
Cholecystokinin and Panic Disorder: Reflections on the History and Some Unsolved Questions

{kind=link}
Abstract
:1. Introduction
2. The Bioactivity of CCK-4
3. The Panicogenic Activity of CCK-4
4. Does the Brain Synthesize CCK-4 as a Separate Neuropeptide?
5. Is Endogenous CCK in Plasma Associated with Panic Disorder?
6. CCK2-Receptor Antagonists in Panic Disorder Therapy
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ivy, A.C.; Oldberg, E. Hormone mechanism for gallbladder contraction and evacuation. Am. J. Physiol. 1928, 86, 599–613. [Google Scholar] [CrossRef]
- Rehfeld, J.F. Cholecystokinin. In Handbook of Physiology, the Gastrointestinal System: Neural and Endocrine Biology; Schultz, S.G., Makhlouf, G.M., Rauner, B.B., Eds.; The American Physiology Society: Bethesda, MD, USA, 1989; pp. 337–358. [Google Scholar]
- Rehfeld, J.F. Cholecystokinin—From local gut hormone to ubiquitous messenger. Front. Endocrinol. 2017, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Rehfeld, J.F. Cholecystokinin and the hormone concept. Endocr. Connect. 2021, 10, R139–R150. [Google Scholar] [CrossRef]
- Vanderhaeghen, J.J.; Signeau, J.C.; Gepts, W. New peptide in the vertebrate CNS reacting with antigastrin antibodies. Nature 1975, 257, 604–605. [Google Scholar] [CrossRef]
- Dockray, G.J. Immunochemical evidence of cholecystokinin-like peptides in brain. Nature 1976, 264, 568–570. [Google Scholar] [CrossRef]
- Müller, J.E.; Strauss, E.; Yalow, R.S. Cholecystokinin and its COOH-terminal octapeptide in the pig brain. Proc. Natl. Acad. Sci. USA 1977, 74, 3035–3037. [Google Scholar] [CrossRef] [Green Version]
- Rehfeld, J.F. Gastrins and cholecystokinins in brain and gut. Acta Pharmacol. Toxicol. 1977, 41, 24–25. [Google Scholar]
- Rehfeld, J.F. Immunochemical studies on cholecystokinin. II. Distribution and molecular heterogeneity in the central nervous system and the small intestine of man and hog. J. Biol. Chem. 1978, 253, 4022–4030. [Google Scholar] [CrossRef]
- Larsson, L.-I.; Rehfeld, J.F. Localization and molecular heterogeneity of cholecystokinin in the central and peripheral nervous system. Brain Res. 1979, 165, 201–218. [Google Scholar] [CrossRef]
- Crawley, J.N. Comparative distribution of cholecystokinin and other neuropeptides: Why is this peptide different from all other peptides? Ann. N. Y. Acad. Sci. 1985, 448, 1–8. [Google Scholar] [CrossRef]
- Dodd, J.; Kelly, J.S. Cholecystokinin peptides: Excitatory effect on hippocampal neurons. J. Physiol. 1979, 195, 61. [Google Scholar]
- Emson, P.C.; Lee, C.M.; Rehfeld, J.F. Cholecystokinin octapeptide: Vesicular localization and calcium dependent release from rat brain in vitro. Life Sci. 1980, 26, 2157–2163. [Google Scholar] [CrossRef]
- Dockray, G.J.; Gregory, R.A.; Hutchison, J.B.; Harris, J.I.; Runswick, M.J. Isolation, structure and biological activity of two cholecystokinin octapeptides from sheep brain. Nature 1978, 274, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Rehfeld, J.F.; Hansen, H.F. Characterization of preprocholecystokinin products in porcine cerebral cortex: Evidence of different processing pathways. J. Biol. Chem. 1986, 261, 5832–5840. [Google Scholar] [CrossRef]
- Agersnap, M.; Zhang, M.-D.; Harkany, T.; Hökfelt, T.; Rehfeld, J.F. Nonsulfated cholecystokinins in cerebral neurons. Neuropeptides 2016, 60, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Eberlein, G.A.; Eysselein, V.E.; Davis, M.T.; Lee, T.D.; Shively, J.E.; Grandt, D.; Niebel, W.; Williams, R.; Moessner, J.; Zeeh, J.; et al. Patterns of prohormone processing: Order revealed by a new procholecystokinin-derived peptide. J. Biol. Chem. 1992, 267, 1517–1521. [Google Scholar] [CrossRef]
- Rehfeld, J.F.; Sun, G.; Christensen, T.; Hillingsø, J.G. The predominant cholocystokinin in human plasm and intestine is cholecystokinin-33. J. Clin. Endocrinol. Metab. 2001, 86, 251–258. [Google Scholar]
- Agersnap, M.; Rehfeld, J.F. Nonsulfated cholecystokinins in the small intestine of pigs and rats. Peptides 2015, 71, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Rehfeld, J.F.; Bundgaard, J.R.; Hannibal, J.; Zhu, X.; Norrbom, C.; Steiner, D.F.; Friis-Hansen, L. The cell-specific pattern of cholecystokinin peptides in endocrine cells versus neurons is governed by the expression of prohormone convertases 1/3, 2, and 5/6. Endocrinology 2008, 149, 1600–1608. [Google Scholar] [CrossRef] [Green Version]
- Kessler, R.C.; McGonagle, K.A.; Zhao, S.; Nelson, C.B.; Hughes, M.; Eshleman, S.; Wittchen, H.U.; Kendler, K.S. Lifetime and 12-month prevalence of DSM-III-R psychiatric disorders in the United States. Results from the national Comorbidity Survey. Arch. Gen. Psychiatry 1994, 51, 8–19. [Google Scholar] [CrossRef]
- LeDoux, J.E. Emotion circuits in the brain. Annu. Rev. Neurosci. 2000, 23, 155–184. [Google Scholar] [CrossRef]
- Davis, M.; Whalen, P.J. The amygdala: Vigilance and emotion. Mol. Psychiatry 2001, 6, 13–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez de la Mora, M.; Hernández-Gómez, A.M.; Arizmendi-Garcia, Y.; Jacobsen, K.X.; Lara-Garcia, D.; Flores-Gracia, C.; Crespo-Ramirez, M.; Gallegos-Cari, A.; Nuche-Bricaire, A.; Fuxe, K. Role of the amygdoloid cholecystokinin (CCK)/gastrin-2 receptors and terminal networks in the modulation of anxiety in the rat. Effects of CCK-4 and CCK-8S on anxiety-like behaviour and [3H] GABA release. Eur. J. Neurosci. 2007, 26, 3614–3630. [Google Scholar] [CrossRef]
- Vanderhaeghen, J.J.; Lotstra, F.; De Mey, J.; Gilles, C. Immunohistochemical localization of cholecystokinin- and gastrin-like peptides in the brain and hypophysis of the rat. Proc. Natl. Acad. Sci. USA 1980, 77, 1190–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mascagni, F.; McDonald, A.J. Immunohistochemical characterization of cholecystokinin containing neurons in the rat basolateral amygdala. Brain Res. 2003, 976, 171–184. [Google Scholar] [CrossRef]
- Zarbin, M.A.; Innis, R.B.; Wamsley, J.K.; Snyder, S.H.; Kuhar, M.J. Autoradiographic localization of cholecystokinin receptors in rodent brain. J. Neurosci. 1983, 3, 877–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, T.; Wada, E.; Battey, J.F.; Wank, S.A. Differential expression of CCK(A) and CCK(B) receptors in the rat brain. Mol. Cell. Neurosci. 1993, 4, 143–154. [Google Scholar] [CrossRef]
- Mercer, L.D.; Le, V.Q.; Nunan, J.; Jones, N.M.; Beart, P.M. Direct visualization of cholecystokinin subtype2 receptors in rat central nervous system using antipeptide antibodies. Neurosci. Lett. 2000, 293, 167–170. [Google Scholar] [CrossRef]
- Rex, A.; Barth, T.; Voigt, J.P.; Domeney, A.M.; Fink, H. Effects of cholecystokinin tetrapeptide and sulfated cholecystokinin octapeptide in rat models of anxiety. Neurosci. Lett. 1994, 172, 139–142. [Google Scholar] [CrossRef]
- Köks, S.; Männistö, P.T.; Bourin, M.; Shlik, J.; Vasar, V.; Vasar, E. Cholecystokinin-induced anxiety in rats: Relevance of pre-experimental stress and seasonal variations. J. Psychiatry Neurosci. 2000, 25, 33–42. [Google Scholar] [PubMed]
- Gregory, R.A.; Tracy, H.J. The constitution and properties of two gastrins extracted from hog antral mucosa. Gut 1964, 5, 103–117. [Google Scholar] [CrossRef] [Green Version]
- Gregory, H.; Hardy, P.M.; Jones, D.S.; Kenner, G.W.; Sheppard, R.C. The antral hormone gastrin. I. Structure of gastrin. Nature 1964, 204, 931–933. [Google Scholar] [CrossRef]
- Morley, J.S.; Tracy, H.J.; Gregory, R.A. Structure-function relationships in the active C-terminal tetrapeptide sequence of gastrin. Nature 1965, 207, 1356–1359. [Google Scholar] [CrossRef] [PubMed]
- Mutt, V.; Jorpes, J.E. Structure of porcine cholecystokinin—Pancreozymin. I. Cleavage with thrombin and with trypsin. Eur. J. Biochem. 1968, 6, 156–162. [Google Scholar] [CrossRef]
- Mutt, V.; Jorpes, J.E. Hormonal polypeptides of the upper intestine. Biochem. J. 1971, 125, 57P–58P. [Google Scholar] [CrossRef] [Green Version]
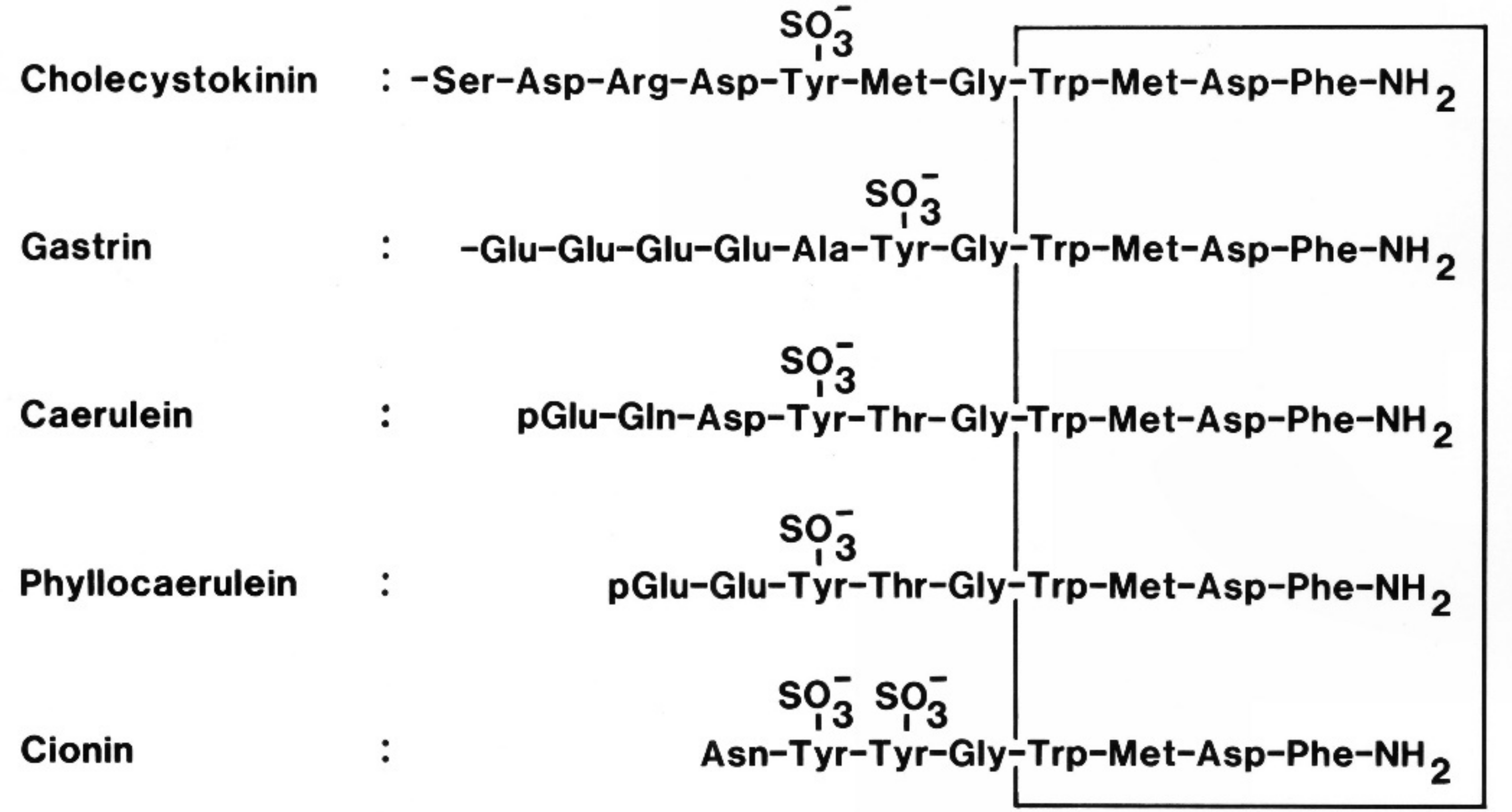
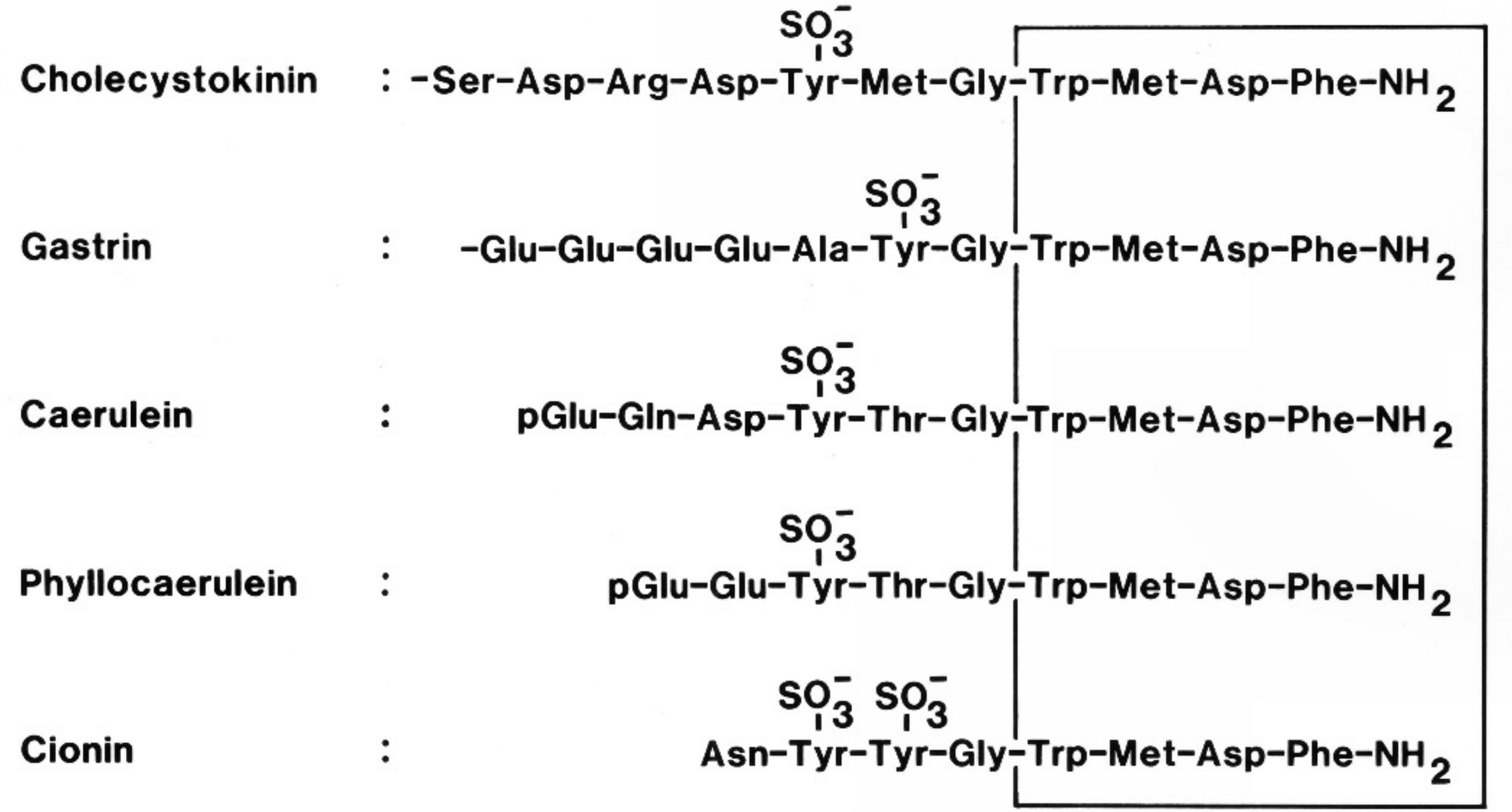
- Larsson, L.-I.; Rehfeld, J.F. Evidence for a common evolutionary origin of gastrin and cholecystokinin. Nature 1977, 269, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, A.H. Phylogeny of the cholecystokinin/gastrin family. Front. Neuroendocrinol. 1998, 19, 73–99. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, B.S.; Patel, O.; Shulkes, A. Evolution of gastrointestinal hormones: The cholecystokinin/gastrin family. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 77–88. [Google Scholar] [CrossRef]
- Dupré, D.; Tostivint, H. Evolution of the gastrin-cholecystokinin gene family by synteny analysis. Gen. Comp. Endocrinol. 2014, 195, 164–173. [Google Scholar] [CrossRef]
- Elrick, H.; Stimmler, L.; Hlad, C.J.; Arai, Y. Plasma insulin responses to oral and intravenous glucose administration. J. Clin. Endocrinol. Metab. 1964, 24, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, N.; Holdsworth, C.D.; Turner, D.A. New interpretation of oral glucose tolerance. Lancet 1964, 2, 20–21. [Google Scholar] [CrossRef]
- Rehfeld, J.F. The origin and understanding of the incretin concept. Front. Endocrinol. 2018, 9, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehfeld, J.F. Effect of gastrin and its C-terminal tetrapeptide on insulin secretion in man. Acta Endocrinol. 1971, 66, 169–176. [Google Scholar] [CrossRef]
- Rehfeld, J.F.; Stadil, F. The effect of gastrin on basal- and glucose-stimulated insulin secretion in man. J. Clin. Investig. 1973, 52, 1415–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehfeld, J.F.; Larsson, L.-I.; Goltermann, N.R.; Schwartz, T.W.; Holst, J.J.; Jensen, S.L.; Morley, J.S. Neural regulation of pancreatic hormone secretion by the C-terminal tetrapeptide of CCK. Nature 1980, 284, 33–38. [Google Scholar] [CrossRef]
- Rehfeld, J.F. CCK and anxiety: An introduction. In Multiple Cholecystokinin Receptors in CNS; Dourish, C.T., Cooper, S.J., Iversen, S.D., Iversen, L.L., Eds.; Oxford University Press: Oxford, UK, 1992; pp. 117–120. [Google Scholar]
- Vanderhaeghen, J.J.; Crawley, J.N. Neuronal cholecystokinin. In International Conference on Neuronal Cholecystokinin 1984: Brussels, Belgium; New York Academy of Sciences: New York, NY, USA, 1985; Volume 448, pp. 1–686. [Google Scholar]
- Bradwejn, J.; de Montigny, C. Benzodiazepine antagonizes cholecystokinin-induced activation of rat hippocampal neurons. Nature 1984, 321, 363–364. [Google Scholar] [CrossRef]
- Bradwejn, J.; de Montigny, C. Antagonism of cholecystokinin-induced activation by benzodiazepine receptor antagonists. Ann. N. Y. Acad. Sci. 1985, 448, 575–580. [Google Scholar] [CrossRef]
- de Montigny, C. Cholecystokinin tetrapeptide induces panic-like attacks in healthy volunteers: Preliminary findings. Arch. Gen. Psychiatry 1989, 46, 511–517. [Google Scholar] [CrossRef]
- Bradwejn, J.; Koszycki, D.; Meterissian, G. Cholecystokinin tetrapeptide induces panic attacks in patients with panic disorders. Can. J. Psychiatry 1990, 35, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Bradwejn, J.; Koszycki, D.; Shriqui, C. Enhanced sensitivity to cholecystokinin tetrapeptide in panic disorder. Arch. Gen. Psychiatry 1991, 48, 603–607. [Google Scholar] [CrossRef]
- Bradwejn, J.; Koszycki, D.; Bourin, M. Dose-ranging study of the effect of CCK-4 in healthy volunteers. J. Psychiatr. Neurosci. 1991, 16, 260–264. [Google Scholar]
- Bradwejn, J.; Koszycki, D.; Couëtoux du Tertre, A.; van Megen, H.; den Boer, J.; Westenberg, H. The panicogenic effects of cholecystokinin tetrapeptide are antagonized by L-365,260, a central cholecystokinin receptor antagonist, in patients with panic disorder. Arch. Gen. Psychiatry 1994, 51, 486–493. [Google Scholar] [CrossRef]
- Bradwejn, J.; Vasar, E. (Eds.) Cholecystokinin and Anxiety: From Neuron to Behavior; Springer: Berlin/Heidelberg, Germany; R.G. Landes Co.: Austin, TX, USA, 1995; pp. 1–181. [Google Scholar]
- Bradwejn, J.; Koszycki, D. Cholecystokinin and panic disorder: Past and future clinical research strategies. Scand. J. Clin. Lab. Investig. 2001, 61 (Suppl. 234), 19–27. [Google Scholar] [CrossRef]
- Wang, H.; Wong, P.T.H.; Spiess, J.; Zhu, Y.Z. Cholecystokinin-2 (CCK2) receptor mediated anxiety-like behaviors in rats. Neurosci. Biobehav. Rev. 2005, 29, 1361–1373. [Google Scholar] [CrossRef]
- Zwanzger, P.; Domschke, K.; Bradwejn, J. Neuronal network of panic disorder: The role of the neuropeptide cholecystokinin. Depress. Anxiety 2012, 29, 762–774. [Google Scholar] [CrossRef] [PubMed]
- McCann, U.D.; Slate, S.O.; Geraci, M.; Roscow-Terrill, D.; Uhde, T.W. A comparison of the effects of intravenous pentagastrin on patients with social phobia, panic disorder and healthy controls. Neuropsychopharmacology 1997, 16, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Koszycki, D.; Zacharko, R.; Le Mellédo, J.M.; Bradwejn, J. Behavioural, cardiovascular and neuroendocrine profiles following CCK-4 challenge in healthy volunteers: A comparison of panickers and non-panickers. Depress. Anxiety 1998, 8, 1–7. [Google Scholar] [CrossRef]
- Shlik, J.; Aluoja, A.; Vasar, V.; Vasar, E.; Podar, T.; Bradwejn, J. Effects of citapram treatment on behavioural, cardiovascular, and neuroendocrine response to cholecystokinin tetrapeptide challenge in panic disorder patients. J. Psychiatry Neurosci. 1997, 22, 332–340. [Google Scholar]
- van Megen, H.J.; Westenberg, G.M.; den Boer, J.A.; Slaap, B.; Scheepmakers, A. Effect of the selective serotonin reuptake inhibitor fluvoxamine on CCK-4 induced panic attacks. Psychoparmacology 1997, 129, 357–364. [Google Scholar] [CrossRef]
- Zacharko, R.M.; Koszycki, D.; Mendella, P.D.; Bradwejn, J. Behavioral, neurochemical, anatomical and electrophysiological correlates of panic disorder: Multiple transmitter interaction and neuropeptide colocalizations. Prog. Neurobiol. 1995, 47, 371–423. [Google Scholar] [CrossRef]
- Crawley, J.N. Interactions between cholecystokinin and other neurotransmitter systems. In Cholecystokinin and Anxiety: From Neuron to Behavior; Bradwejn, J., Vasar, E., Eds.; Springer: Berlin/Heidelberg, Germany; R.G. Landes Co.: Austin, TX, USA, 1995; pp. 33–56. [Google Scholar]
- Koszycki, D.; Zacharko, R.; Le Melledo, J.M.; Young, S.N.; Bradwejn, J. Effect of acute tryptophan depletion on behavioural, cardiovascular and hormonal sensitivity to cholecystokinin-tetrepeptide challenge in healthy volunteers. Biol. Psychiatry 1996, 40, 648–655. [Google Scholar] [CrossRef]
- Le Mellédo, J.M.; Bradwejn, J.; Koszycki, D.; Bichet, D.G.; Ballavance, F. The role of the beta-noradrenergic system in cholecystokinin-tetrapeptide-induced panic symptoms. Biol. Psychiatry 1998, 44, 364–366. [Google Scholar] [CrossRef]
- Benkelfat, C.; Bradwejn, J.; Meyer, E.; Ellenbogen, M.; Milot, S.; Gjedde, A.; Evans, A. Functional neuroanatomy of CCK4-induced anxiety in normal healthy volunteers. Am. J. Psychiatry 1995, 152, 1180–1182. [Google Scholar]
- Javanmard, M.; Shilk, J.; Kennedy, S.H.; Vaccarino, F.J.; Houle, S.; Bradwejn, J. Neuroanatomic correlates of CCK-4-induced panic attacks in healthy humans: A comparison of two time points. Biol. Psychiatry 1999, 45, 872–882. [Google Scholar] [CrossRef]
- Denavit-Saubié, M.; Hurlé, M.A.; Morin-Surun, M.P.; Foutz, A.S.; Champagnat, J. The effects of cholecystokinin-8 in the nucleus tractus solitarius. Ann. N. Y. Acad. Sci. 1985, 448, 375–384. [Google Scholar] [CrossRef] [PubMed]
- de Leeuw, A.S.; den Boer, J.A.; Slaap, B.R.; Westenberg, H.G. Pentagastrin has panic-inducing properties in obsessive compulsive disorder. Psychopharmacology 1996, 126, 229–237. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, I.M.; Westenberg, H.G.; Slaap, B.R.; den Boer, J.A.; Ho Pial, K.L. Anxiogenic effects of pentagastrin in patients with social phobia and healthy controls. Biol. Psychiatry 1997, 42, 76–78. [Google Scholar] [CrossRef]
- Brawman-Minzter, O.; Lydiard, R.B.; Bradwejn, J.; Villarreal, G.; Knapp, R.; Emmanuel, N.; Ware, M.R.; He, Q.; Ballenger, J.C. Effects of the cholecystokinin agonist pentagastrin in patients with generalized anxiety disorder. Am. J. Psychiatry 1997, 154, 700–702. [Google Scholar]
- Hansen, T.V.O.; Rehfeld, J.F.; Nielsen, F.C. Function of the C-36 to T polymorphism in the human cholecystokinin gene promoter. Mol. Psychiatry 2000, 5, 443–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Valdes, J.; Noyes, R.; Zoega, T.; Crowe, R.R. Possible association of a cholecystokinin promotor polymorphism (CCK-36CT) with panic disorder. Am. J. Med. Genet. 1998, 81, 228–234. [Google Scholar] [CrossRef]
- Garvey, M.J.; Crowe, R.R.; Wang, Z. An association of NAG levels and a mutation of the CCK gene in panic disorder patients. Psychiatry Res. 1998, 80, 149–153. [Google Scholar] [CrossRef]
- Koefoed, P.; Woldbye, D.P.; Hansen, T.V.O.; Hansen, E.S.; Knudsen, G.M.; Bolwig, T.G.; Rehfeld, J.F. Gene variations in the cholecystokinin system in patients with panic disorder. Psychiatr. Genet. 2010, 20, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Hösing, V.G.; Schirmacher, A.; Kuhlenbäumer, G.; Freitag, C.; Sand, P.; Schlesiger, C.; Jacob, C.; Fritze, J.; Franke, P.; Rietschel, M.; et al. Cholecystokinin- and cholecystokinin-B-receptor gene polymorphism in panic disorder. J. Neural Transmission. Suppl. 2004, 68, 147–156. [Google Scholar]
- Hattori, E.; Yamada, K.; Toyota, T.; Yoshitsugu, K.; Toru, M.; Shibuya, H.; Yoshikawa, T. Association studies of the CT repeat polymorphism in the 5’ upstream region of the cholecystokinin B receptor gene with panic disorder and schizophrenia in Japanese subjects. Am. J. Med. Genet. 2001, 105, 779–782. [Google Scholar] [CrossRef]
- Eysselein, V.E.; Reeve, J.R.; Shively, J.E.; Miller, C.; Walsh, J.H. Isolation of a large cholecystokinin precursor from canine brain. Proc. Natl. Acad. Sci. USA 1984, 81, 6565–6568. [Google Scholar] [CrossRef] [Green Version]
- Tatemoto, K.; Jörnvall, H.; Siimesmaa, S.; Halldén, G.; Mutt, V. Isolation and characterization of cholecystokinin-58 (CCK-58) from porcine brain. FEBS Lett. 1984, 174, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Shively, J.; Reeve, J.R.; Eysselein, V.E.; Ben-Avram, C.; Vigna, S.R.; Walsh, J.H. CCK-5: Sequence analysis of a small cholecystokinin from canine brain and intestine. Am. J. Physiol. 1987, 252, G272–G275. [Google Scholar] [CrossRef]
- Süsens, U.; Hermans-Borgmeyer, I.; Urny, J.; Schaller, H.C. Characterisation and differential expression of two very closely related G-protein-coupled receptors, GPR139 and GPR142, in mouse tissue and during mouse development. Neuropharmacology 2006, 50, 512–520. [Google Scholar] [CrossRef]
- Regard, J.B.; Sato, I.T.; Coughlin, S.R. Anatomical profiling of G protein-coupled receptor expression. Cell 2008, 135, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, O.; Shang, J.; Munk, A.; Ekberg, J.P.; Petersen, N.; Engelstoft, M.S.; Egerod, K.L.; Hjorth, S.A.; Wu, M.; Feng, Y.; et al. The aromatic amino acid sensor GPR142 controls metabolism through balanced regulation of pancreatic and gut hormones. Mol. Metab. 2019, 19, 49–64. [Google Scholar] [CrossRef]
- Reubi, J.C.; Waser, B.; Gugger, M.; Friess, H.; Kleeff, J.; Kayed, H.; Büchler, M.W.; Laissue, J.A. Distribution of CCK1 and CCK2 receptors in normal and diseased human pancreatic tissue. Gastroenterology 2003, 125, 98–106. [Google Scholar] [CrossRef]
- Rehfeld, J.F. Accumulation of nonamidated preprogastrin and preprocholecystokinin products in porcine pituitary corticotrophs: Evidence of post-translational control of cell differentiation. J. Biol. Chem. 1986, 261, 5841–5847. [Google Scholar] [CrossRef]
- Rehfeld, J.F. Preprocholecystokinin processing in the normal human anterior pituitary. Proc. Natl. Acad. Sci. USA 1987, 84, 3019–3023. [Google Scholar] [CrossRef] [Green Version]
- Rehfeld, J.F.; Johnsen, A.H.; Ødum, L.; Bardram, L.; Schifter, S.; Scopsi, L. Non-sulphated cholecystokinin in human medullary thyroid carcinomas. J. Endocrinol. 1990, 124, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Bardram, L.; Hilsted, L.; Rehfeld, J.F. Cholecystokinin, gastrin and their precursors in pheochromocytomas. Acta Endocrinol. 1989, 120, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Persson, H.; Rehfeld, J.F.; Ericsson, A.; Schalling, M.; Pelto-Huikko, M.; Hökfelt, T. Transient expression of the cholecystokinin gene in male germ cells and accumulation of the peptide in the acrosomal granule: Possible role of cholecystokinin in fertilization. Proc. Natl. Acad. Sci. USA 1989, 86, 6166–6170. [Google Scholar] [CrossRef] [Green Version]
- Rehfeld, J.F. Accurate measurement of cholecystokinin in plasma. Clin. Chem. 1998, 44, 991–1001. [Google Scholar] [CrossRef] [Green Version]
- Rehfeld, J.F.; Sennels, H.P.; Jørgensen, H.L.; Fahrenkrug, J. Circadian variations in plasma concentrations of cholecystokinin and gastrin in man. Scand. J. Clin. Lab. Investig. 2020, 80, 546–551. [Google Scholar] [CrossRef]
- Gibbs, J.; Young, R.C.; Smith, G.P. Cholecystokinin elicits satiety in rats with open gastric fistulas. Nature 1973, 245, 323–325. [Google Scholar] [CrossRef]
- Smith, G.P.; Jerome, C.; Cushin, B.J.; Eterno, R.; Simansky, K.J. Abdominal vagotomy blocks the satiety effect of cholecystokinin in the rat. Science 1981, 213, 1036–1037. [Google Scholar] [CrossRef]
- Kramer, M.S.; Cutler, N.R.; Ballenger, J.C.; Patterson, W.M.; Mendels, J.; Chenault, A.; Shrivastava, R.; Matzura-Wolfe, D.; Lines, C.; Reines, S. A placebo-controlled trial of L-365,260, a CCKB antagonist, in panic disorder. Biol. Psychiatry 1995, 37, 462–466. [Google Scholar] [CrossRef]
- Bradwejn, J.; Koszycki, D.; Paradis, M.; Reece, P.; Hinton, J.; Sedman, A. Effect of CI-988 on cholecystokinin tetrapeptide-induced panic symptoms in healthy volunteers. Biol. Psychiatry 1995, 38, 742–746. [Google Scholar] [CrossRef]
- van Megen, H.J.; Westenberg, G.M.; den Boer, J.A.; Slaap, B.; van Es-Radhakishun, F.; Pande, A.C. The cholecystokinin-B receptor antagonist CI-988 failed to affect CCK-4 induced symptoms in panic disorder patients. Psychopharmacology 1997, 129, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Ballaz, S.J.; Bourin, M.; Akil, H.; Watson, S.J. Blockade of the cholecystokinin CCK-2 receptor prevents the normalization of anxiety levels in the rat. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 96, 109761. [Google Scholar] [CrossRef]
- Innis, R.B.; Snyder, S.H. Distinct cholecystokinin receptors in brain and pancreas. Proc. Natl. Acad. Sci. USA 1980, 77, 6917–6921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.M.; Beinborn, M.; McBride, E.W.; Lu, M.; Kolakowski, L.F.; Kopin, A.S. The human brain cholecystokinin-B/gastrin receptor. Cloning and characterization. J. Biol. Chem. 1993, 268, 8164–8169. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rehfeld, J.F. Cholecystokinin and Panic Disorder: Reflections on the History and Some Unsolved Questions. Molecules 2021, 26, 5657. https://doi.org/10.3390/molecules26185657
Rehfeld JF. Cholecystokinin and Panic Disorder: Reflections on the History and Some Unsolved Questions. Molecules. 2021; 26(18):5657. https://doi.org/10.3390/molecules26185657
Chicago/Turabian StyleRehfeld, Jens F. 2021. "Cholecystokinin and Panic Disorder: Reflections on the History and Some Unsolved Questions" Molecules 26, no. 18: 5657. https://doi.org/10.3390/molecules26185657
APA StyleRehfeld, J. F. (2021). Cholecystokinin and Panic Disorder: Reflections on the History and Some Unsolved Questions. Molecules, 26(18), 5657. https://doi.org/10.3390/molecules26185657