin prevents against adiposity, but impairs glucose tolerance in high fat diet-fed KK/HlJ mice. Basic Clin Pharmacol. Toxicol. 2009, 105, 188–198. [Google Scholar] [CrossRef]
- Buellesfeld, L.; Grube, E. ABT-578-eluting stents. Herz 2004, 29, 167–170. [Google Scholar] [CrossRef]
- Hohenforst-Schmidt, W.; Zarogoulidis, P.; Pitsiou, G.; Linsmeier, B.; Tsavlis, D.; Kioumis, I.; Rittger, H. Drug eluting stents for malignant airway obstruction: A critical review of the literature. J. Cancer 2016, 7, 377. [Google Scholar] [CrossRef] [Green Version]
- Burke, S.E.; Kuntz, R.E.; Schwartz, L.B. Zotarolimus (ABT-578) eluting stents. Adv. Drug Deliv. Rev. 2006, 58, 437–446. [Google Scholar] [CrossRef]
- Mehilli, J.; Richardt, G.; Valgimigli, M.; Schulz, S.; Singh, A.; Abdel-Wahab, M.; Ott, I. Zotarolimus-versus everolimus-eluting stents for unprotected left main coronary artery disease. J. Am. Coll. Cardiol. 2013, 62, 2075–2082. [Google Scholar] [CrossRef] [Green Version]
- Lane, H.A.; Wood, J.M.; McSheehy, P.M.; Allegrini, P.R.; Boulay, A.; Brueggen, J.; O’Reilly, T. mTOR inhibitor RAD001 (everolimus) has antiangiogenic/vascular properties distinct from a VEGFR tyrosine kinase inhibitor. Clin. Cancer Res. 2009, 15, 1612–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cejka, D.; Preusser, M.; Woehrer, A.; Sieghart, W.; Strommer, S.; Werzowa, J.; Wacheck, V. Everolimus (RAD001) and anti-angiogenic cyclophosphamide show long-term control of gastric cancer growth in vivo. Cancer Biol. Ther. 2008, 7, 1377–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.D.; Eng, C. The promise of mTOR inhibitors in the treatment of colorectal cancer. Expert Opin. Investig. Drugs 2012, 21, 1775–1788. [Google Scholar] [CrossRef]
- Vodenkova, S.; Buchler, T.; Cervena, K.; Veskrnova, V.; Vodicka, P.; Vymetalkova, V. 5-fluorouracil and other fluoropyrimidines in colorectal cancer: Past, present and future. Pharmacol. Ther. 2020, 206, 107447. [Google Scholar] [CrossRef]
- Wei, Y.; Yang, P.; Cao, S.; Zhao, L. The combination of curcumin and 5-fluorouracil in cancer therapy. Arch. Pharm. Res. 2018, 41, 1–13. [Google Scholar] [CrossRef]
- Li, T.; Yan, G.; Bai, Y.; Wu, M.; Fang, G.; Zhang, M.; Xie, Y.; Borjigidai, A.; Fu, B. Papain bioinspired gold nanoparticles augmented the anticancer potency of 5-FU against lung cancer. J. Exp. Nanosci. 2020, 15, 109–128. [Google Scholar] [CrossRef]
- Polk, A.; Vaage-Nilsen, M.; Vistisen, K.; Nielsen, D.L. Cardiotoxicity in cancer patients treated with 5-fluorouracil or capecitabine: A systematic review of incidence, manifestations and predisposing factors. Cancer Treat. Rev. 2013, 39, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.F.; Wu, C.Y.; Chiou, R.Y.; Yang, W.C.; Lin, C.F.; Wang, C.M.; Hou, P.H.; Lin, T.C.; Kuo, C.Y.; Chang, G.R. The Anti-Cancer Effects of a Zotarolimus and 5-Fluorouracil Combination Treatment on A549 Cell-Derived Tumors in BALB/c Nude Mice. Int. J. Mol. Sci. 2021, 22, 4562. [Google Scholar] [CrossRef] [PubMed]
- Kyrylkova, K.; Kyryachenko, S.; Leid, M.; Kioussi, C. Detection of apoptosis by TUNEL assay. Methods Mol. Biol. 2012, 887, 41–47. [Google Scholar]
- Sankari, S.L.; Masthan, K.M.; Babu, N.A.; Bhattacharjee, T.; Elumalai, M. Apoptosis in cancer—An update. Asian Pac. J. Cancer Prev. 2012, 13, 4873–4878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.J.; Prickril, B.; Rasooly, A. Mechanisms of phytonutrient modulation of cyclooxygenase-2 (COX-2) and inflammation related to cancer. Nutr. Cancer 2018, 70, 350–375. [Google Scholar] [CrossRef] [PubMed]
- Guan, X. Cancer metastases: Challenges and opportunities. Acta Pharm. Sin. B 2015, 5, 402–418. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Li, X.; Zhang, J. mTOR signaling in cancer and mTOR inhibitors in solid tumor targeting therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, I.A.; Arteaga, C.L. The PI3K/AKT pathway as a target for cancer treatment. Annu. Rev. Med. 2016, 67, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Leite, R.; Arantes-Rodrigues, R.; Sousa, N.; Oliveira, P.A.; Santos, L. mTOR inhibitors in urinary bladder cancer. Tumor Biol. 2016, 37, 11541–11551. [Google Scholar] [CrossRef] [PubMed]
- Wislez, M.; Spencer, M.L.; Izzo, J.G.; Juroske, D.M.; Balhara, K.; Cody, D.D.; Price, R.E.; Hittelman, W.N.; Wistuba, I.I.; Kurie, J.M. Inhibition of mammalian target of rapamycin reverses alveolar epithelial neoplasia induced by oncogenic K-ras. Cancer Res. 2005, 65, 3226–3235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Li, X.; Zhen, L.; Chen, W.; Mu, L.; Zhang, Y.; Song, A. Everolimus inhibits breast cancer cell growth through PI3K/AKT/mTOR signaling pathway. Mol. Med. Rep. 2018, 17, 7163–7169. [Google Scholar] [CrossRef] [Green Version]
- Ohara, T.; Takaoka, M.; Toyooka, S.; Tomono, Y.; Nishikawa, T.; Shirakawa, Y.; Naomoto, Y. Inhibition of mTOR by temsirolimus contributes to prolonged survival of mice with pleural dissemination of non-small-cell lung cancer cells. Cancer Sci. 2011, 102, 1344–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera, V.M.; Squillace, R.M.; Miller, D.; Berk, L.; Wardwell, S.D.; Ning, Y.; Clackson, T. Ridaforolimus (AP23573; MK-8669), a potent mTOR inhibitor, has broad antitumor activity and can be optimally administered using intermittent dosing regimens. Mol. Cancer Ther. 2011, 10, 1059–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cejka, D.; Preusser, M.; Fuereder, T.; Sieghart, W.; Werzowa, J.; Strommer, S.; Wacheck, V. mTOR inhibition sensitizes gastric cancer to alkylating chemotherapy in vivo. Anticancer Res. 2008, 28, 3801–3808. [Google Scholar] [PubMed]
- Cirstea, D.; Hideshima, T.; Rodig, S.; Santo, L.; Pozzi, S.; Vallet, S.; Raje, N. Dual inhibition of Akt/mammalian target of rapamycin pathway by nanoparticle albumin-bound–rapamycin and perifosine induces antitumor activity in multiple myeloma. Mol. Cancer Ther. 2010, 9, 963–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeffer, C.M.; Singh, A.T. Apoptosis: A target for anticancer therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Zaman, S.; Wang, R.; Gandhi, V. Targeting the apoptosis pathway in hematologic malignancies. Leuk. Lymphoma 2014, 55, 1980–1992. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Andersen, J.S.; Mann, M.; Terada, N.; Korsmeyer, S.J. p70S6 kinase signals cell survival as well as growth, inactivating the pro-apoptotic molecule BAD. Proc. Natl. Acad. Sci. USA 2001, 98, 9666–9670. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A. Inflammation by remote control. Nature 2005, 435, 752–753. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.L.; Berggren, K.L.; Restrepo Cruz, S.; Gan, G.N.; Beswick, E.J. Inhibition of MK2 suppresses IL-1β, IL-6, and TNF-α-dependent colorectal cancer growth. Int. J. Cancer 2018, 142, 1702–1711. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.C.; Chaen, Y.L.; Hsu, C.P. Clinical significance of tissue expression of interleukin-6 in colorectal carcinoma. Anticancer Res. 2006, 26, 3905–3911. [Google Scholar]
- El-Omar, E.M.; Rabkin, C.S.; Gammon, M.D.; Vaughan, T.L.; Risch, H.A.; Schoenberg, J.B.; Chow, W.H. Increased risk of noncardia gastric cancer associated with proinflammatory cytokine gene polymorphisms. Gastroenterology 2003, 124, 1193–1201. [Google Scholar] [CrossRef]
- Wu, J.; Cai, Q.; Li, H.; Cai, H.; Gao, J.; Yang, G.; Shu, X.O. Circulating C-reactive protein and colorectal cancer risk: A report from the Shanghai Men’s Health Study. Carcinogenesis 2013, 34, 2799–2803. [Google Scholar] [CrossRef] [Green Version]
- Erlinger, T.P.; Platz, E.A.; Rifai, N.; Helzlsouer, K.J. C-reactive protein and the risk of incident colorectal cancer. JAMA 2004, 291, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Otani, T.; Iwasaki, M.; Sasazuki, S.; Inoue, M.; Tsugane, S.; Japan Public Health Center–Based Prospective Study Group. Plasma C-reactive protein and risk of colorectal cancer in a nested case-control study: Japan Public Health Center–based prospective study. Cancer Epidemiol. Biomarkers Prev. 2006, 15, 690–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shay, T.; Lederer, J.A.; Benoist, C. Genomic responses to inflammation in mouse models mimic humans: We concur, apples to oranges comparisons won’t do. Proc. Natl. Acad. Sci. 2015, 112, E346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, J.W.; Baldwin, A.S. IKK/nuclear factor-kappaB and oncogenesis: Roles in tumor-initiating cells and in the tumor microenvironment. Adv. Cancer Res. 2014, 121, 125–145. [Google Scholar] [PubMed]
- Morotti, A.; Crivellaro, S.; Panuzzo, C.; Carrà, G.; Guerrasio, A.; Saglio, G. IκB-α: At the crossroad between oncogenic and tumor-suppressive signals. Oncol. Lett. 2017, 13, 531–534. [Google Scholar] [CrossRef] [Green Version]
- Voboril, R.; Weberova-Voborilova, J. Constitutive NF-kappaB activity in colorectal cancer cells: Impact on radiation-induced NF-kappaB activity, radiosensitivity, and apoptosis. Neoplasma 2006, 53, 518–523. [Google Scholar]
- Lin, R.; Piao, M.; Song, Y.; Liu, C. Quercetin suppresses AOM/DSS-induced colon carcinogenesis through its anti-inflammation effects in mice. J. Immunol. Res. 2020, 2020, 9242601. [Google Scholar] [CrossRef]
- Kleniewska, P.; Hoffmann, A.; Pniewska, E.; Pawliczak, R. The influence of probiotic Lactobacillus casei in combination with prebiotic inulin on the antioxidant capacity of human plasma. Oxid. Med. Cell Longev. 2016, 2016, 1340903. [Google Scholar] [CrossRef] [Green Version]
- Johansson, E.; Grassi, E.S.; Pantazopoulou, V.; Tong, B.; Lindgren, D.; Berg, T.J.; Pietras, A. CD44 interacts with HIF-2α to modulate the hypoxic phenotype of perinecrotic and perivascular glioma cells. Cell Rep. 2017, 20, 1641–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, N.R.; Cha, J.H.; Jang, J.W.; Bae, S.H.; Jang, B.; Kim, J.H.; Yoon, S.K. Synergistic effects of CD44 and TGF-β1 through AKT/GSK-3β/β-catenin signaling during epithelial-mesenchymal transition in liver cancer cells. Biochem. Biophys. Res. Commun. 2016, 477, 568–574. [Google Scholar] [CrossRef]
- Massagué, J.; Blain, S.W.; Lo, R.S. TGFβ signaling in growth control, cancer, and heritable disorders. Cell 2000, 103, 295–309. [Google Scholar] [CrossRef] [Green Version]
- Ciftci, K.; Su, J.; Trovitch, P.B. Growth factors and chemotherapeutic modulation of breast cancer cells. J. Pharm. Pharmacol. 2003, 55, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Baselga, J.; Arteaga, C.L. Critical update and emerging trends in epidermal growth factor receptor targeting in cancer. J. Clin. Oncol. 2005, 23, 2445–2459. [Google Scholar] [CrossRef]
- Kerbel, R.S. Tumor angiogenesis. N. Engl. J. Med. 2008, 358, 2039–2049. [Google Scholar] [CrossRef] [Green Version]
- Lichtenberger, B.M.; Tan, P.K.; Niederleithner, H.; Ferrara, N.; Petzelbauer, P.; Sibilia, M. Autocrine VEGF signaling synergizes with EGFR in tumor cells to promote epithelial cancer development. Cell 2010, 140, 268–279. [Google Scholar] [CrossRef] [Green Version]
- Keegan, N.M.; Gleeson, J.P.; Hennessy, B.T.; Morris, P.G. PI3K inhibition to overcome endocrine resistance in breast cancer. Expert Opin. Investig. Drugs 2018, 27, 1–15. [Google Scholar] [CrossRef]
- Wang, H.; Jia, X.H.; Chen, J.R.; Wang, J.Y.; Li, Y.J. Osthole shows the potential to overcome P-glycoprotein-mediated multidrug resistance in human myelogenous leukemia K562/ADM cells by inhibiting the PI3K/Akt signaling pathway. Oncol. Rep. 2016, 35, 3659–3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Shao, R.; Li, L.; Xu, Z.P.; Gu, W. Effective inhibition of colon cancer cell growth with MgAl-layered double hydroxide (LDH) loaded 5-FU and PI3K/mTOR dual inhibitor BEZ-235 through apoptotic pathways. Int. J. Nanomed. 2014, 9, 3403. [Google Scholar]
- Zhang, B.; Jin, K.; Jiang, T.; Wang, L.; Shen, S.; Luo, Z.; Pang, Z. Celecoxib normalizes the tumor microenvironment and enhances small nanotherapeutics delivery to A549 tumors in nude mice. Sci. Rep. 2017, 7, 1–12. [Google Scholar]
- Park, J.H.; Zhao, M.; Han, Q.; Sun, Y.; Higuchi, T.; Sugisawa, N.; Yamamoto, J.; Singh, S.R.; Clary, B.; Bouvet, M.; et al. Efficacy of oral recombinant methioninase combined with oxaliplatinum and 5-fluorouracil on primary colon cancer in a patient-derived orthotopic xenograft mouse model. Biochem. Biophys. Res. Commun. 2019, 518, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Attafi, I.M.; Bakheet, S.A.; Korashy, H.M. The role of NF-κB and AhR transcription factors in lead-induced lung toxicity in human lung cancer A549 cells. Toxicol. Mech. Methods 2020, 30, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Li, H.; Chen, Y.; Lv, H.; Liu, L.; Ran, J.; Li, H. Survivin activates NF-κB p65 via the IKKβ promoter in esophageal squamous cell carcinoma. Mol. Med. Rep. 2016, 13, 1869–1880. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Weight (g) | Tumor Inhibition Rate (%) | |
|---|---|---|
| Control | 7.10 ± 1.19 | - |
| 5-FU | 2.69 ± 0.44 *** | 62.1 |
| Zotarolimus | 4.51 ± 0.21 ***### | 36.5 |
| Zotarolimus + 5-FU | 1.31 ± 0.15 ***###+++ | 81.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, G.-R.; Kuo, C.-Y.; Tsai, M.-Y.; Lin, W.-L.; Lin, T.-C.; Liao, H.-J.; Chen, C.-H.; Wang, Y.-C. Anti-Cancer Effects of Zotarolimus Combined with 5-Fluorouracil Treatment in HCT-116 Colorectal Cancer-Bearing BALB/c Nude Mice. Molecules 2021, 26, 4683. https://doi.org/10.3390/molecules26154683
Chang G-R, Kuo C-Y, Tsai M-Y, Lin W-L, Lin T-C, Liao H-J, Chen C-H, Wang Y-C. Anti-Cancer Effects of Zotarolimus Combined with 5-Fluorouracil Treatment in HCT-116 Colorectal Cancer-Bearing BALB/c Nude Mice. Molecules. 2021; 26(15):4683. https://doi.org/10.3390/molecules26154683
Chicago/Turabian StyleChang, Geng-Ruei, Chan-Yen Kuo, Ming-Yang Tsai, Wei-Li Lin, Tzu-Chun Lin, Huei-Jyuan Liao, Chung-Hung Chen, and Yu-Chen Wang. 2021. "Anti-Cancer Effects of Zotarolimus Combined with 5-Fluorouracil Treatment in HCT-116 Colorectal Cancer-Bearing BALB/c Nude Mice" Molecules 26, no. 15: 4683. https://doi.org/10.3390/molecules26154683
APA StyleChang, G.-R., Kuo, C.-Y., Tsai, M.-Y., Lin, W.-L., Lin, T.-C., Liao, H.-J., Chen, C.-H., & Wang, Y.-C. (2021). Anti-Cancer Effects of Zotarolimus Combined with 5-Fluorouracil Treatment in HCT-116 Colorectal Cancer-Bearing BALB/c Nude Mice. Molecules, 26(15), 4683. https://doi.org/10.3390/molecules26154683