
Anti-Fungal Hevein-like Peptides Biosynthesized from Quinoa Cleavable Hololectins

 ,
, 
 ,
,
Abstract
:
1. Introduction
2. Results and Discussions
2.1. Mass Spectrometry Screening of Quinoa Seed Extracts
2.2. Primary Sequence Determination of Chenotides
2.3. Solution Structure of Chenotide cQ2
2.4. Peptide Stability of Cystine-knotted Chenotide cQ2
2.5. Sequence and Structural Comparison of Chenotides with Other Chitin-Binding Hevein-Like Peptides
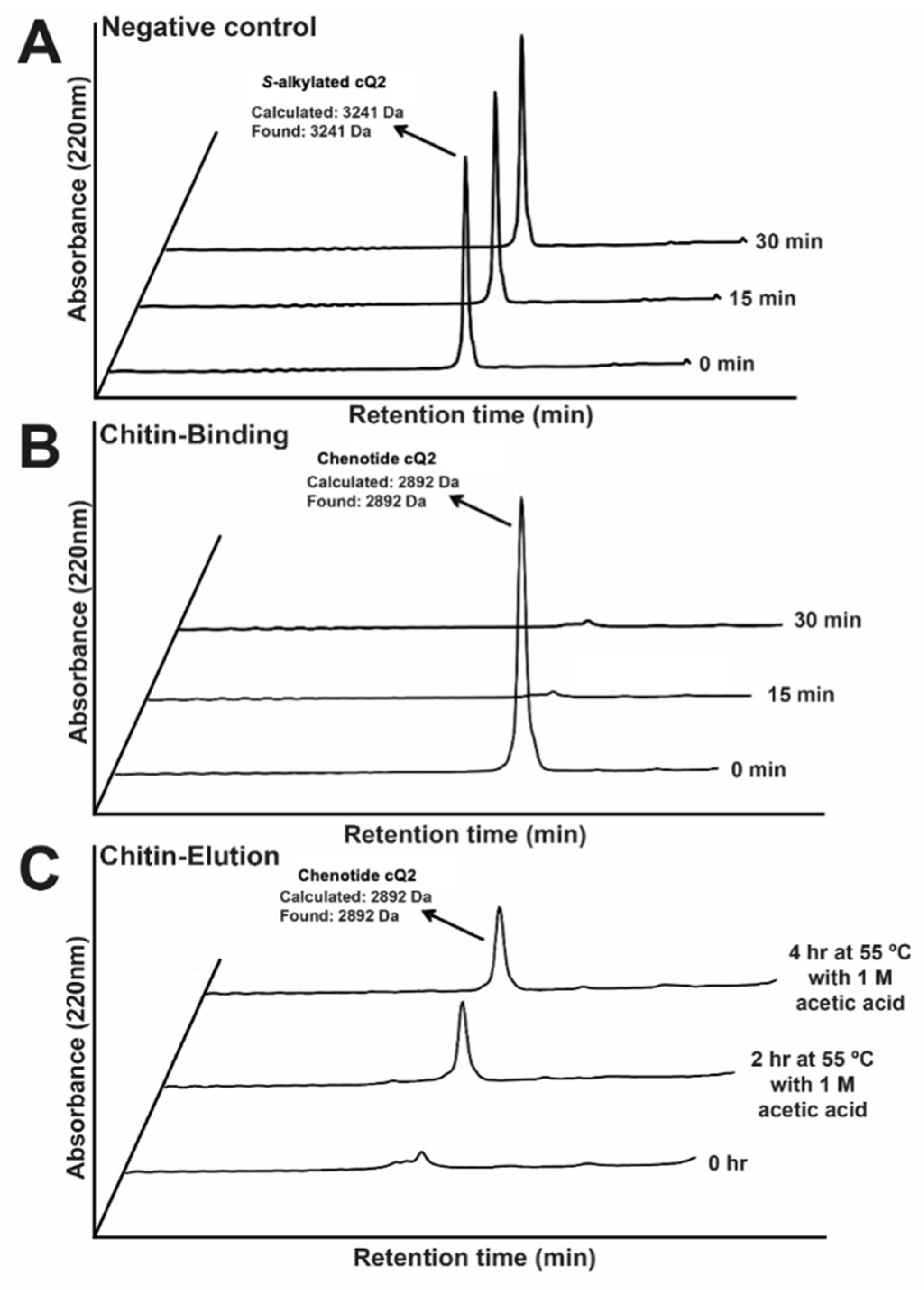
2.6. Chenotide cQ2 Is Chitin-Binding
2.7. Chenotide cQ2 Is an Anti-Fungal Peptide
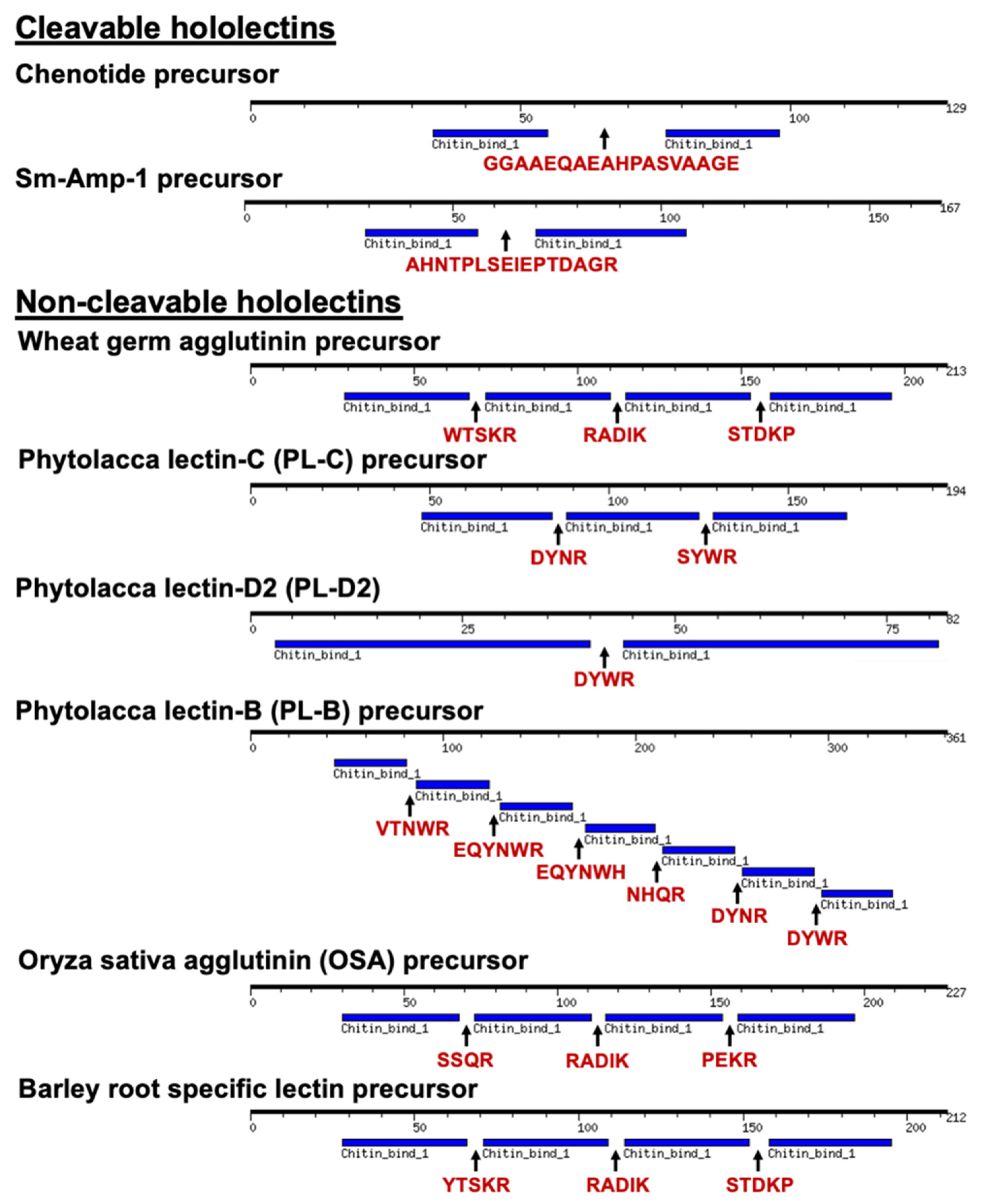
2.8. Chenotide Precursors Belong to a New Family of Cleavable Hololectins
3. Materials and Methods
3.1. Materials
3.2. Plant Materials
3.3. Extraction, Isolation, and Purification
3.4. Matrix-assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry
3.5. Sequence Determination
3.6. NMR Structural Study
3.7. Ligand Peptide Docking
3.8. Chitin-binding Assay
3.9. Peptide Stability Assays
3.10. Anti-Fungal Assay
3.11. Bioinformatics and Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bazile, D.; Jacobsen, S.-E.; Verniau, A. The global expansion of quinoa: Trends and limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [Green Version]
- Graf, B.L.; Rojas-Silva, P.; Rojo, L.; Delatorre-Herrera, J.; Baldeón, M.E.; Raskin, I. Innovations in health value and functional food development of quinoa (Chenopodium quinoa Willd.). Compr. Rev. Food Sci. Food Saf. 2015, 14, 431–445. [Google Scholar] [CrossRef] [Green Version]
- Walling, L.L. Chapter 13 Adaptive defense responses to pathogens and insects. Adv. Bot. Res. 2009, 51, 551–612. [Google Scholar]
- Doughari, J. An overview of plant immunity. J. Plant Pathol. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Delaux, P.-M.; Schornack, S. Plant evolution driven by interactions with symbiotic and pathogenic microbes. Science 2021, 371, eaba6605. [Google Scholar] [CrossRef] [PubMed]
- Egorov, T.A.; Odintsova, T.I. Defense peptides of plant immunity. Russ. J. Bioorganic Chem. 2012, 38, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kini, S.G.; Wong, K.H.; Tan, W.L.; Xiao, T.; Tam, J.P. Morintides: Cargo-free chitin-binding peptides from Moringa oleifera. BMC Plant Biol. 2017, 17, 68. [Google Scholar] [CrossRef] [Green Version]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial peptides from plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Montesinos, E. Antimicrobial peptides and plant disease control. FEMS Microbiol. Lett. 2007, 270, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lenardon, M.D.; Munro, C.; Gow, N.A. Chitin synthesis and fungal pathogenesis. Curr. Opin. Microbiol. 2010, 13, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Gidrol, X.; Chrestin, H.; Tan, H.; Kush, A. Hevein, a lectin-like protein from Hevea brasiliensis (rubber tree) is involved in the coagulation of latex. J. Biol. Chem. 1994, 269, 9278–9283. [Google Scholar] [CrossRef]
- Van Parijs, J.; Broekaert, W.F.; Goldstein, I.J.; Peumans, W.J. Hevein: An antifungal protein from rubber-tree (Hevea brasiliensis) latex. Planta 1991, 183, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Archer, B.L. The proteins of Hevea brasiliensis Latex. 4. Isolation and characterization of crystalline hevein. Biochem. J. 1960, 75, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Romero, A.; Ravichandran, K.; Soriano-García, M. Crystal structure of hevein at 2.8 Å resolution. FEBS Lett. 1991, 291, 307–309. [Google Scholar] [CrossRef] [Green Version]
- Andersen, N.H.; Cao, B.; Rodriguez-Romero, A.; Arreguin, B. Hevein: NMR assignment and assessment of solution-state folding for the agglutinin-toxin motif. Biochemistry 1993, 32, 1407–1422. [Google Scholar] [CrossRef]
- Loo, S.; Kam, A.; Xiao, T.; Tam, J.P. Bleogens: Cactus-Derived Anti-Candida Cysteine-Rich Peptides with Three Different Precursor Arrangements. Front. Plant Sci. 2017, 8, 2162. [Google Scholar] [CrossRef] [Green Version]
- Kam, A.; Loo, S.; Fan, J.-S.; Sze, S.K.; Yang, D.; Tam, J.P. Roseltide rT7 is a disulfide-rich, anionic, and cell-penetrating peptide that inhibits proteasomal degradation. J. Biol. Chem. 2019, 294, 19604–19615. [Google Scholar] [CrossRef]
- Loo, S.; Kam, A.; Li, B.B.; Feng, N.; Wang, X.; Tam, J.P. Discovery of Hyperstable Noncanonical Plant-Derived Epidermal Growth Factor Receptor Agonist and Analogs. J. Med. Chem. 2021, 64, 7746–7759. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Tan, W.L.; Serra, A.; Xiao, T.; Sze, S.K.; Yang, D.; Tam, J.P. Ginkgotides: Proline-Rich Hevein-Like Peptides from Gymnosperm Ginkgo biloba. Front. Plant Sci. 2016, 7, 1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, J.P.; Nguyen, G.K.T.; Loo, S.; Wang, S.; Yang, D.; Kam, A. Ginsentides: Cysteine and glycine-rich peptides from the ginseng family with unusual disulfide connectivity. Sci. Rep. 2018, 8, 16201. [Google Scholar] [CrossRef] [PubMed]
- Loo, S.; Kam, A.; Xiao, T.; Nguyen, G.K.T.; Liu, C.F.; Tam, J.P. Identification and Characterization of Roseltide, a Knottin-type Neutrophil Elastase Inhibitor Derived from Hibiscus sabdariffa. Sci. rep. 2016, 6, 39401. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.H.; Tan, W.L.; Kini, S.G.; Xiao, T.; Serra, A.; Sze, S.K.; Tam, J.P. Vaccatides: Antifungal Glutamine-Rich Hevein-Like Peptides from Vaccaria hispanica. Front. Plant Sci. 2017, 8, 1100. [Google Scholar] [CrossRef] [Green Version]
- Kini, S.G.; Nguyen, P.Q.T.; Weissbach, S.; Mallagaray, A.; Shin, J.; Yoon, H.S.; Tam, J.P. Studies on the Chitin Binding Property of Novel Cysteine-Rich Peptides from Alternanthera sessilis. Biochemistry 2015, 54, 6639–6649. [Google Scholar] [CrossRef]
- Li, S.-S.; Claeson, P. Cys/Gly-rich proteins with a putative single chitin-binding domain from oat (Avena sativa) seeds. Phytochemistry 2003, 63, 249–255. [Google Scholar] [CrossRef]
- Lee, O.S.; Lee, B.; Park, N.; Koo, J.C.; Kim, Y.H.; Karigar, C.; Chun, H.J.; Jeong, B.R.; Kim, D.H.; Nam, J. Pn-AMPs, the hevein-like proteins from Pharbitis nil confers disease resistance against phytopathogenic fungi in tomato, Lycopersicum esculentum. Phytochemistry 2003, 62, 1073–1079. [Google Scholar] [CrossRef]
- Koo, J.C.; Lee, S.Y.; Chun, H.J.; Cheong, Y.H.; Choi, J.S.; Kawabata, S.-i.; Miyagi, M.; Tsunasawa, S.; Ha, K.S.; Bae, D.W.; et al. Two hevein homologs isolated from the seed of Pharbitis nil L. exhibit potent antifungal activity. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1998, 1382, 80–90. [Google Scholar] [CrossRef]
- Martins, J.; Maes, D.; Loris, R.; Pepermans, H.A.; Wyns, L.; Willem, R.; Verheyden, P. 1H NMR Study of the Solution Structure of Ac-AMP2, a Sugar Binding Antimicrobial Protein Isolated fromAmaranthus caudatus. J. Mol. Biol. 1996, 258, 322–333. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Marien, W.; Terras, F.R.G.; De Bolle, M.F.G.; Proost, P.; Van Damme, J.; Dillen, L.; Claeys, M.; Rees, S.B. Antimicrobial peptides from Amaranthus caudatus seeds with sequence homology to the cysteine/glycine-rich domain of chitin-binding proteins. Biochemistry 1992, 31, 4308–4314. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.-H.; Xiang, Y.; Liu, X.-Z.; Zhang, Y.; Hu, Z.; Wang, D.-C. Two novel antifungal peptides distinct with a five-disulfide motif from the bark of Eucommia ulmoides Oliv. FEBS Lett. 2002, 521, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.-H.; Xiang, Y.; Tu, G.-Z.; Zhang, Y.; Wang, D.-C. Solution structure of Eucommia antifungal peptide: A novel structural model distinct with a five-disulfide motif. Biochemistry 2004, 43, 6005–6012. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Huang, R.-H.; Liu, X.-Z.; Zhang, Y.; Wang, D.-C. Crystal structure of a novel antifungal protein distinct with five disulfide bridges from Eucommia ulmoides Oliver at an atomic resolution. J. Struct. Biol. 2004, 148, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, K.A.; Moskal Jr, W.A.; Wu, H.C.; Underwood, B.A.; Graham, M.A.; Town, C.D.; VandenBosch, K.A. Small cysteine-rich peptides resembling antimicrobial peptides have been under-predicted in plants. Plant J. 2007, 51, 262–280. [Google Scholar] [CrossRef] [PubMed]
- Slavokhotova, A.A.; Shelenkov, A.; Andreev, Y.A.; Odintsova, T.I. Hevein-like antimicrobial peptides of plants. Biochem. 2017, 82, 1659–1674. [Google Scholar] [CrossRef] [PubMed]
- Porto, W.F.; Souza, V.A.; Nolasco, D.O.; Franco, O.L. In silico identification of novel hevein-like peptide precursors. Peptides 2012, 38, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Kam, A.; Loo, S.; Dutta, B.; Sze, S.K.; Tam, J.P. Plant-derived mitochondria-targeting cysteine-rich peptide modulates cellular bioenergetics. J. Biol. Chem. 2019. [Google Scholar] [CrossRef] [Green Version]
- Verheyden, P.; Jurgen, P.; Dominique, M.; Henri, A.M.P.; Lode, W.; Rudolph, W.; JoséC, M. 1H NMR study of the interaction of N, N′, N ″-triacetyl chitotriose with Ac-AMP2, a sugar binding antimicrobial protein isolated from Amaranthus caudatus. FEBS Lett. 1995, 370, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Andreev, Y.A.; Korostyleva, T.V.; Slavokhotova, A.A.; Rogozhin, E.A.; Utkina, L.L.; Vassilevski, A.A.; Grishin, E.V.; Egorov, T.A.; Odintsova, T.I. Genes encoding hevein-like defense peptides in wheat: Distribution, evolution, and role in stress response. Biochimie 2012, 94, 1009–1016. [Google Scholar] [CrossRef]
- Odintsova, T.I.; Vassilevski, A.; Slavokhotova, A.A.; Musolyamov, A.K.; Finkina, E.I.; Khadeeva, N.V.; Rogozhin, E.A.; Korostyleva, T.V.; Pukhalsky, V.A.; Grishin, E.V.; et al. A novel antifungal hevein-type peptide from Triticum kiharae seeds with a unique 10-cysteine motif. FEBS J. 2009, 276, 4266–4275. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Shelenkov, A.A.; Korostyleva, T.V.; Rogozhin, E.A.; Melnikova, N.V.; Kudryavtseva, A.V.; Odintsova, T.I. Defense peptide repertoire of Stellaria media predicted by high throughput next generation sequencing. Biochimie 2017, 135, 15–27. [Google Scholar] [CrossRef]
- Lipkin, A.; Anisimova, V.; Nikonorova, A.; Babakov, A.; Krause, E.; Bienert, M.; Grishin, E.; Egorov, T. An antimicrobial peptide Ar-AMP from amaranth (Amaranthus retroflexus L.) seeds. Phytochemistry 2005, 66, 2426–2431. [Google Scholar] [CrossRef]
- Bergh, K.P.V.D.; Van Damme, E.J.; Peumans, W.J.; Coosemans, J. Ee-CBP, a hevein-type antimicrobial peptide from bark of the spindle tree (Euonymus europaeus L.). Mededelingen 2002, 67, 327–331. [Google Scholar]
- Bergh, K.P.V.D.; Proost, P.; Van Damme, J.; Coosemans, J.; Van Damme, E.J.; Peumans, W.J. Five disulfide bridges stabilize a hevein-type antimicrobial peptide from the bark of spindle tree (Euonymus europaeus L.). FEBS Lett. 2002, 530, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Craik, D.J.; Malik, U. Cyclotide biosynthesis. Curr. Opin. Chem. Biol. 2013, 17, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.-H. Evolution of Gene Duplication in Plants. Plant physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [Green Version]
- Peumans, W.J.; Van Damme, E. Lectins as plant defense proteins. Plant physiol. 1995, 109, 347. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.; Lannoo, N.; Peumans, W.J. Plant lectins. Adv. Bot. Res. 2008, 48, 107–209. [Google Scholar]
- Mishra, A.; Behura, A.; Mawatwal, S.; Kumar, A.; Naik, L.; Mohanty, S.S.; Manna, D.; Dokania, P.; Mishra, A.; Patra, S.K.; et al. Structure-function and application of plant lectins in disease biology and immunity. Food Chem. Toxicol. 2019, 134, 110827. [Google Scholar] [CrossRef]
- Smith, J.J.; Raikhel, N.V. Nucleotide sequences of cDNA clones encoding wheat germ agglutinin isolectins A and D. Plant Mol. Biol. 1989, 13, 601–603. [Google Scholar] [CrossRef]
- Wright, H.T.; Brooks, D.M.; Wright, C.S. Evolution of the multidomain protein wheat germ agglutinin. J. Mol. Evol. 1985, 21, 133–138. [Google Scholar] [CrossRef]
- Zhang, W.; Peumans, W.J.; Barre, A.; Astoul, C.H.; Rovira, P.; Rougé, P.; Proost, P.; Truffa-Bachi, P.; Jalali, A.A.; Van Damme, E.J. Isolation and characterization of a jacalin-related mannose-binding lectin from salt-stressed rice (Oryza sativa) plants. Planta 2000, 210, 970–978. [Google Scholar]
- Yamaguchi, K.-i.; Mori, A.; Funatsu, G. The complete amino acid sequence of lectin-C from the roots of pokeweed (Phytolacca americana). Biosci. Biotechnol. Biochem. 1995, 59, 1384–1385. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.-i.; Mori, A.; Funatsu, G. Amino acid sequence and some properties of lectin-D from the roots of pokeweed (Phytolacca americana). Biosci. Biotechnol. Biochem. 1996, 60, 1380–1382. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.-i.; Yurino, N.; Kino, M.; Ishiguro, M.; Funatsu, G. The amino acid sequence of mitogenic lectin-B from the roots of pokeweed (Phytolacca americana). Biosci. Biotechnol. Biochem. 1997, 61, 690–698. [Google Scholar] [CrossRef] [Green Version]
- Lerner, D.R.; Raikhel, N.V. Cloning and characterization of root-specific barley lectin. Plant physiol. 1989, 91, 124–129. [Google Scholar] [CrossRef] [Green Version]
- Delaglio, F.; Grzesiek, S.; Vuister, G.W.; Zhu, G.; Pfeifer, J.; Bax, A. NMRPipe: A multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 1995, 6, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced software for biomolecular NMR spectroscopy. Bioinformatics 2014, 31, 1325–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brünger, A.T.; Adams, P.D.; Clore, G.M.; DeLano, W.L.; Gros, P.; Grosse-Kunstleve, R.W.; Jiang, J.-S.; Kuszewski, J.; Nilges, M.; Pannu, N.S.; et al. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D. 1998, 54, 905–921. [Google Scholar]
- Laskowski, R.A.; Rullmann, J.A.C.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef]
- Huang, C.C.; Couch, G.S.; Pettersen, E.F.; Ferrin, T.E. Chimera: An extensible molecular modeling application constructed using standard components. In Proceedings of the Pacific Symposium on Biocomputing, Hawaii, HI, USA, 3–6 January 1996; p. 724. [Google Scholar]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Species | Amino Acid Sequence | Mass (Da) 1 | Charge 2 | pI | Similarity (%) |
|---|---|---|---|---|---|---|
| cQ1 | C. quinoa | AGECVRGRCPGGLCCSKFGFCGSGPAYCGGA | 2963 | +2 | 8.35 | 100 |
| cQ2 | C. quinoa | AGECVRGRCPGGLCCSKFGFCGSGPAYCGG | 2892 | +2 | 8.35 | 100 |
| cQ3 | C. quinoa | AGECVRGRCPGGLCCSKFGFCGSGPAYCG | 2835 | +2 | 8.35 | 100 |
| Ac-AMP1 | A. caudatus | VGECVRGRCPSGMCCSQFGYCGKGPKYCG | 3025 | +3 | 8.66 | 89.3 |
| Ac-AMP2 | A. caudatus | VGECVRGRCPSGMCCSQFGYCGKGPKYCGR | 3181 | +4 | 8.92 | 89.3 |
| Ar-AMP | A. retroflexus L. | AGECVQGRCPSGMCCSQFGYCGRGPKYCGR | 3153 | +3 | 8.68 | 89.7 |
| IWF4 | B.vulgaris L. | SGECNMYGRCPPGYCCSKFGYCGVGRAYCG | 3181 | +2 | 8.27 | 80.0 |
| aSG1 | A. sessilis | APGQCNHGRCPSGLCCSQYGYCGTGPAYCG | 3004 | +1 | 7.82 | 89.3 |
| aSG2 | A. sessilis | AGECNHGRCPSGLCCSQYGYCGTGPRYCG | 2992 | +1 | 7.82 | 86.2 |
| aSG3 | A. sessilis | APGQCNHGRCPSGICCSQYGYCGTGPAYCGG | 3060 | +1 | 7.82 | 89.7 |
| aSR1 | A. sessilis | VGECVQGRCPPGLCCSRFGYCGTGPAYCG | 2932 | +1 | 7.75 | 96.4 |
| aSR2 | A. sessilis | APGECKHGRCPPGICCSQYGYCGTGPAYCG | 3029 | +1 | 7.81 | 89.3 |
| aSR3 | A. sessilis | APGECKHGRCPPGICCSQYGYCGTGPAYC | 2973 | +1 | 7.81 | 88.9 |
| SmAMP1.1a | S. media | VGPGGECGRFGGCAGGQCCSRFGFCGSGPKYCAH | 3305 | +2 | 8.34 | 78.6 |
| SmaAMP3 | S. media | SGPNGQCGPGWGGCRGGLCCSQYGYCGSGPKYCAH | 3460 | +2 | 8.29 | 75.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loo, S.; Tay, S.V.; Kam, A.; Tang, F.; Fan, J.-S.; Yang, D.; Tam, J.P. Anti-Fungal Hevein-like Peptides Biosynthesized from Quinoa Cleavable Hololectins. Molecules 2021, 26, 5909. https://doi.org/10.3390/molecules26195909
Loo S, Tay SV, Kam A, Tang F, Fan J-S, Yang D, Tam JP. Anti-Fungal Hevein-like Peptides Biosynthesized from Quinoa Cleavable Hololectins. Molecules. 2021; 26(19):5909. https://doi.org/10.3390/molecules26195909
Chicago/Turabian StyleLoo, Shining, Stephanie V. Tay, Antony Kam, Fan Tang, Jing-Song Fan, Daiwen Yang, and James P. Tam. 2021. "Anti-Fungal Hevein-like Peptides Biosynthesized from Quinoa Cleavable Hololectins" Molecules 26, no. 19: 5909. https://doi.org/10.3390/molecules26195909
APA StyleLoo, S., Tay, S. V., Kam, A., Tang, F., Fan, J.-S., Yang, D., & Tam, J. P. (2021). Anti-Fungal Hevein-like Peptides Biosynthesized from Quinoa Cleavable Hololectins. Molecules, 26(19), 5909. https://doi.org/10.3390/molecules26195909