
Alpha-Glucosidase Inhibitory Assay-Screened Isolation and Molecular Docking Model from Bauhinia pulla Active Compounds




Abstract
:1. Introduction
2. Results and Discussion
2.1. Fractionation and Isolation Screening Based on Bioactivity
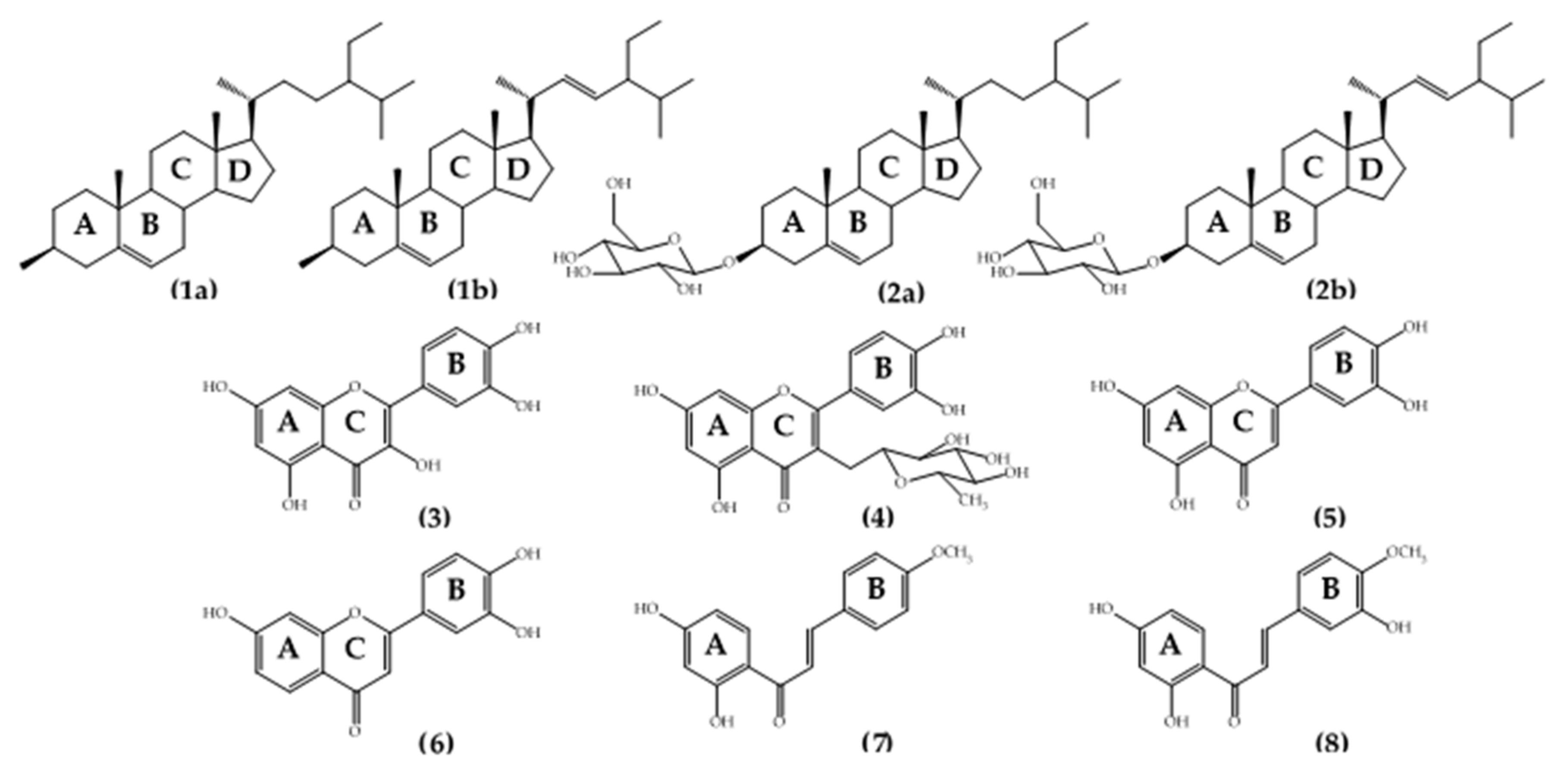
2.2. Identification of the Isolated Compounds
2.3. Alpha-Glucosidase Inhibitory Assay
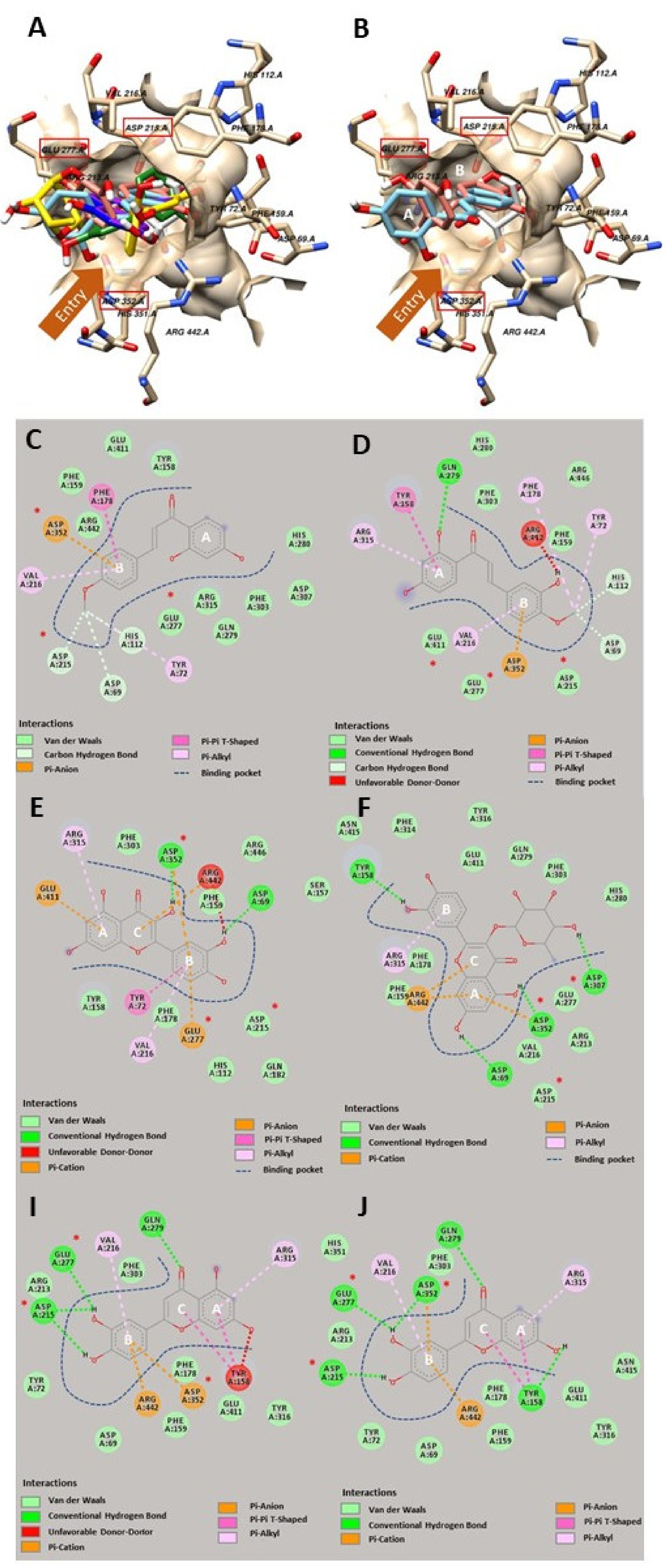
2.4. Computerized Molecular Docking
3. Discussion
4. Materials and Methods
4.1. General
4.2. Plant Extract Preparation
4.3. Alpha-Glucosidase Inhibitory Assay
4.4. Bioassay-Guidance Isolation
4.5. Computerized Molecular Docking
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Benalla, W.; Bellachen, S.; Bnouham, M. Antidiabetic medicinal plants as a source of alpha glucosidase inhibitors. Curr. Diabetes Rev. 2010, 6, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Houghton, P.J.; Soumyanath, A. α-Amylase inhibitory activity of some Malaysian plants used to treat diabetes with particular reference to Phyllanthus amarus. J. Ethnopharmacol. 2006, 107, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhang, G.; Pan, J.; Wang, Y. α-Glucosidase inhibition by luteolin: Kinetics, interaction and molecular docking. Int. J. Biol. Macromol. 2014, 64, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, A.; Singh, B.; Arora, R.; Saroj, A. In vitro evaluation of the α-glucosidase inhibitory potential of methanolic extracts of traditionally used in antidiabetic plants. BMC Complement Altern Med. 2019, 19, 74. [Google Scholar] [CrossRef] [Green Version]
- Wongon, M.; Limpeanchob, N. Inhibitory effect of Artocarpus lakoocha Roxb and oxyresveratrol on α-glucosidase and sugar digestion in Caco-2 cells. Heliyon 2020, 6, e03458. [Google Scholar] [CrossRef]
- Cechinel-Zanchett, C.C.; de Andrade, S.F.; Cechinel-Filho, V. Ethnopharmacological, phytochemical, pharmacological and toxicological aspects of Bauhinia forficata: A mini-review covering the last five years. Nat. Prod. Commun. 2018, 13, 911–916. [Google Scholar] [CrossRef] [Green Version]
- Gromek, K.; Drumond, N.; Paula, S. Pharmacovigilance of herbal medicines. Int. J. Risk Saf. Med. 2015, 25, 55–65. [Google Scholar] [CrossRef]
- Dej-adisai, S.; Pitakbut, T. Determination of α-glucosidase inhibitory activity from selected Fabaceae plants. Pak. J. of Pharm. Sci. 2015, 28, 1679–1683. [Google Scholar]
- Saisaard, P.; Chatan, W.; Maneechai, S. Diversity of the genus Bauhinia L. in the North-East of Thailand. SDU Res. J. Sci. Tech. 2015, 8, 87–115. [Google Scholar]
- Cechinel-Filho, V. Chemical composition and biological potential of plants from the genus Bauhinia. Phytother. Res. 2009, 23, 1347–1354. [Google Scholar] [CrossRef]
- Bunyaprapasorn, N.; Chokchaijarenpoon, O. Traditional Herbal Handbook; Faculty of Pharmacy, Mahidol University: Nakhon Pathom, Thailand, 2000; Volume 4–5, pp. 348, 700–701. [Google Scholar]
- Keo, S.; Lee, D.S.; Li, B.; Choi, H.G.; Kim, K.S.; Ko, W.; Oh, H.; Kim, Y.C. Neuroprotective effects of Cambodian plant extracts on glutamate-induced cytotoxicity in HT22 cells. Nat. Prod. Sci. 2012, 18, 177–182. [Google Scholar]
- De-Eknamkul, W.; Potduang, B. Biosynthesis of β-sitosterol and stigmasterol in Croton sublyratus proceeds via a mixed origin of isoprene units. Phytochem. 2003, 62, 389–398. [Google Scholar] [CrossRef]
- Khatun, M.; Billah, M.; Abdulquader, M. Sterols and sterol glucoside from Phyllanthus species. Dhaka. Univ. J. Sci. 2012, 60, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.W.; Chang, F.R.; Wu, Y.C. The contituents of Lindera glauca. J. Chin. Chem. Soc. 2000, 47, 373–380. [Google Scholar] [CrossRef]
- Hasan, M.S.; Ahmed, M.I.; Mondal, S.; Jamaluddin, S.; Masud, M.M.; Sadhu, S.K.; Ishibashi, M. Antioxidant, antinociceptive activity and general toxicity study of Dendrophthoe falcata and isolation of quercitrin as the major component. Orient. Pharm. Exp. Med. 2006, 6, 355–360. [Google Scholar]
- Zheng, Z.P.; Cheng, K.W.; Chao, J.; Wu, J.; Wang, M. Tyrosinase inhibitors from paper mulberry (Broussonetia papyrifera). Food Chem. 2008, 106, 529–535. [Google Scholar] [CrossRef]
- Júnior, G.M.V.; Sousa, C.M.M.; Cavalheiro, A.J.; Lago, J.H.; Chaves, M.H. Phenolic derivatives from fruits of Dipteryx lacunifera Ducke and evaluation of their antiradical activity. Helv. Chim. Acta. 2008, 91, 2159–2167. [Google Scholar] [CrossRef]
- Rao, Y.K.; Fang, S.H.; Tzeng, Y.M. Differential effects of synthesized 2’-oxygenated chalcone derivatives: Modulation of human cell cycle phase distribution. Bioorg. Med. Chem. 2004, 12, 2679–2686. [Google Scholar] [CrossRef]
- Rocha, S.; Sousa, A.; Ribeiro, D.; Correia, C.M.; Silva, V.L.; Santos, C.M.; Silva, A.M.; Araújo, A.N.; Fernandes, E.; Freitas, M. A study towards drug discovery for the management of type 2 diabetes mellitus through inhibition of the carbohydrate-hydrolyzing enzymes α-amylase and α-glucosidase by chalcone derivatives. Food Funct. 2019, 10, 5510–5520. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Liu, X.; Jiang, Z.; Geng, S.; Ma, H.; Liu, B. Interaction mechanism of flavonoids and α-glucosidase: Experimental and molecular modelling studies. Foods 2019, 8, 355. [Google Scholar] [CrossRef] [Green Version]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.T.; Sousa, J.L.C.; Tomé, S.M.; Ramos, M.J.; Silva, A.M.S.; Fernandes, P.A.; Fernandes, A. α-Glucosidase inhibition by flavonoids: An in vitro and in silico structure-activity relationship study. J. Enzyme Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Petitjean, M.; Teste, M.A.; Kooli, W.; Tranier, S.; François, J.M.; Parrou, J.L. Similarities and differences in the biochemical and enzymological properties of the four isomaltases from Saccharomyces cerevisae. FEBS Open Bio 2014, 4, 200–212. [Google Scholar] [CrossRef]
- Yamamoto, K.; Miyake, H.; Kusunoki, M.; Osaki, S. Crystal structures of isomaltase from Saccharomyces cerevisiae and in complex with its competitive inhibitor maltose. FEBS J. 2010, 277, 4205–4214. [Google Scholar] [CrossRef]
- Lu, H.; Qi, Y.; Zhao, Y.; Jin, N. Effects of hydroxyl group on the interaction of carboxylated flavonoid derivatives with S. cerevisae α-glucosidase. Curr. Comput.-Aided Des. 2020, 16, 31–44. [Google Scholar]
- Hadni, H.; Elhallaoui, M. Molecular docking and QSAR studies for modeling the antimalarial activity of hybrids 4-anilinoquinoline-triazines derivatives with the wild-type and mutant receptor pf-DHFR. Heliyon 2019, 5, e02357. [Google Scholar] [CrossRef] [Green Version]
- Ninomiya, M.; Koketsu, M. Minor Flavonoids (Chalcones, Flavanones, Dihydrochalcones, and Aurones); Ramawat, K.G., Mérillon, J.-M., Eds.; Natural Products; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1867–1900. [Google Scholar]
- Wink, M. Evolution of secondary metabolites in legumes (Fabaceae). S. Afr. J. Bot. 2013, 89, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Mirossay, L.; Varinská, L.; Mojžiš, J. Antiangiogenic effect of flavonoids and chalcones: An update. Int. J. Mol. Sci. 2018, 19, 27. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chetty, S.; Soliman, M.E. Possible allosteric binding site on Gyrase B, a key target for novel anti-TB drugs: Homology modelling and binding site identification using molecular dynamics simulation and binding free energy calculations. Med. Chem. Res. 2015, 24, 2055–2074. [Google Scholar] [CrossRef]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosco, P.; Stiefl, N.; Landrum, G. Bringing the MMFF force field to the RDKit: Implementation and validation. J. Cheminform. 2014, 6, 37. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-a visualization system for exploratory research and analysis. J. Comput Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biovia Discovery Studio. Discovery Studio Visualizer; Biovia Discovery Studio: San Diego, CA, USA, 2017; Volume 936. [Google Scholar]
- Phoopha, S.; Wattanapiromsakul, C.; Pitakbut, T.; Dej-adisai, S. Chemical constituents of Litsea elliptica and their alpha-glucosidase inhibition with molecular docking. Pharmacog. Mag. 2020, 16, 327–334. [Google Scholar]
- Rais, I.R. Phytochemical Investigation and Anti-diabetic Activity of Bauhinia pulla and Bauhinia winitii. Ph.D. Thesis, Prince of Songkla University, Hatyai, Songkhla, Thailand, 2020. [Google Scholar]

{kind=link}
{kind=link}
| Part | Extract | Alpha-Glucosidase Inhibition (%) ± SD | IC50 (µg/mL) |
|---|---|---|---|
| Leaves | n-Hexane (HLBP) | 15.09 ± 3.09 | - |
| Ethyl acetate (EALBP) | 52.34 ± 4.41 | 529.60 | |
| Ethanol (ETLBP) | 97.44 ± 0.92 | 138.95 | |
| Water (WLBP) | 54.08 ± 3.30 | 641.66 | |
| Woods | n-Hexane (HWBP) | 29.36 ± 2.12 | - |
| Ethyl acetate (EAWBP) | 58.73 ± 0.98 | 957.13 | |
| Ethanol (ETWBP) | 69.53 ± 2.29 | 693.98 | |
| Water (WWBP) | 58.59 ± 3.06 | 655.36 | |
| Standard | Acarbose | 86.09 ± 1.54 | 193.37 |
| Compounds | Extract | IC50 (µg/mL) | Proportion * |
|---|---|---|---|
| Acarbose | - | 124.11 | - |
| Quercetin | EALBP, ETLBP, ETWBP | 5.41 | 22.94 |
| Quercitrin | ETLBP | 49.69 | 2.50 |
| Luteolin | ETWBP | 164.05 | 0.76 |
| 5-deoxyluteolin | ETWBP | 574.57 | 0.22 |
| 4-methyl ether isoliquiritigenin | EAWBP, ETWBP | 220.05 | 0.56 |
| 3,2’,4’-trihydroxy-4-methoxychalcone | ETWBP | 1273.31 | 0.10 |
| Steroids | EALBP, HWBP, ETWBP | 1392.81 | 0.09 |
| Steroids glycoside | ETLBP | 8204.20 | 0.02 |
| Cpd. | Vdw + Hbond Energy (kcal/mol) [1] | Electrostatic Energy (kcal/mol) [2] | Desolvation Energy (kcal/mol) [3] | Intermolecular Energy (kcal/mol) [4] | Total Internal Energy (kcal/mol) [5] | Torsional Free Energy (kcal/mol) [6] | Unbound System’s Energy (kcal/mol) [7] | Estimated Free Energy of Binding (kcal/mol) [4 + 5+6-7] |
|---|---|---|---|---|---|---|---|---|
| 7 | −8.31 | 0.17 | 2.77 | −5.36 | −0.40 | 1.79 | 0.00 | −3.97 |
| 8 | −8.39 | 0.20 | 2.70 | −5.49 | −0.67 | 2.09 | 0.00 | −4.07 |
| Δ | 0.08 | 0.03 | 0.07 | 0.12 | 0.27 | 0.30 * | 0.00 | 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dej-adisai, S.; Rais, I.R.; Wattanapiromsakul, C.; Pitakbut, T. Alpha-Glucosidase Inhibitory Assay-Screened Isolation and Molecular Docking Model from Bauhinia pulla Active Compounds. Molecules 2021, 26, 5970. https://doi.org/10.3390/molecules26195970
Dej-adisai S, Rais IR, Wattanapiromsakul C, Pitakbut T. Alpha-Glucosidase Inhibitory Assay-Screened Isolation and Molecular Docking Model from Bauhinia pulla Active Compounds. Molecules. 2021; 26(19):5970. https://doi.org/10.3390/molecules26195970
Chicago/Turabian StyleDej-adisai, Sukanya, Ichwan Ridwan Rais, Chatchai Wattanapiromsakul, and Thanet Pitakbut. 2021. "Alpha-Glucosidase Inhibitory Assay-Screened Isolation and Molecular Docking Model from Bauhinia pulla Active Compounds" Molecules 26, no. 19: 5970. https://doi.org/10.3390/molecules26195970