
Induction of G2/M Cell Cycle Arrest via p38/p21Waf1/Cip1-Dependent Signaling Pathway Activation by Bavachinin in Non-Small-Cell Lung Cancer Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Cytotoxic Effect of Bavachinin on Non-Small-Cell Lung Cancer Cells
2.2. Induction of G2/M Cell Cycle Arrest in Bavachinin-Inhibited Non-Small Cell Lung Cancer Cell Viability
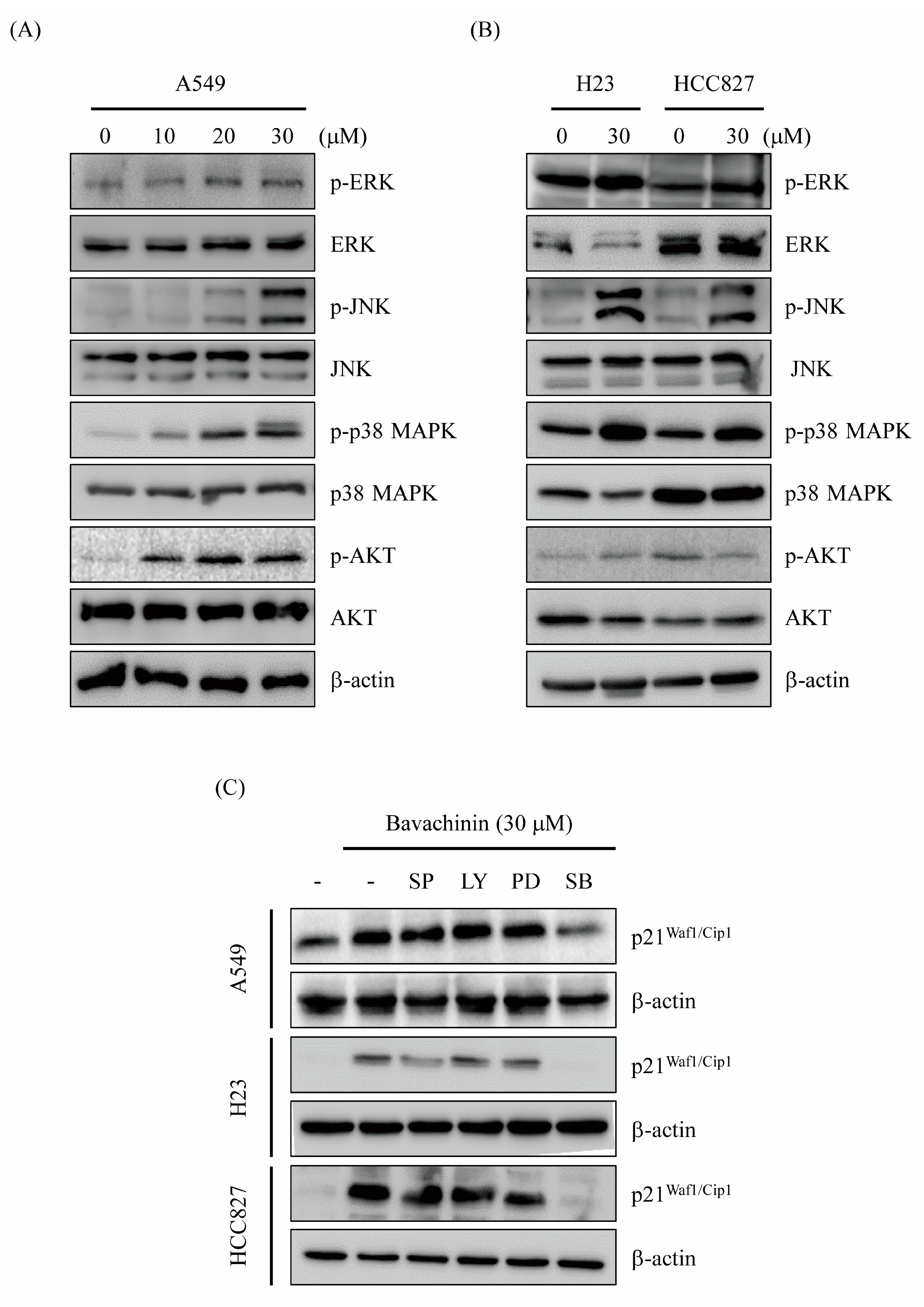
2.3. Induced G2/M Cells Accumulation via Bavachinin through p21Waf1/Cip1-Mediated Signaling Pathway
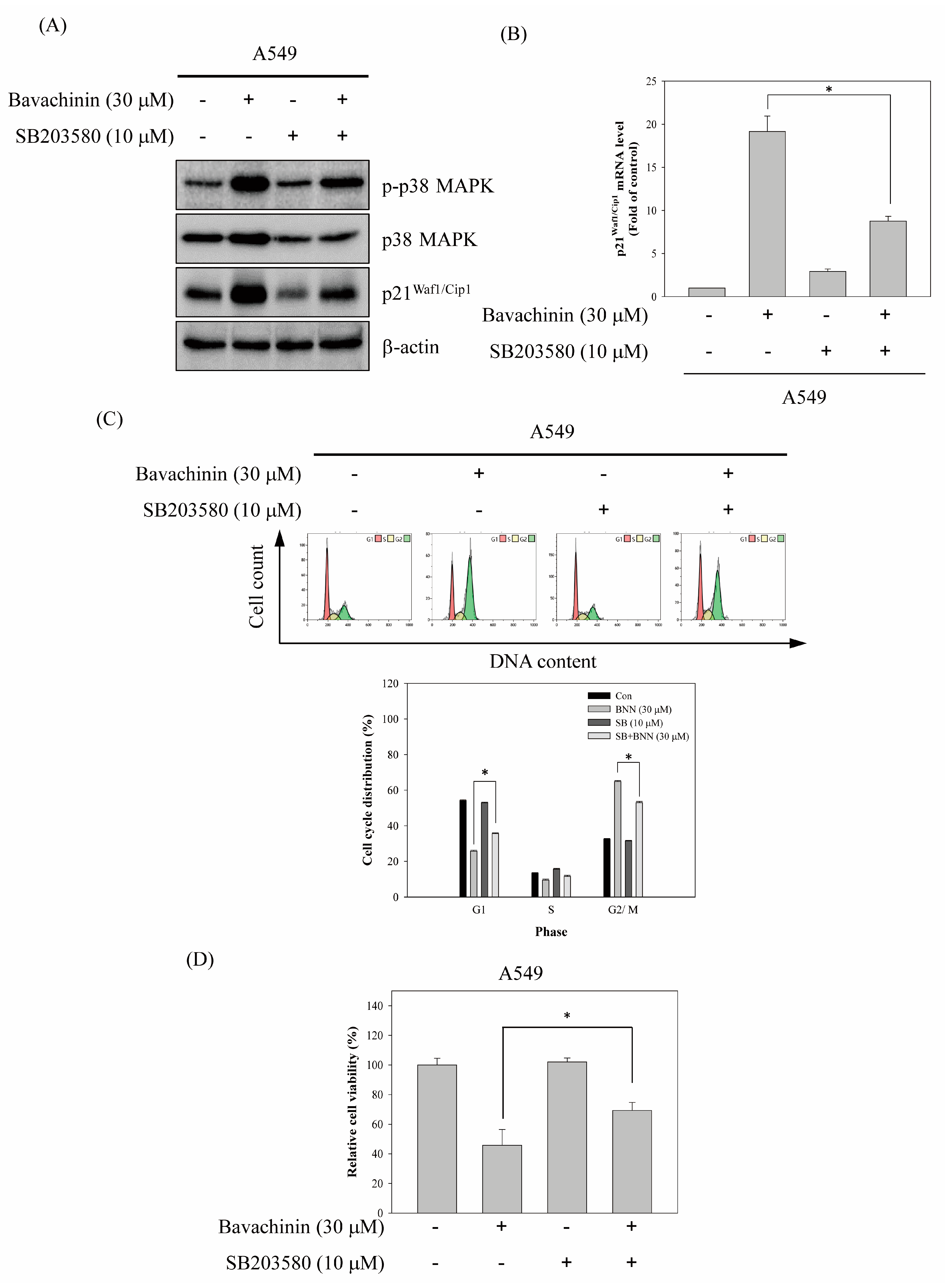
2.4. Up-Regulation of p21Waf1/Cip1 Expression by Bavachinin through p38 MAPK-Mediated Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture and Cell Viability Assay
4.3. Colony Formation Assay
4.4. Cell Cycle Distribution Analysis
4.5. Western Blot Analysis
4.6. Immunoprecipitation
4.7. Quantitative Reverse-Transcription Polymerase Chain Reaction
4.8. Small Interfering RNA Transfection
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Ethics Statements
References
- Inamura, K. Lung Cancer: Understanding Its Molecular Pathology and the 2015 WHO Classification. Front. Oncol. 2017, 7, 193. [Google Scholar] [CrossRef] [Green Version]
- Blandin Knight, S.; Crosbie, P.A.; Balata, H.; Chudziak, J.; Hussell, T.; Dive, C. Progress and prospects of early detection in lung cancer. Open Biol. 2017, 7, 170070. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Kaldis, P. Cdks, cyclins and CKIs: Roles beyond cell cycle regulation. Development 2013, 140, 3079–3093. [Google Scholar] [CrossRef] [Green Version]
- Sherr, C.J.; Roberts, J.M. CDK inhibitors: Positive and negative regulators of G1-phase progression. Genes Dev. 1999, 13, 1501–1512. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Dutta, A. p21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Gazzeri, S.; Gouyer, V.; Vour’ch, C.; Brambilla, C.; Brambilla, E. Mechanisms of p16INK4A inactivation in non small-cell lung cancers. Oncogene 1998, 16, 497–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betticher, D.C.; Heighway, J.; Hasleton, P.S.; Altermatt, H.J.; Ryder, W.D.; Cerny, T.; Thatcher, N. Prognostic significance of CCND1 (cyclin D1) overexpression in primary resected non-small-cell lung cancer. Br. J. Cancer 1996, 73, 294–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.; Inoue, S.; Umemura, T.; Moriya, J.; Arakawa, M.; Nagashima, K.; Kato, H. Cyclin D1, p16 and retinoblastoma gene product expression as a predictor for prognosis in non-small cell lung cancer at stages I and II. Lung Cancer 2001, 34, 207–218. [Google Scholar] [CrossRef]
- Eymin, B.; Gazzeri, S. Role of cell cycle regulators in lung carcinogenesis. Cell Adh. Migr. 2010, 4, 114–123. [Google Scholar] [CrossRef]
- Shoji, T.; Tanaka, F.; Takata, T.; Yanagihara, K.; Otake, Y.; Hanaoka, N.; Miyahara, R.; Nakagawa, T.; Kawano, Y.; Ishikawa, S.; et al. Clinical significance of p21 expression in non-small-cell lung cancer. J. Clin. Oncol. 2002, 20, 3865–3871. [Google Scholar] [CrossRef]
- Esposito, V.; Baldi, A.; Tonini, G.; Vincenzi, B.; Santini, M.; Ambrogi, V.; Mineo, T.C.; Persichetti, P.; Liuzzi, G.; Montesarchio, V.; et al. Analysis of cell cycle regulator proteins in non-small cell lung cancer. J. Clin. Pathol. 2004, 57, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Soria, J.C.; Jang, S.J.; Khuri, F.R.; Hassan, K.; Liu, D.; Hong, W.K.; Mao, L. Overexpression of cyclin B1 in early-stage non-small cell lung cancer and its clinical implication. Cancer Res. 2000, 60, 4000–4004. [Google Scholar] [PubMed]
- Singhal, S.; Amin, K.M.; Kruklitis, R.; DeLong, P.; Friscia, M.E.; Litzky, L.A.; Putt, M.E.; Kaiser, L.R.; Albelda, S.M. Alterations in cell cycle genes in early stage lung adenocarcinoma identified by expression profiling. Cancer Biol. Ther. 2003, 2, 291–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, F.; Khan, G.N.; Asad, M. Psoralea corylifolia L: Ethnobotanical, biological, and chemical aspects: A review. Phytother. Res. 2018, 32, 597–615. [Google Scholar] [CrossRef] [PubMed]
- Nepal, M.; Choi, H.J.; Choi, B.Y.; Kim, S.L.; Ryu, J.H.; Kim, D.H.; Lee, Y.H.; Soh, Y. Anti-angiogenic and anti-tumor activity of Bavachinin by targeting hypoxia-inducible factor-1alpha. Eur. J. Pharmacol. 2012, 691, 28–37. [Google Scholar] [CrossRef]
- Ge, L.N.; Yan, L.; Li, C.; Cheng, K. Bavachinin exhibits antitumor activity against nonsmall cell lung cancer by targeting PPARgamma. Mol. Med. Rep. 2019, 20, 2805–2811. [Google Scholar] [PubMed]
- Wang, S.; Wang, M.; Wang, M.; Tian, Y.; Sun, X.; Sun, G.; Sun, X. Bavachinin Induces Oxidative Damage in HepaRG Cells through p38/JNK MAPK Pathways. Toxins 2018, 10, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Ghosh, B.; Chakraborty, T.; Roy, S. Bavachinin mitigates DMH induced colon cancer in rats by altering p53/Bcl2/BAX signaling associated with apoptosis. Biotech. Histochem. 2021, 96, 179–190. [Google Scholar] [CrossRef]
- Dong, X.; Zhu, Y.; Wang, S.; Luo, Y.; Lu, S.; Nan, F.; Sun, G.; Sun, X. Bavachinin inhibits cholesterol synthesis enzyme FDFT1 expression via AKT/mTOR/SREBP-2 pathway. Int. Immunopharmacol. 2020, 88, 106865. [Google Scholar] [CrossRef]
- Hung, S.Y.; Lin, S.C.; Wang, S.; Chang, T.J.; Tung, Y.T.; Lin, C.C.; Ho, C.T.; Li, S. Bavachinin Induces G2/M Cell Cycle Arrest and Apoptosis via the ATM/ATR Signaling Pathway in Human Small Cell Lung Cancer and Shows an Antitumor Effect in the Xenograft Model. J. Agric. Food Chem. 2021, 69, 6260–6270. [Google Scholar] [CrossRef]
- Yu, S.Y.; Liao, C.H.; Chien, M.H.; Tsai, T.Y.; Lin, J.K.; Weng, M.S. Induction of p21(Waf1/Cip1) by garcinol via downregulation of p38-MAPK signaling in p53-independent H1299 lung cancer. J. Agric. Food Chem. 2014, 62, 2085–2095. [Google Scholar] [CrossRef]
- Lee, S.R.; Park, J.H.; Park, E.K.; Chung, C.H.; Kang, S.S.; Bang, O.S. Akt-induced promotion of cell-cycle progression at G2/M phase involves upregulation of NF-Y binding activity in PC12 cells. J. Cell. Physiol. 2005, 205, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.P.; Ho, P.Y.; Liang, Y.C.; Ho, Y.S.; Lee, W.S. Involvement of the JNK activation in terbinafine-induced p21 up-regulation and DNA synthesis inhibition in human vascular endothelial cells. J. Cell. Biochem. 2009, 108, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Ciccarelli, C.; Marampon, F.; Scoglio, A.; Mauro, A.; Giacinti, C.; De Cesaris, P.; Zani, B.M. p21WAF1 expression induced by MEK/ERK pathway activation or inhibition correlates with growth arrest, myogenic differentiation and onco-phenotype reversal in rhabdomyosarcoma cells. Mol. Cancer 2005, 4, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malumbres, M.; Barbacid, M. Mammalian cyclin-dependent kinases. Trends Biochem. Sci. 2005, 30, 630–641. [Google Scholar] [CrossRef]
- Schmidt, M.; Rohe, A.; Platzer, C.; Najjar, A.; Erdmann, F.; Sippl, W. Regulation of G2/M Transition by Inhibition of WEE1 and PKMYT1 Kinases. Molecules 2017, 22, 2045. [Google Scholar] [CrossRef] [Green Version]
- Niculescu, A.B., 3rd; Chen, X.; Smeets, M.; Hengst, L.; Prives, C.; Reed, S.I. Effects of p21(Cip1/Waf1) at both the G1/S and the G2/M cell cycle transitions: pRb is a critical determinant in blocking DNA replication and in preventing endoreduplication. Mol. Cell Biol. 1998, 18, 629–643. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Kato, T.; Kiyotani, K.; Tarhan, Y.E.; Saloura, V.; Chung, S.; Ueda, K.; Nakamura, Y.; Park, J.H. p53-independent p21 induction by MELK inhibition. Oncotarget 2017, 8, 57938–57947. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, T.; Roth, J.A. Induction of apoptosis in human lung cancer cells after wild-type p53 activation by methoxyestradiol. Oncogene 1997, 14, 379–384. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.W.; Lin, C.H.; Hsiao, T.H.; Lo, C.C.; Hsieh, C.Y.; Huang, C.C.; Sher, Y.P. Therapeutic effects of statins against lung adenocarcinoma via p53 mutant-mediated apoptosis. Sci. Rep. 2019, 9, 20403. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; He, G.; Nelman-Gonzalez, M.; Ashorn, C.L.; Gallick, G.E.; Stukenberg, P.T.; Kirschner, M.W.; Kuang, J. Regulation of Cdc25C by ERK-MAP kinases during the G2/M transition. Cell 2007, 128, 1119–1132. [Google Scholar] [CrossRef] [Green Version]
- Thornton, T.M.; Rincon, M. Non-classical p38 map kinase functions: Cell cycle checkpoints and survival. Int. J. Biol. Sci. 2009, 5, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, W.; Wang, Y.; Lu, J.; Chen, X. The Chemical Constituents and Bioactivities of Psoralea corylifolia Linn.: A Review. Am. J. Chin. Med. 2016, 44, 35–60. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Nam, J.W.; Song, Y.S.; Viswanath, A.N.; Pae, A.N.; Kil, Y.S.; Kim, H.D.; Park, J.H.; Seo, E.K.; Chang, M. Psoralidin, a coumestan analogue, as a novel potent estrogen receptor signaling molecule isolated from Psoralea corylifolia. Bioorg. Med. Chem. Lett. 2014, 24, 1403–1406. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Yun, X.; Tao, R.; Wang, Y.; Fan, G.; Zhu, Y.; Cai, T.; Zhu, Z.; Yan, C.; Gao, X. Bakuchiol exhibits anti-metastasis activity through NF-kappaB cross-talk signaling with AR and ERbeta in androgen-independent prostate cancer cells PC-3. J. Pharmacol. Sci. 2018, 138, 1–8. [Google Scholar] [CrossRef]
- Chen, Z.; Jin, K.; Gao, L.; Lou, G.; Jin, Y.; Yu, Y.; Lou, Y. Anti-tumor effects of bakuchiol, an analogue of resveratrol, on human lung adenocarcinoma A549 cell line. Eur. J. Pharmacol. 2010, 643, 170–179. [Google Scholar] [CrossRef]
- Hao, W.; Zhang, X.; Zhao, W.; Chen, X. Psoralidin induces autophagy through ROS generation which inhibits the proliferation of human lung cancer A549 cells. PeerJ 2014, 2, e555. [Google Scholar] [CrossRef] [Green Version]
- Panno, M.L.; Giordano, F. Effects of psoralens as anti-tumoral agents in breast cancer cells. World J. Clin. Oncol 2014, 5, 348–358. [Google Scholar] [CrossRef]
- Wang, X.; Peng, P.; Pan, Z.; Fang, Z.; Lu, W.; Liu, X. Psoralen inhibits malignant proliferation and induces apoptosis through triggering endoplasmic reticulum stress in human SMMC7721 hepatoma cells. Biol. Res. 2019, 52, 34. [Google Scholar] [CrossRef]
- Du, G.; Zhao, Y.; Feng, L.; Yang, Z.; Shi, J.; Huang, C.; Li, B.; Guo, F.; Zhu, W.; Li, Y. Design, Synthesis, and Structure-Activity Relationships of Bavachinin Analogues as Peroxisome Proliferator-Activated Receptor gamma Agonists. ChemMedChem 2017, 12, 183–193. [Google Scholar] [CrossRef]
- Lin, L.C.; Hsu, S.L.; Wu, C.L.; Hsueh, C.M. TGFbeta can stimulate the p(38)/beta-catenin/PPARgamma signaling pathway to promote the EMT, invasion and migration of non-small cell lung cancer (H460 cells). Clin. Exp. Metastasis 2014, 31, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Ai, X.; Mao, F.; Shen, S.; Shentu, Y.; Wang, J.; Lu, S. Bexarotene inhibits the viability of non-small cell lung cancer cells via slc10a2/PPARgamma/PTEN/mTOR signaling pathway. BMC Cancer 2018, 18, 407. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.H.; Sang, S.; Ho, C.T.; Lin, J.K. Garcinol modulates tyrosine phosphorylation of FAK and subsequently induces apoptosis through down-regulation of Src, ERK, and Akt survival signaling in human colon cancer cells. J. Cell Biochem. 2005, 96, 155–169. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pai, J.-T.; Hsu, M.-W.; Leu, Y.-L.; Chang, K.-T.; Weng, M.-S. Induction of G2/M Cell Cycle Arrest via p38/p21Waf1/Cip1-Dependent Signaling Pathway Activation by Bavachinin in Non-Small-Cell Lung Cancer Cells. Molecules 2021, 26, 5161. https://doi.org/10.3390/molecules26175161
Pai J-T, Hsu M-W, Leu Y-L, Chang K-T, Weng M-S. Induction of G2/M Cell Cycle Arrest via p38/p21Waf1/Cip1-Dependent Signaling Pathway Activation by Bavachinin in Non-Small-Cell Lung Cancer Cells. Molecules. 2021; 26(17):5161. https://doi.org/10.3390/molecules26175161
Chicago/Turabian StylePai, Jih-Tung, Ming-Wei Hsu, Yann-Lii Leu, Kuo-Ting Chang, and Meng-Shih Weng. 2021. "Induction of G2/M Cell Cycle Arrest via p38/p21Waf1/Cip1-Dependent Signaling Pathway Activation by Bavachinin in Non-Small-Cell Lung Cancer Cells" Molecules 26, no. 17: 5161. https://doi.org/10.3390/molecules26175161
APA StylePai, J. -T., Hsu, M. -W., Leu, Y. -L., Chang, K. -T., & Weng, M. -S. (2021). Induction of G2/M Cell Cycle Arrest via p38/p21Waf1/Cip1-Dependent Signaling Pathway Activation by Bavachinin in Non-Small-Cell Lung Cancer Cells. Molecules, 26(17), 5161. https://doi.org/10.3390/molecules26175161