1. Introduction
Ceratonia siliqua L., also known as the Mediterranean carob tree, has been widely studied for the functional chemicals contained in its fruit. The carob fruit contains two major parts (by weight): the pulp (90%) and the seeds (10%). The chemical composition of the carob pulp differs widely depending on cultivar or species, origin, climate, as well as stage of maturity during harvesting [
1,
2,
3]. Recently, research has focused on the many beneficial effects of carobs for human health [
4,
5]. In addition to carobs, recent evidence suggests that extracts derived from peels [
6], leaves [
7] and seeds [
8] of different plant species, edible or not, may induce diverse health-promoting effects, often different from that expected, due to their rich content of bioactive compounds. The carob fruit is a complex mixture of mainly sugars and fibers, followed by a great diversity of polyphenols. Carob pulp constituents, including fibers, cyclitols, polyphenols and tannins have been associated with the health-promoting effects of carobs against diabetes, diarrhea and cancer [
9,
10,
11,
12,
13]. Additionally, the anti-cancer properties of carob pulp have been attributed mainly to the presence of polyphenolic compounds, which constitute up to 20% of the carob pulp [
14,
15].
The ripening of the carob fruit significantly affects its composition in phenolic content and, therefore, its biological properties. Unripe carobs have higher amounts of phenolic acids, polyphenols, flavonoids and tannins and display higher in vitro anti-oxidant capacity whereas the ripe fruit contains high levels of Gallic acid (GA), due to the enzymatic hydrolysis of gallotannins in immature fruit [
3]. GA, a phenolic acid, as well as Myricetin, Naringenin and Kaempferol, flavonoids commonly found in carob fruit, have been shown to exert anti-cancer properties in vitro by activating caspase-dependent programmed cell death (CD-PCD) and cell cycle arrest [
16,
17,
18,
19,
20,
21]. Mitochondrial or intrinsic apoptosis is controlled by the members of the Bcl-2 superfamily. Anti-apoptotic Bcl-2 is often overexpressed in cancer cells and blocks mitochondrial membrane pore permeabilization (MOMP). Following apoptosis induction, Bcl-2 is reduced, pro-apoptotic factors exit the mitochondria and activate caspase-9 which cleaves downstream caspases-3, -6 and -7 [
22]. In addition to CD-PCD, many natural agents can also cause cell death by caspase-independent apoptosis (CI-PCD), via cleavage of the apoptosis inducing factor (AIF). Even though different proteins are implicated in these two pathways, the crosstalk between them has been reported in the literature [
23].
Cyprus is among the top producers of carobs worldwide. The chemical and biological properties of Cypriot carobs and derived products have recently come to focus. Kyriacou et al. revealed that major metabolic events occur during carob ripening that significantly affect the phenolic content and anti-oxidant capacity of carob-pod derived extracts [
3]. Kibble size was found to be an important parameter in determining the chemical characteristics of the resulting carob juice produced by different extraction methods [
24]. The geographical origin, type and nutritional composition of carobs from Cyprus and other countries, were differentiated based on Fourier transform infrared (FTIR) spectroscopy and chemometrics [
25,
26]. In addition, the profile of biogenic volatile organic compounds (VOCs) emitted from carob fruit that contribute to its unique aroma, has been recently described [
27]. However, the potential anticancer effects of carob fruit extracts have not yet been explored. Our main objectives were to evaluate and compare for the first time, the anti-cancer potential of carob extracts derived from Cypriot carobs at different maturity stages and from different parts of the carob pods. In addition, we assessed the properties of extracts derived in solvents of different polarity, that is known to affect polyphenol content [
28]. We investigated the anti-proliferative and pro-apoptotic effect of the carob extracts using established cancer and normal breast cell lines. Furthermore, we determined the total polyphenolic content (TPC) of these extracts and their anti-oxidant ability. Finally, we performed LC-MS and identified the dominant polyphenols in the most potent anti-cancer extracts. Our results indicate that depending on the extraction solvent used and stage of maturity, carob fruit extracts from trees grown in Cyprus have anti-cancer properties that may be attributed to the presence of polyphenols that can induce caspase-dependent and independent apoptosis in breast cancer cells.
3. Discussion
As part of this study, we obtained extracts from the whole carob fruit, as well as the seeds and pulp separately. We investigated the anti-proliferative effects of Cypriot carob extracts in cancer cells and normal breast cell lines, their ability to induce apoptosis as well as the mechanism by which they exert their action. In addition, we determined the TPC of each extract as well as their anti-oxidant activity. Finally, we performed LC-MS analysis in the most promising anti-cancer extracts. Although several studies have been performed for the identification and quantification of polyphenols in carob fruit, research has highlighted that different extraction methods produce diverse patterns and compositions, changing the phenolic profile, and, therefore, the function of the derived extract [
24,
34].
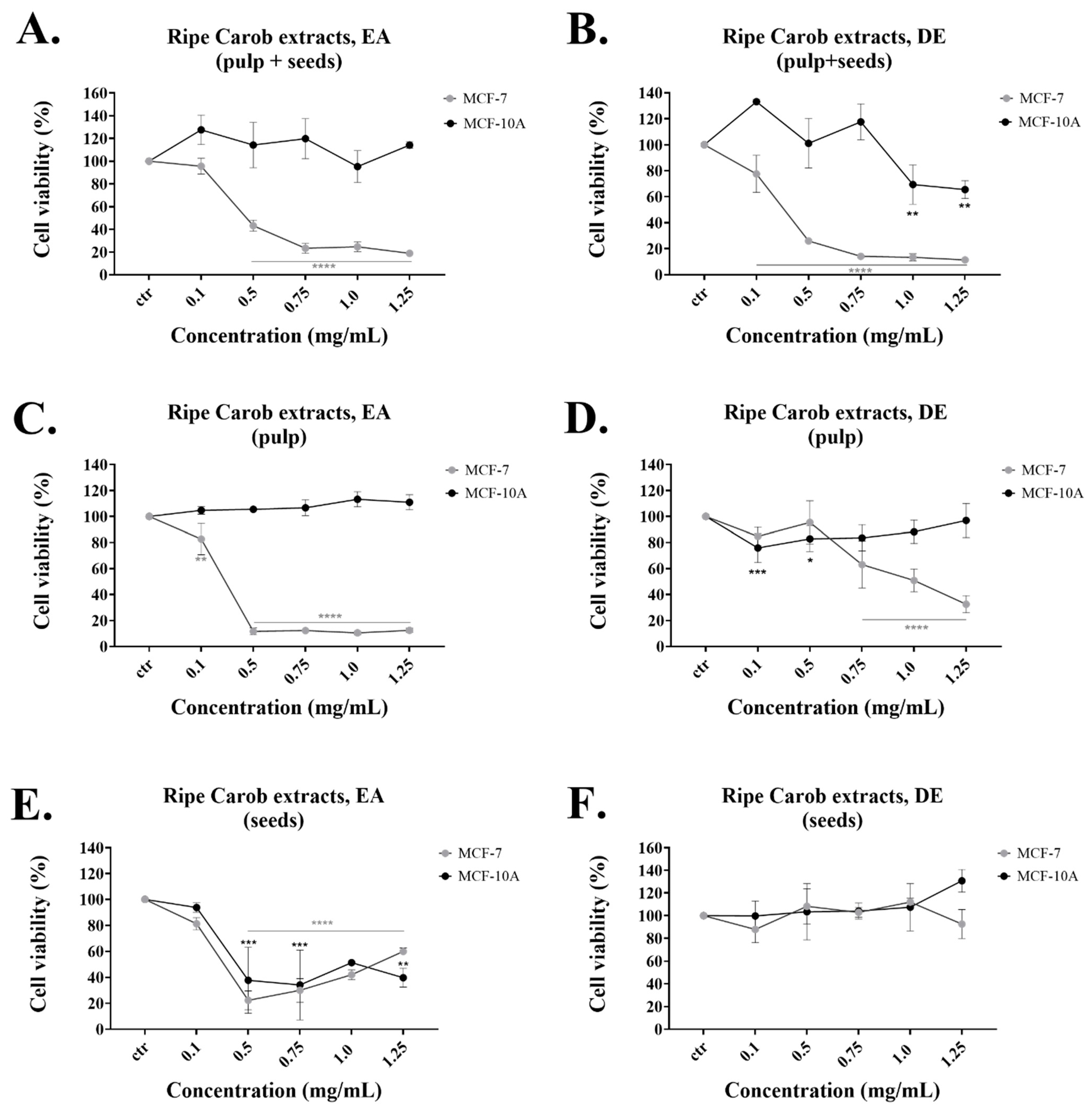
The DE extract from the “ripe pulp + seeds” had the lowest IC
50 and was selective for inhibiting the proliferation of cancer cells (
Table 1 and
Figure 1B). Even though the DE extract from the unripe “pulp + seeds” exerted high cell growth inhibition on the MCF-7 cell line, it also inhibited the growth of the normal MCF-10A cells (
Table 3 and
Figure S2B). The reason behind this may be due to the variations in phenolic content based on the carob’s ripening stage [
35]. Research studies have reported that during maturity, the total phenolic content varies. The unripe carobs contain higher total amount of polyphenols, but the ripe carobs contain the highest amounts of GA, possibly due to the degradation of gallotannins [
3,
35]. In fact, in this study, it was demonstrated that the highest TPC was found in extracts made using the unripe carobs (
Table 4). The DE ripe pulp had low TPC but was relatively selective for cancer cells (
Table 4 and
Figure 1D). The extracts made from the DE ripe seeds did not exert any anti-proliferative effects on either cell line. This is in agreement with our results from the TPC analysis (
Table 4) and other research studies, which reported that the carob seeds have lower amounts of polyphenols compared to the pulp [
36].
On the other hand, the EA extracts obtained from the “ripe” and “unripe pulp + seeds” both selectively reduced the viability of the MCF-7 cell line at the concentrations tested and also displayed relatively high TPC (
Figure 1A,
Figure S2A and
Table 4). The EA ripe seeds extract displayed a TPC of 14.74 mg/g but was not selective for either cell line while the EA ripe pulp had similar TPC content (11.57 mg/g) but was efficient only against the cancer cells. Finally, the EA ripe seeds had no anti-proliferative effect and insignificant TPC (
Figure S2E and
Table 4). Taken together, the current findings demonstrate the dose-dependent anti-proliferative impacts of the extracts examined, with the most potent being the “Ripe Pulp + Seeds” extracted with DE, followed by “Ripe Pulp” extracted with EA and the “Unripe Pulp + Seeds” extracted with EA. Based on the US National Cancer Institute (NCI) guidelines, a crude extract is generally considered to have high in vitro cytotoxic activity if the IC
50 value is ≤ 30 μg/mL [
37,
38]. Pezzuto et al. consider extracts with higher IC
50 values (100 μg/mL) to be cytotoxic and eligible candidates for further studies [
39], while several recently published papers [
40,
41,
42,
43,
44,
45] explore the biological activity of plant extracts with IC
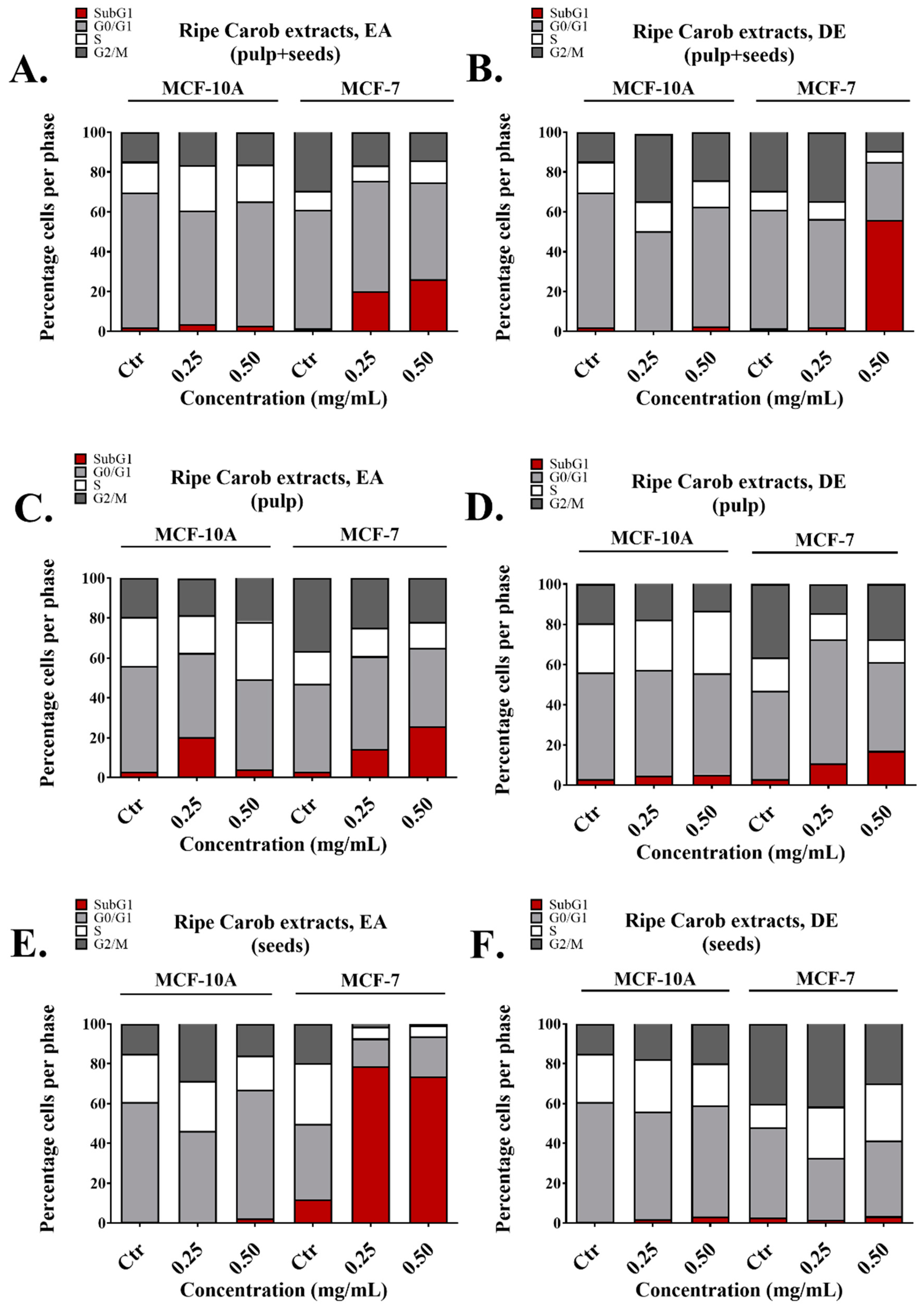
50 values higher than 200 μg/mL. Importantly, all the extracts made from the whole fruit using EA and DE induced apoptosis in MCF-7 cells with no significant effects on the normal breast cell line MCF-10A. This was also evident by the ability of these extracts to increase the subG1 fraction selectively in MCF-7 cells as evaluated with flow cytometry (
Figure 2A,B). The presence of the subG1 fraction is indicative of apoptosis [
46,
47]. Interestingly, in some cases, apoptosis induction was measured around 20% (
Figure 2), while cell viability as measured by the MTT assay, decreased by almost 80% (
Figure 1). This indicates that other forms of cell death, such as necrosis, may contribute to the observed anti-cancer effects of the carob extracts; Annexin V/Propidium iodide staining followed by flow cytometry as well as cell morphology as observed by microscopy is necessary to distinguish between apoptotic and necrotic death potentially induced by effective extracts. In addition, even though the EA extract from ripe pulp displayed potent anti-proliferative capacity (
Figure 1C,
Table 2), it also increased the subG1 fraction of normal cells (
Figure 2C). Future studies should include the sub-fractionation of crude carob extracts in an effort to improve their IC
50 values against cancer cells and screening of the crude extracts and their sub-fractions against a panel of cancer and normal cell lines to evaluate their effectiveness in a range of different cancer types.
Interestingly, the EA ripe seeds extract caused a significant increase of the subG1 phase selectively in cancer cells, revealing that it may have anti-cancer properties (
Figure 2E). Recent reports suggest that carob seed extracts could potentially be a source of bioactive anti-oxidant compounds [
48]. Even though the seeds, leaves and roots of different plant species have been mainly considered as food waste for years, they are now receiving attention for their bioactive constituents and their potential uses in the production of dietary supplements [
7,
49] or as sources of plant biostimulants [
50,
51]. Further characterization of the chemical composition of extracts derived from waste plant matrices is needed to reveal their full potential as possible sources of functional compounds [
52].
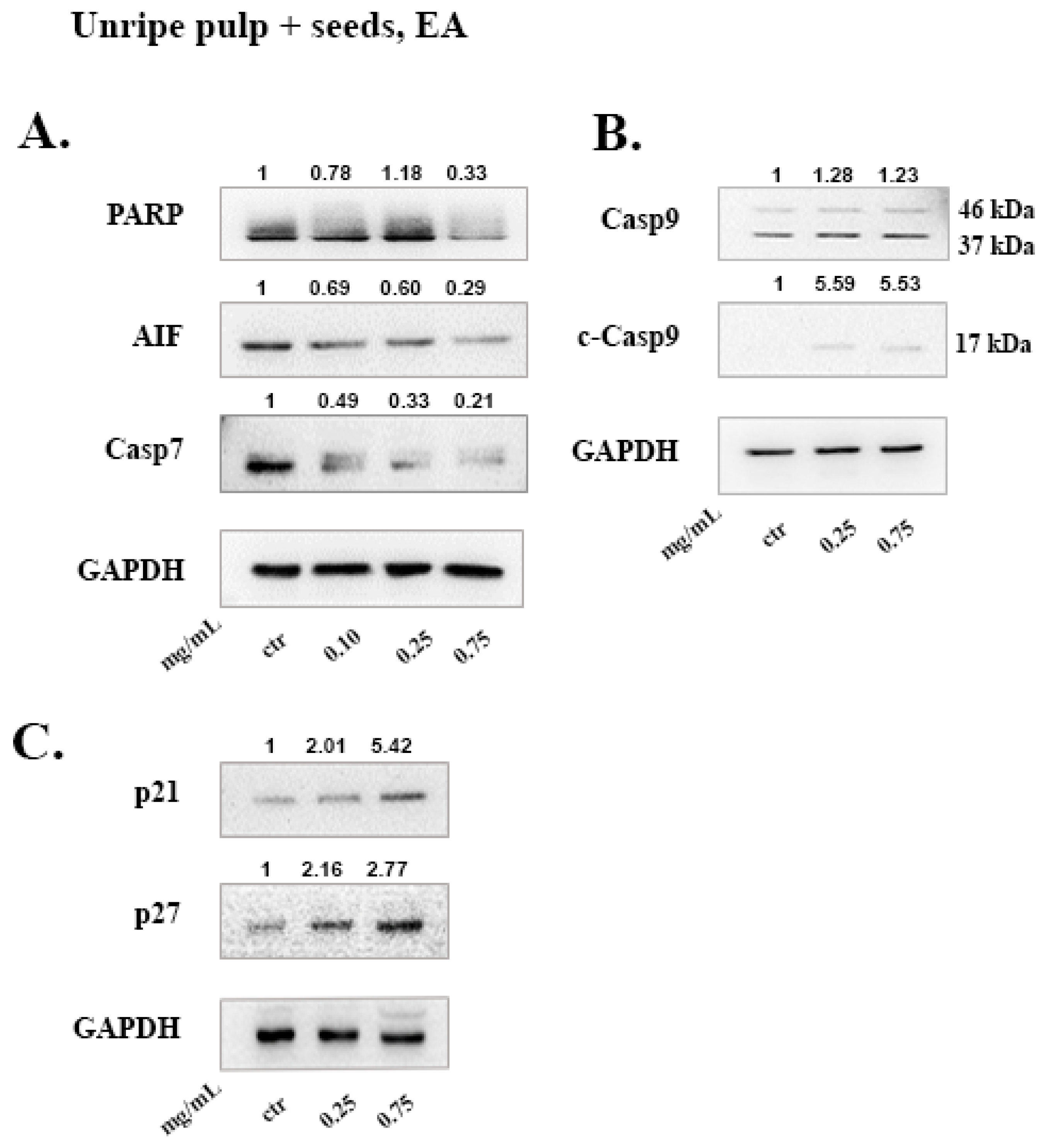
To evaluate the anti-oxidant activity of carob extracts, the DE and EA extracts were further analyzed using the DPPH assay. The most effective anti-oxidant extract was found to be the EA unripe pulp + seeds, with an EC
50 of 11.51 μg/mL and a TPC of 140.41 mg/g as determined using the Folin–Ciocalteu method. This is in agreement with a recent study by Kyriacou et. al, that highlighted the anti-oxidant potential of immature carob fruits [
3]. The study reported that during maturity, there was a swift decline in the fruit total phenolic content, catechins, tannins and flavonol glycosides, which resulted in the loss anti-oxidant capacity from the ripe fruit pulp extracts.
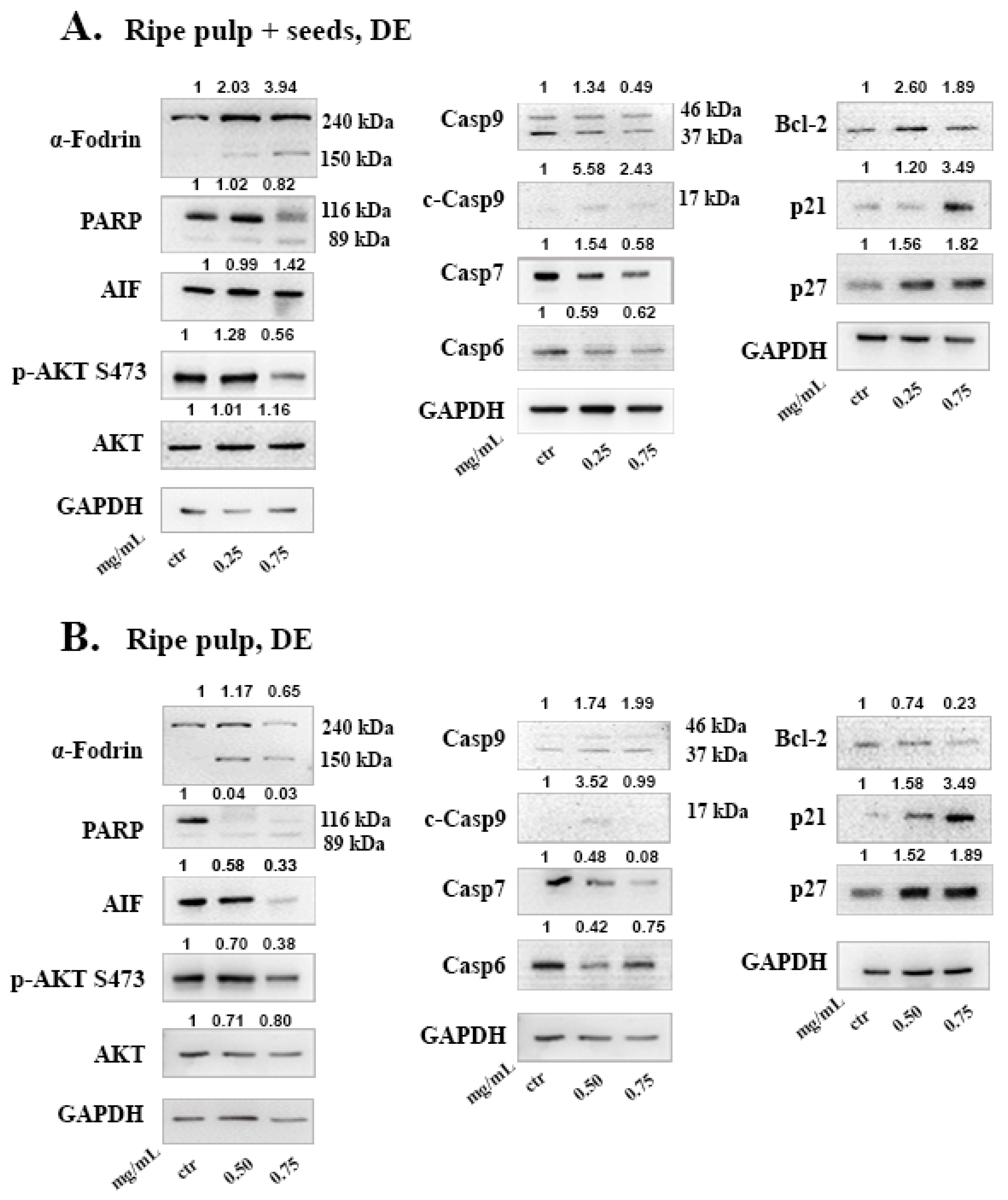
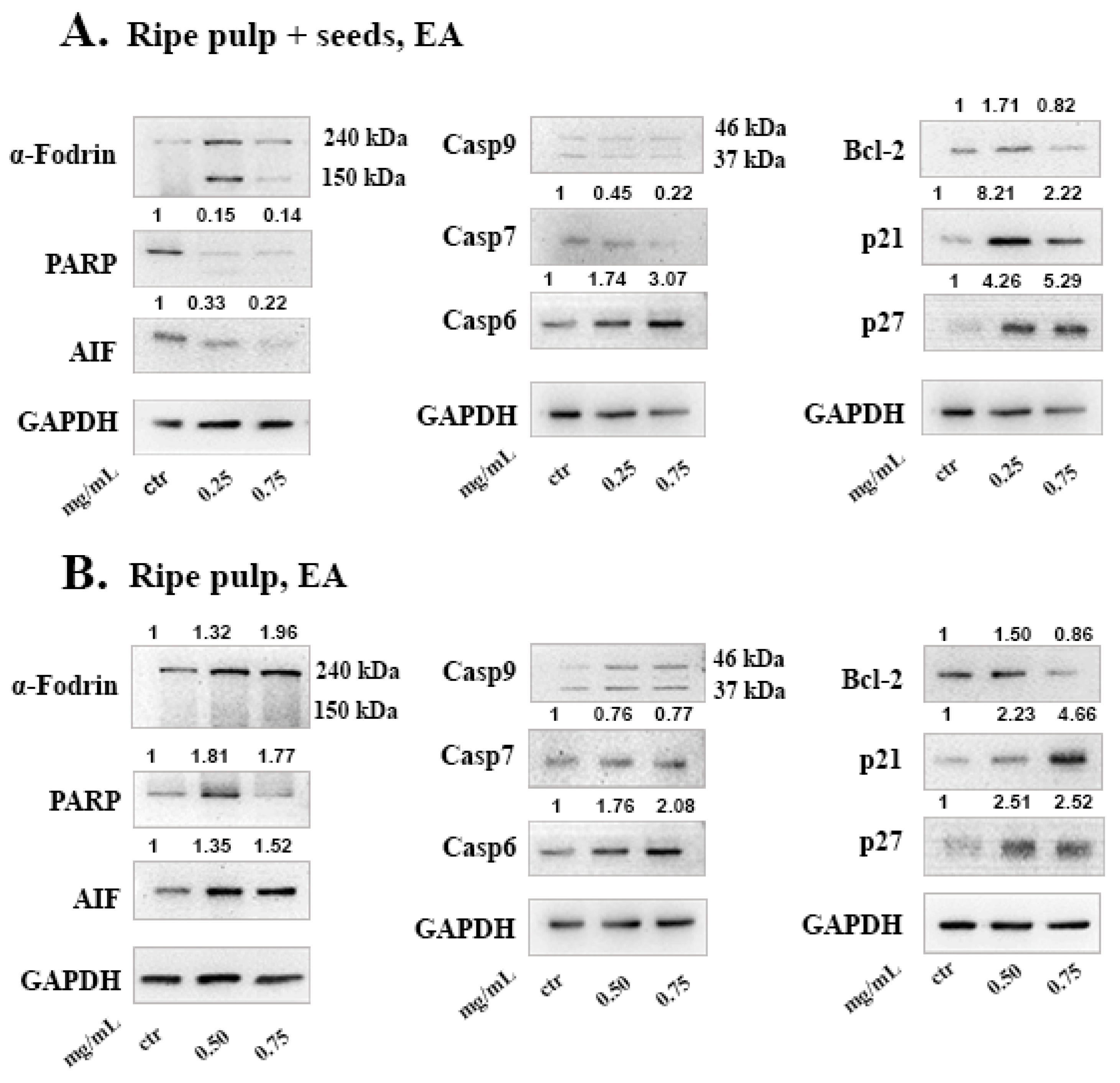
To investigate the mechanism of cell death induced by the carob extracts, we monitored the potential activation of key proteins involved in major apoptotic pathways. Caspase-9 functions in the intrinsic pathway of apoptosis and it is activated following cytochrome c release from the mitochondria. Once Caspase-9 is activated it can trigger the activation of the effector caspases -3, -6 and -7. Subsequently, effector caspases can cleave various substrates such as PARP and α-Fodrin resulting in cell disruption and eventually cell death [
22]. Caspase-7 takes on the role of the main effector caspase, when caspase-3 is missing [
53]. In caspase-3 deficient MCF-7 cells, we show that, DE “ripe pulp + seeds” and “ripe pulp” treatment induces the proteolysis of α-Fodrin accompanied by a decrease of the full-length protein expression of Caspase-7 (
Figure 4 and
Figure 5). PARP expression was decreased following treatment with all of the carob extracts at selected concentrations. Additionally, the EA unripe pulp + seeds, DE “ripe pulp + seeds” and “ripe pulp” extracts induced the cleavage of caspase-9. Importantly, Bcl-2, an anti-apoptotic protein, was decreased following treatment with the DE ripe pulp, EA “ripe pulp + seeds” and “ripe pulp” extracts. However, Bcl-2 expression did not decrease following treatment with the EA unripe pulp + seeds (data not shown) and DE ripe pulp + seeds extracts, suggesting that other protein members of the Bcl-2 family may be involved in inducing MOMP.
Following treatment with the carob extracts, AIF was decreased from its 67 kDa form which indicates its proteolysis to its soluble form (57 kDa), with the exception of EA ripe pulp treatment which resulted in increased protein levels of AIF. This indicates the accumulation and subsequent proteolysis of AIF to its soluble form and the involvement of the caspase-independent mechanism of apoptosis [
54]. In addition, we showed that the DE carob extracts inhibited the activation of the AKT pathway. Enhanced activation of AKT which occurs mainly via the phosphorylation at Ser473, leads to suppression of apoptosis [
55,
56,
57]. The decrease in AKT phosphorylation may explain the observed apoptosis induced by the carob extracts. The cell cycle inhibitors, p21 and p27, that are targets of the AKT pathway, were found to be upregulated following treatment with all of the carob extracts, indicative of inhibition of proliferation [
58].
Carob polyphenols have attracted scientific interest due to their health promoting effects. GA, the most abundant phenolic acid found in carobs, displays anti-cancer effects by inducing cell cycle arrest, and apoptosis via activating the caspase-dependent pathway and ROS generation [
59]. Previous studies have reported that GA was found at the highest concentration in the pulp compared to seeds. Indeed, the total phenolic content of GA in the pulp has been estimated to be between 0.45 and 53.7 mg equivalents/g of dry extract [
4] while in the seeds it has been estimated to be 0.19 mg equivalents/g of dry extract [
36,
60]. Based on the LC-MS analysis (
Table 5), Myricetin had the highest concentration in both tested samples from the ripe whole fruit; the DE extract had the highest levels of Myricetin and also displayed better IC50 values than the EA extract (
Table 1). Myricetin has been previously found to induce apoptosis in MCF-7 and triple negative MDA-MB-231 breast cancer cells [
61,
62]. Naringenin that was also detected in high levels in both extracts, has been found to induce tumor cell death and inhibit angiogenesis in malignant melanoma while it affected inflammatory and apoptosis pathways to inhibit migration of breast cancer cells [
63,
64]. Interestingly, in the DE extract, Kaempferol was approximately 5 times the concentration of that in EA. Kaempferol has been reported to induce apoptosis in breast cancer cell lines in vitro, through downregulation of Bcl-2 and cleavage of PARP [
65,
66], similarly to the carob extracts (
Figure 4A and
Figure 5A). Even though Myricetin, Naringenin and Kaempferol were detected in high levels in the examined extracts and these compounds have been found to induce apoptosis in cancer cells by previous studies, it is not clear to what extend they contribute to the anti-cancer effects observed here. Sub-fractionation of the most potent extracts and thorough investigation of their mechanism of action will reveal whether these effects may be attributed to polyphenols or other constituents.
Overall, our results showed that the anti-proliferative capacity of carob extracts varied with the stage of maturity of carobs and the solvent used for the extraction process. Regarding the ripe carobs, our results demonstrated that the DE and EA whole ripe extracts were highly specific in reducing the viability of MCF-7 cells compared to seeds, pulp, EtOH and Aqueous extracts. Previous studies showed that polyphenols are more soluble in solvents less polar than water [
28], suggesting that the DE and EA solvent extraction was able to dissolve higher amounts of polyphenols compared to EtOH and Aqueous extraction. These findings could explain the anti-proliferative effects observed using the DE and EA whole ripe carob extracts compared to the other solvents used.
4. Materials and Methods
4.1. Plant Material
Carobs were collected from a single tree growing near Delikipos village bearing fruits typical of the predominate local landrace “Tillyria”. Delikipos is a semi-mountainous area (300 m altitude) located in the central part of the island within one of the traditional centers of carob cultivation. Agro-environmental conditions at the Delikipos area favors the production of carobs with higher concentrations of condensed tannins, total phenolics, organic acids and other secondary metabolites compared to carobs produced to the coastal zone of the island [
67].
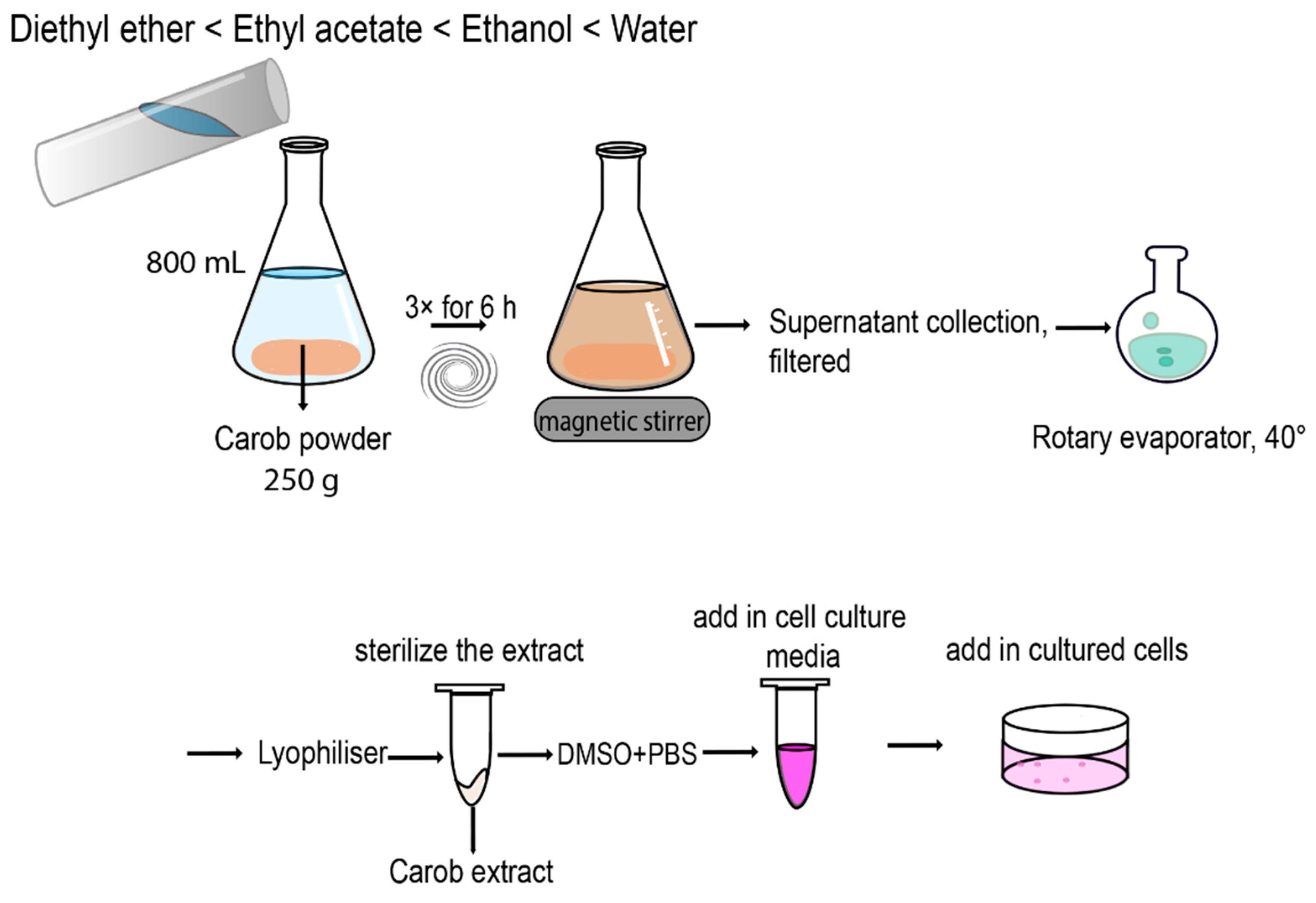
4.2. Extraction Methods
The carobs and by-products (seeds and pulp) were cut in small pieces and grounded using a food processor (Thermomix). Next, the samples were immersed in liquid nitrogen and freeze-dried under pressure for 24 h. The lyophilized samples (seeds, pulp and whole carob) were grounded in a food processor to produce a fine powder and sieved through a stainless-steel mesh. Subsequently, 250 g of carob powder obtained from the whole ripe carobs was successively extracted using a series of solvents of increasing polarity (3 × 800 mL, for each solvent for 6 h) under magnetic stirring at room temperature. The order of polarity is as follows: Diethyl ether (DE) < Ethyl acetate (EA) < Ethanol (EtOH) < Water.
For the aqueous extract the procedure was performed once. The supernatants of each solvent were combined and filtered using a Whatman No. 1 filter paper. The Aqueous extract was lyophilized for 24 h while the DE, EA and EtOH extracts were placed into a round bottom flask and evaporated under pressure at 40 °C using the rotary evaporator, lyophilized for 24 h and stored at −20 °C for future use. For the extraction of the pulp, seeds, and pulp + seeds, from the ripe and unripe carobs we used the DE and EA solvents only and as described above (
Figure 7).
The extract yields were calculated as follows:
where A is the weight of the extract and B is the weight of the initial carobs.
The yield of extraction is shown in
Table 6.
Among the four solvents used in the extraction process, the DE and EA extracts were found to possess the strongest anti-proliferative effects in established cell lines (
Table 1). Based on these results, we used only these two solvents for the extraction of unripe carobs (pulp + seeds, pulp or seeds). The yield of extraction of unripe carobs is shown in
Table 7.
4.3. Cell Culture and Reagents
MCF-7 and MCF-10A cell lines were obtained from the American Type Culture Collection (ATCC) (Manassas, VA, USA). MCF-7 breast cancer cell line was cultured in DMEM supplemented with 10% Fetal Bovine Serum (FBS) and 1% antibiotic/antimycotic. MCF-10A immortalized breast cell line was cultured in DMEM F12 supplemented with 20 ng/mL EGF, 100 ng/mL Cholera Toxin, 500 ng/mL Hydrocortizone, 10 μg/mL Insulin, 5% Horse Serum (HS) and 1% antibiotic/antimycotic. DMEM, FBS, HS, antibiotic/antimycotic and trypsin were purchased from Gibco, Invitrogen. EA, DE and EtOH used for the preparation of carob extracts were purchased from Sigma-Aldrich (St. Louis, MO, USA). PARP, AKT, p-AKT, Bcl-2, AIF and Caspases-6, -7, -8, -9 antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA). GAPDH, p21, p27 and α-Fodrin antibodies were purchased from Santa Cruz Biotechnology Inc. All polyphenolic standards are analytical grade. Apigenin, Quercetin, Sinapic acid, Caffeic acid, Naringenin and Gallic acid were purchased from Sigma-Aldrich (St. Louis, MO, USA). Catechin, Myricetin and Quercitrin were supplied from HWI ANALYTIK GMBH (Rulzheim, Germany). Kaempferol and Rutin were purchased from LGC Standards GmbH (Luckenwalde, Germany) and PhytoLab GmbH and Co (Vestenbergsgreuth, Germany), respectively. All other reagents were purchased from Sigma Aldrich.
4.4. MTT Assay
The MTT assay was performed as described previously [
68] with a few modifications. A total of 5 × 10
4 cells were seeded per well of a 96-well plate. The breast cell lines MCF-7 and MCF-10A were incubated overnight to allow for cell attachment and recovery. Cells were treated with increasing doses of Carob extracts (0.1–2.5 mg/mL) and incubated for 48 h at 37 °C. Cell viability was measured using the MTT 3-(4,5-dimethylthiazol-2-yl)-2,5-mono tetrazolium bromide assay. At the end of each incubation period, 20 μL of MTT dye (1 mg/mL; Sigma St. Louis, MO, USA) was added in each well and the plate was incubated at 37 °C for 4 h. Subsequently, the plates were immediately read on a microplate reader (Wallac, PerkinElmer, MA, USA) at 570 nm. Absorbance was proportional to the number of viable cells per well. Percentage of cell viability in each group was calculated after normalization to its own control.
4.5. Cell Cycle Analysis
Following incubation, cells were treated with different concentrations (as described in the figure legends) of carob extracts for 48 h. They were harvested, fixed in 70% ethanol, incubated with the Propidium Iodide (PI) staining solution (containing 1 mg/mL PI and 100 μg/mL Rnase) for 30 min at 37 °C and analyzed for DNA content using the Guava EasyCyte flow cytometer and the GuavaSoft analysis software (Millipore, Watford, UK).
4.6. Western Blotting
Following incubation with selected carob extracts at different concentrations (as indicated in the figures), cells were washed with ice-cold PBS and lysed in RIPA buffer (150 mM NaCl, 50 mM Tris, 5 mM EDTA [Na2], 1% (
v/
v) Triton X-100, 1% (
w/
v) deoxycholate (24 mM), 0.1% (
w/
v) SDS (35 mM)) containing protease and phosphatase inhibitors (Complete Mini, Roche), in order to achieve the cleavage of the cell membranes. The total cellular extracts of the proteins were collected and the protein levels in each sample were measured by using the Bradford method and run on SDS page electrophoresis as described elsewhere [
69]. The intensity values from the densitometry analysis of Western blots were normalized against the corresponding loading control using ImageJ analysis software (NIH).
4.7. Folin–Ciocalteu Method for the Quantification of Total Phenolic Content
The total phenolic content of the extracts was determined using the Folin–Ciocalteu method as previously described [
70] with a few modifications. Folin–Ciocalteu was used as the reagent and GA as the standard. Initially, a set of GA standard solutions at concentrations of 0.010, 0.025, 0.050, 0.100, 0.250 and 0.500 μg/mL were prepared and used to construct a calibration curve. 100 μL of extract (concentration 2 mg/mL) or standard solution was added to a vial. Subsequently, 4.5 mL of distilled water and 100 μL Folin–Ciocalteu reagent were added, and the vial was shaken vigorously. After 3 min, 300 μL of Na
2CO
3 (2%) solution was added and the mixture was allowed to react for 2 h shaking occasionally. The absorbance was measured at 760 nm on a UV spectrophotometer (UV-1700 series, Shimadzu, Kyoto, Japan). The total phenolic content was expressed as GA equivalents (mg GAE/g extract).
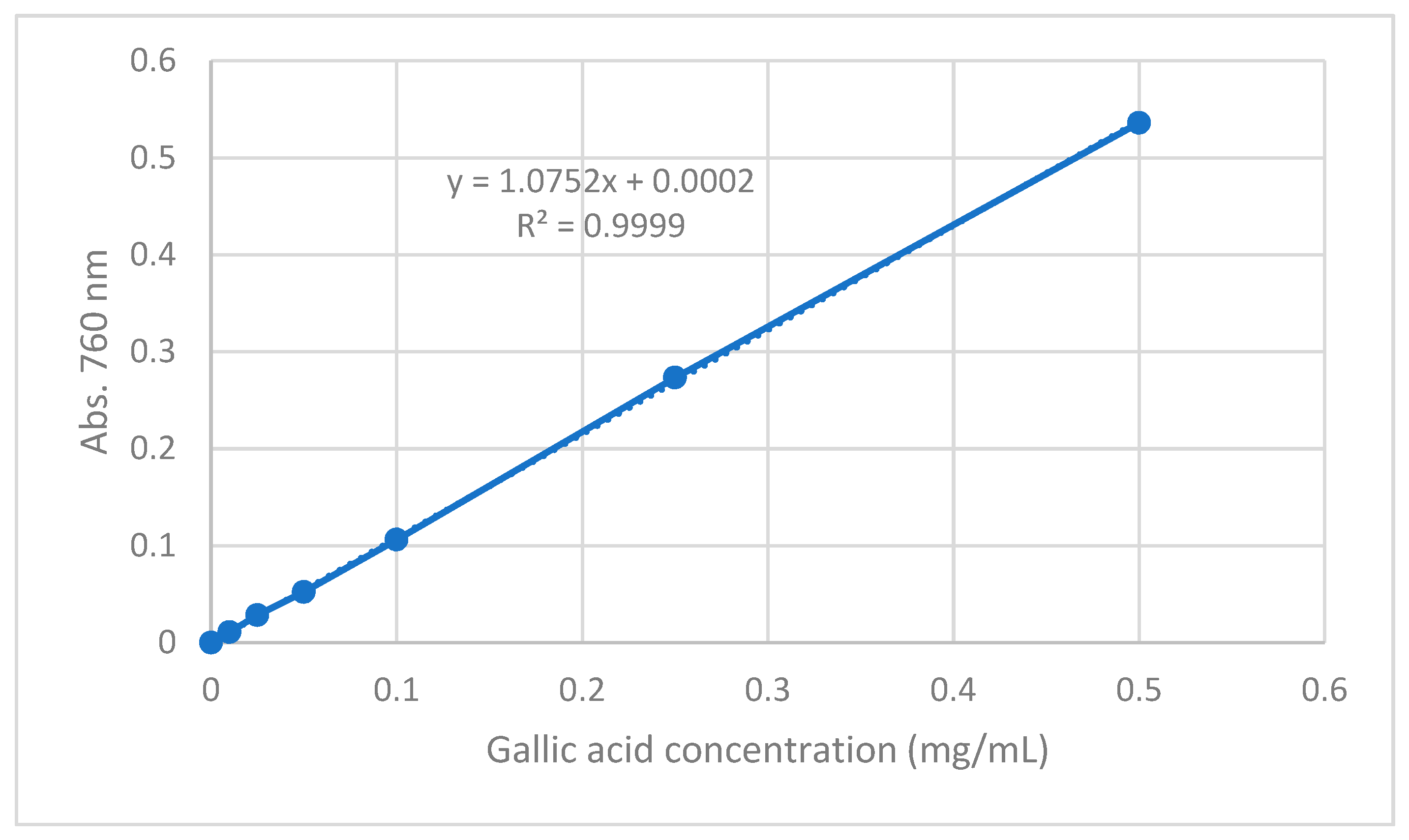
The linearity of standard calibration curve was evaluated by linear regression analysis. The standard calibration curve was calculated by the least squares regression method to calculate the calibration equation and the determination coefficient (R
2).
Figure 8 shows the standard calibration curve of the F–C method. A linear correlation was found between absorbance of the blue complex at 760 nm and concentration of gallic acid in the range 0.010–0.500 mg/mL. The determination coefficient obtained from the linear regression was 0.9999, indicating excellent linear correlation between the data. From the slope of the calibration curve, LOD and LOQ were established. The LOD and LOQ for the F-C method were 0.0055 and 0.166 mg GAE/mL, respectively.
4.8. DPPH (2,2-Diphenyl-1-picryl-hydrazyl-hydrate) Radical Scavenging Assay
The DPPH radical scavenging assay was performed according to the protocol reported by Sarikurkcu et al. (2009) [
71] with some modifications. 1 mL of each extract solution of concentrations 0.001, 0.002, 0.005, 0.01, 0.05, 0.1, 0.5 and 1 mg/mL were prepared and added to a 1 mL of DPPH fresh radical solution in methanol (0.2 mM). A control sample was prepared, which contained 1 mL H
2O and 1 mL DPPH. After the mixtures were shaken vigorously and left in the dark for 30 min at room temperature, the absorbance was measured at 517 nm (Shimadzu, UV-1700). Radical scavenging capacity was expressed as a percentage effect (I%) and calculated using the following equation:
where
Acontrol is the absorbance of the control and
Asample is the absorbance of the extract.
Antiradical curves were plotted referring to concentration on the x axis and their relative scavenging capacity on the y axis. The EC50 values were calculated using the Origin program.
4.9. LC-MS Analysis
Analyte identification criteria were relative retention time within ±0.05 min of the mean calibrator retention time and SIM mode of parent ion of each compound. For quantitative determination, calibration curves were constructed by plotting the peak areas of polyphenols against their concentrations, which were in the range 1–15 ng/mL for Apigenin, Gallic acid and Ferulic acid, 3–25 ng/mL for Rutin and Kaempferol, 10–50 ng/mL for Naringenin and 15–100 ng/mL for Myricetin. The obtained regression equations revealed a linear relationship between the peak area of each polyphenol species and their concentration, with corresponding correlation coefficients higher than 0.995. The limit of detection (LOD) and limit of quantification (LOQ) were calculated based on the standard deviation of the response and the slope (
Table S1). The detection limits were expressed as:
where: σ = the standard deviation of the response at low concentrations. Slope = the slope of the calibration curve.
DE and EA ripe pulp + seeds extracts were dissolved in methanol (LC-MS grade) to obtain the final concentration of 250 ppb, sonicated and filtered through 0.45 μm PTFE syringe filters prior to analysis.
HPLC-MS analysis was performed according to a previously reported method with some modifications [
72]. Analyses were performed using an Agilent Technologies Infinity liquid 1260 chromatographic system, consisting of an autosampler, a column thermostat and a binary solvent management system. MS data were acquired via positive electrospray ionization (ESI) in SIM mode. The chromatographic separation was accomplished using a Venusil XBP C18 column (150 × 4.6 mm, 5 μm) with a pre-column of the same material. A binary A/B gradient elution was used with solvent A: Milli Q water with 0.01% TFA, and solvent B: ACN. The gradient program was as follows: initial conditions were 100% A, raised to 98% B over the first 30 min. For re-equilibration of the system before the next injection, 15 min post time was used. Ten microliters were injected into the system with a flow rate of 0.8 mL/min and column temperature at 25 °C.
4.10. Statistical Analysis
Results for continuous variables were presented as Mean Standard Error. Two-group differences in continuous variables were assessed by the unpaired t-test. P-values are two-tailed with confidence intervals 95%. Statistical analysis was performed by comparing treated samples with untreated control. All statistical tests were conducted using Prism software version 8.0 (Graphpad, San Diego, CA, USA).



 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}