
Inhibitory Effect of Thai Purple Rice Husk Extract on Chemically Induced Carcinogenesis in Rats

 ,
,  ,
, 
Abstract
:
1. Introduction
2. Results
2.1. Chemical Constituents in Purple Rice Husk Extract
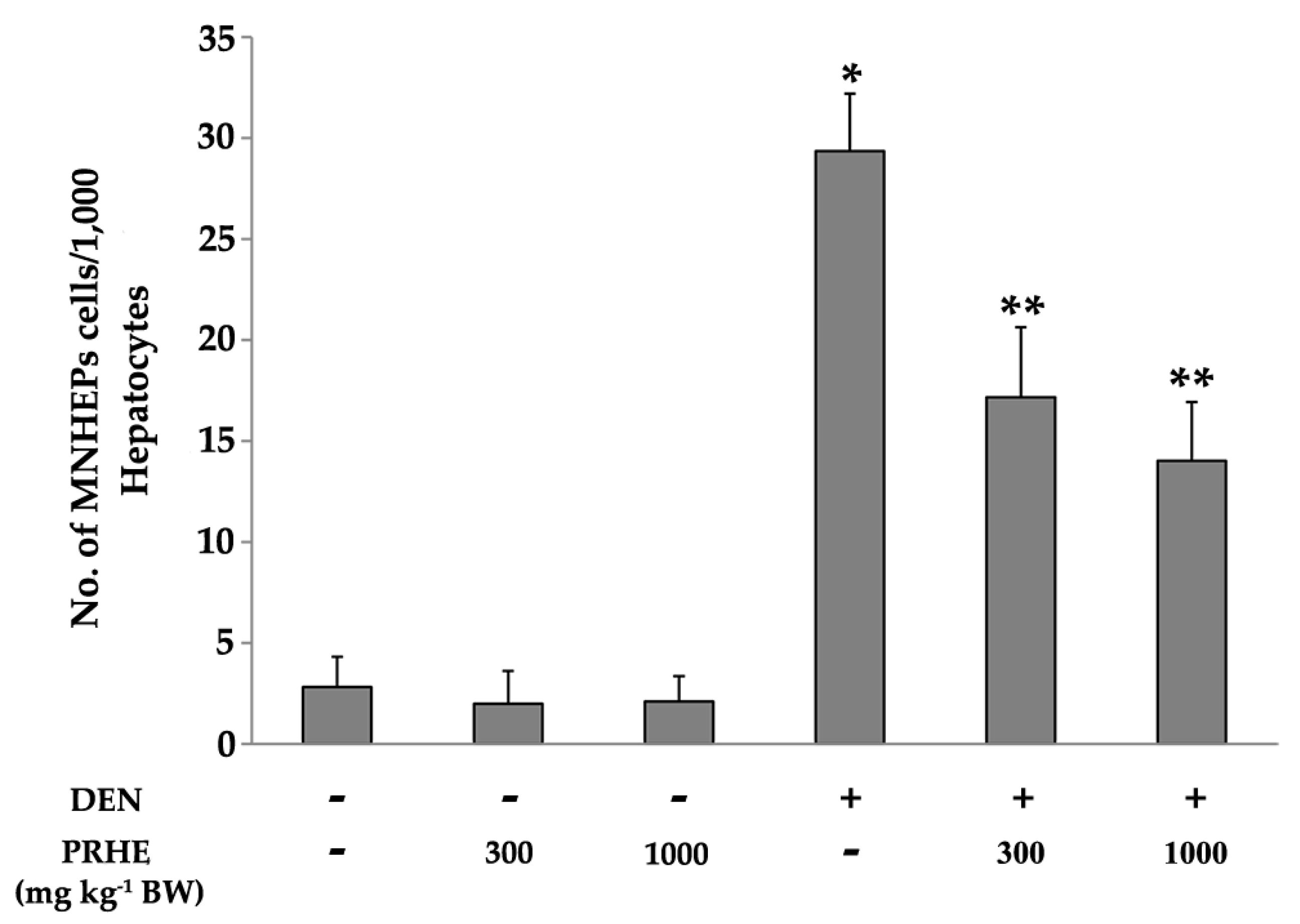
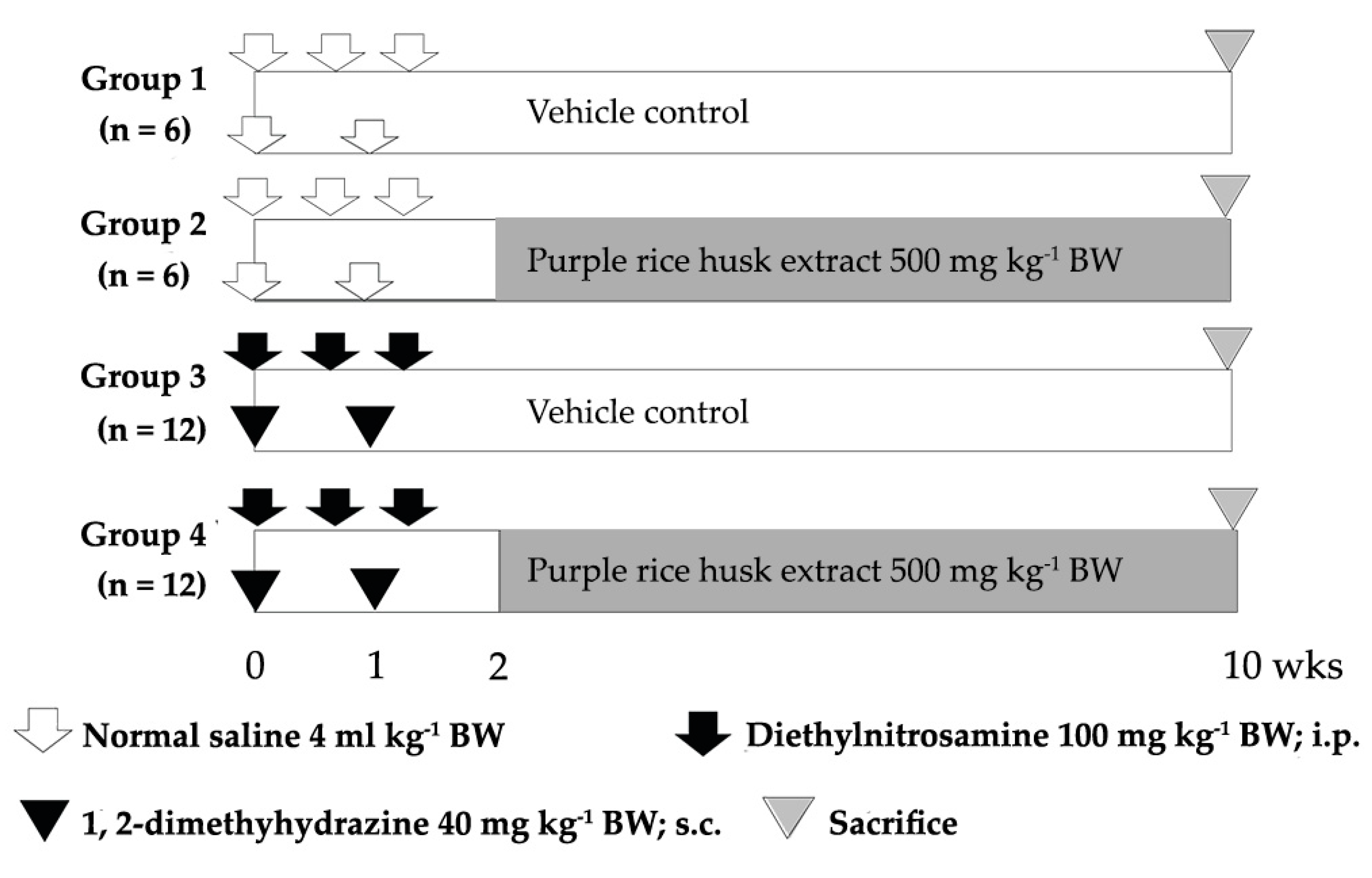
2.2. Acute Toxicity, Clastogenicity, and Anticlastogenicity of Purple Rice Husk Extract
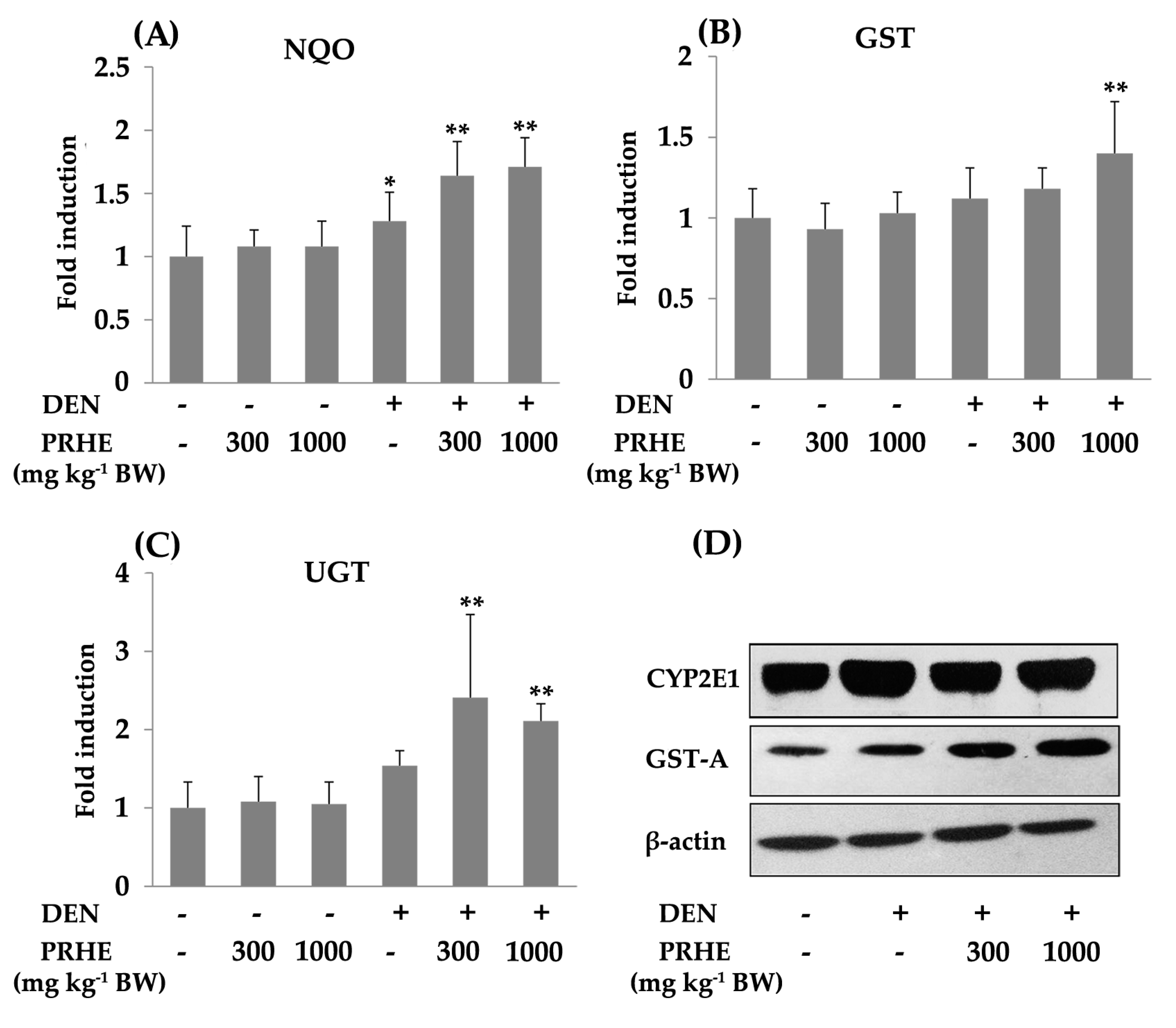
2.3. Carcinogenicity and Anticarcinogenicity of Purple Rice Husk Extract
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of Purple Rice Husk Extract
4.3. Determination of Some Chemical Constituents in Purple Rice Husk Extract
4.4. Animals
4.5. Acute Toxicity Test
4.6. Short-Term Carcinogenicity Test
4.7. Determination of Phases I and II Xenobiotic-Metabolizing Enzymes
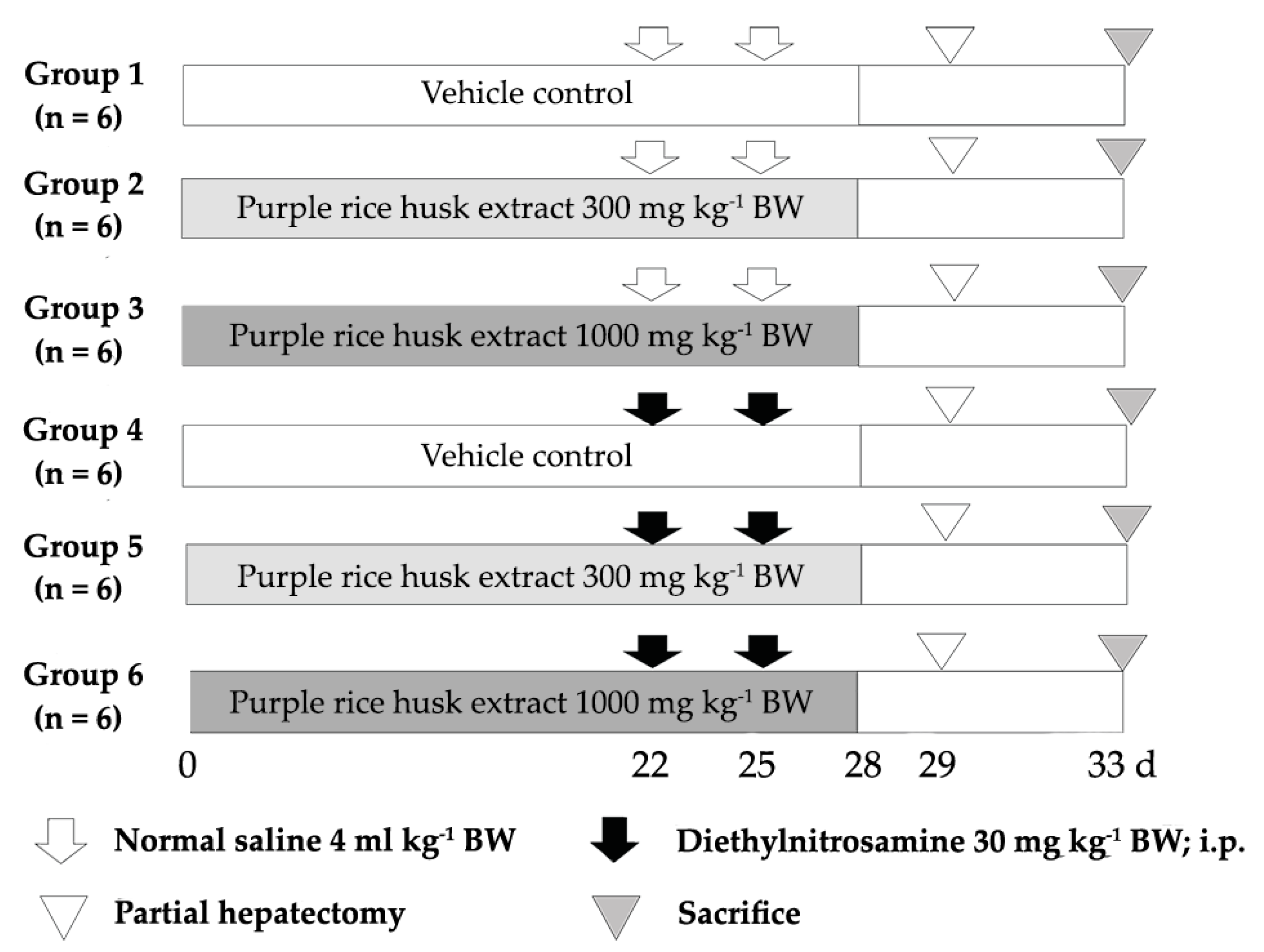
4.8. Medium-Term Liver and Colon Carcinogenicity Test
4.9. Determination of Preneoplastic Lesions in Liver and Colon
4.10. Determination of Proliferating Cell Nuclear Antigen and Apoptotic Hepatocytes by Double-Staining Immunohistochemistry
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pourhoseingholi, M.A.; Vahedi, M.; Baghestani, A.R. Burden of gastrointestinal cancer in Asia: An overview. Gastroenterol. Hepatol. Bed Bench 2015, 8, 19–27. [Google Scholar]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Varzakas, T.; Zakynthinos, G.; Verpoort, F. Plant food residues as a source of nutraceuticals and functional foods. Foods 2016, 5, 88. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, H.; Liang, H.; Yuan, Q. Purification, antioxidant activity and protein-precipitating capacity of punicalin from pomegranate husk. Food Chem. 2013, 138, 437–443. [Google Scholar] [CrossRef]
- Xiang, L.; Wang, Y.; Yi, X.; Wang, X.; He, X. Chemical constituent and antioxidant activity of the husk of Chinese hickory. J. Funct. Foods 2016, 23, 378–388. [Google Scholar] [CrossRef]
- Barreca, D.; Laganà, G.; Leuzzi, U.; Smeriglio, A.; Trombetta, D.; Bellocco, E. Evaluation of the nutraceutical, antioxidant and cytoprotective properties of ripe pistachio (Pistacia vera L., variety Bronte) hulls. Food Chem. 2016, 196, 493–502. [Google Scholar] [CrossRef]
- Park, J.Y.; Shin, M.S.; Kim, S.N.; Kim, H.Y.; Kim, K.H.; Shin, K.S.; Kang, K.S. Polysaccharides from Korean Citrus hallabong peels inhibit angiogenesis and breast cancer cell migration. Int. J. Biol. Macromol. 2016, 85, 522–529. [Google Scholar] [CrossRef]
- Esa, N.M.; Ling, T.B.; Peng, L.S. By-products of rice processing: An overview of health benefits and applications. Rice Res. 2013, 1, 1–11. [Google Scholar]
- Wanyo, P.; Meeso, N.; Siriamornpun, S. Effects of different treatments on the antioxidant properties and phenolic compounds of rice bran and rice husk. Food Chem. 2014, 157, 457–463. [Google Scholar] [CrossRef]
- Nilnumkhum, A.; Punvittayagul, C.; Chariyakornkul, A.; Wongpoomchai, R. Effects of hydrophilic compounds in purple rice husk on AFB1-induced mutagenesis. Mol. Cell Toxicol. 2017, 13, 171–178. [Google Scholar] [CrossRef]
- Punvittayagul, C.; Sringarm, K.; Chaiyasut, C.; Wongpoomchai, R. Mutagenicity and antimutagenicity of hydrophilic and lipophilic extracts of Thai northern purple rice. Asian Pac. J. Cancer Prev. 2014, 15, 9517–9522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busat, S.; Siriamornpun, S. Phenolic acids and antioxidant activities in husk of different Thai rice varieties. Food Sci. Technol. Int. 2020, 16, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Chariyakornkul, A.; Punvittayagul, C.; Taya, S.; Wongpoomchai, R. Inhibitory effect of purple rice husk extract on AFB1-induced micronucleus formation in rat liver through modulation of xenobiotic metabolizing enzymes. BMC Complement. Altern. Med. 2019, 19, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, N.J.; Choi, K.C.; Lee, S.A.; Baek, J.A.; Lee, J.C. Rice hull extracts inhibit proliferation of MCF-7 cells with G1 cell cycle arrest in parallel with their antioxidant activity. J. Med. Food 2015, 18, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Park, H.R.; Park, E.; Lee, S.C. Cytotoxic and antitumor activity of momilactone B from rice hulls. J. Agric. Food Chem. 2007, 55, 1702–1706. [Google Scholar] [CrossRef]
- Sankam, P.; Punvittayagul, C.; Sringam, K.; Chaiyasut, C.; Wongpoomchai, R. Antimutagenicity and anticlastogenicity of glutinous purple rice hull using in vitro and in vivo testing systems. Mol. Cell Toxicol. 2013, 9, 169–176. [Google Scholar] [CrossRef]
- Xu, Z.; Hua, N.; Godber, J.S. Antioxidant activity of tocopherols, tocotrienols, and gamma-oryzanol components from rice bran against cholesterol oxidation accelerated by 2,2′-azobis(2-methylpropionamidine) dihydrochloride. J. Agric. Food Chem. 2001, 49, 2077–2081. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E. Scientific evidence of rice by-products for cancer prevention: Chemopreventive properties of waste products from rice milling on carcinogenesis in vitro and in vivo. Biomed Res. Int. 2017, 2017, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.N.; Chu, S.C.; Chiou, H.L.; Chiang, C.L.; Yang, S.F.; Hsieh, Y.S. Cyanidin-3-glucoside and peonidin-3-glucoside inhibit tumor cell growth and induce apoptosis in vitro and suppress tumor growth in vivo. Nutr. Cancer 2005, 53, 232–243. [Google Scholar] [CrossRef]
- Tomankova, E.; Balik, J.; Soural, I.; Bednar, P.; Papouskova, B. Colour and antioxidant properties of malvidin-3-glucoside and vitisin A. Acta Aliment. 2016, 45, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steward, W.P.; Brown, K. Cancer chemoprevention: A rapidly evolving field. Br. J. Cancer 2013, 109, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Wanibuchi, H.; Morimura, K.; Gonzalez, F.J.; Fukushima, S. Role of CYP2E1 in diethylnitrosamine-induced hepatocarcinogenesis in vivo. Cancer Res. 2007, 67, 11141–11146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.C.; Lee, S.; Cho, M.H. The simplest flowchart stating the mechanisms for organic xenobiotics-induced toxicity: Can it possibly be accepted as a “central dogma” for toxic mechanisms? Toxicol. Res. 2014, 30, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Mason, E.F.; Rathmell, J.C. Cell metabolism: An essential link between cell growth and apoptosis. Biochim. Biophys. Acta 2011, 1813, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Al-Ishaq, R.K.; Overy, A.J.; Büsselberg, D. Phytochemicals and gastrointestinal cancer: Cellular mechanisms and effects to change cancer progression. Biomolecules 2020, 10, 105. [Google Scholar] [CrossRef] [Green Version]
- Hollman, P.C.H. Absorption, bioavailability, and metabolism of flavonoids. Pharm. Biol. 2004, 42, 74–83. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Li, Y.W.; Wang, D.; Li, X.G.; Jin, Y. Anthocyanins extracted from Chinese blueberry and its anticancer effects on HepG2 cells. Adv. Mater. Res. 2014, 887, 592–595. [Google Scholar] [CrossRef]
- Tanaka, T.; Tanaka, T.; Tanaka, M. Potential cancer chemopreventive activity of protocatechuic acid. J. Exp. Clin. Med. 2011, 3, 27–33. [Google Scholar] [CrossRef]
- Krajka-Kuzniak, V.; Kaczmarek, J.; Baer-Dubowska, W. Effect of naturally occurring phenolic acids on the expression of glutathione S-transferase isozymes in the rat. Food Chem. Toxicol. 2008, 46, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zhou, S.; Yang, S. Vanillic acid suppresses HIF-1α expression via inhibition of mTOR/p70S6K/4E-BP1 and Raf/MEK/ERK pathways in human colon cancer HCT116 cells. Int. J. Mol. Sci. 2019, 20, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.S.; Luo, P.; Zeng, Z.; Wang, H.; Malafa, M.; Suh, N. Vitamin E and cancer prevention: Studies with different forms of tocopherols and tocotrienols. Mol. Carcinsog. 2020, 59, 365–389. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Tang, M.; Wu, J. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar]
- OECD. Test No: 425 Acute Oral Toxicity: Up and Down Procedure. In OECD Guidelines for the Testing of Chemicals, Section 4; OECD Publishing: Paris, France, 2008. [Google Scholar]
- Punvittayagul, C.; Chariyakornkul, A.; Chewonarin, T.; Jarukamjorn, K.; Wongpoomchai, R. Augmentation of diethylnitrosamine-induced early stages of rat hepatocarcinogenesis b 1,2-dimethylhydrazine. Drug Chem. Toxicol. 2019, 42, 641–648. [Google Scholar] [CrossRef]
- Thumvijit, T.; Taya, S.; Punvittayagul, C.; Peerapornpisal, Y.; Wongpoomchai, R. Cancer chemopreventive effect of Spirogyra neglecta (Hassall) Kützing on diethylnitrosamine-induced hepatocarcinogenesis in rats. Asian Pac. J. Cancer Prev. 2014, 15, 1611–1616. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytes | Content (mg g−1 Extract) |
|---|---|
| α-Tocopherol | 0.75 ± 0.02 |
| β-Tocopherol | 0.08 ± 0.00 |
| γ-Tocopherol | 0.13 ± 0.00 |
| δ-Tocopherol | 0.03 ± 0.01 |
| γ-Tocotrienol | 0.10 ± 0.00 |
| Protocatechuic acid | 0.45 ± 0.01 |
| Vanillic acid | 1.53 ± 0.02 |
| p-Coumaric acid | 0.54 ± 0.00 |
| Ferulic acid | 0.06 ± 0.00 |
| Rutin | 0.17 ± 0.05 |
| Cyanidin-3-glucoside | 0.32 ± 0.00 |
| Peonidin-3-glucoside | 0.55 ± 0.01 |
| Malvidin-3-glucoside | 0.71 ± 0.02 |
| Delphinidin-3-glucoside | 0.04 ± 0.00 |
| Cyanidin | 0.56 ± 0.05 |
| Peonidin | 0.04 ± 0.00 |
| Treatment | Preneoplastic Lesions | |||
|---|---|---|---|---|
| Liver | Colon | |||
| No. of GST-P+ foci/cm2 | Area of GST-P+ Foci (mm2/cm2) | Aberrant Crypt/Rat | Aberrant Crypt/Focus | |
| NSS | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| NSS + PRHE 500 mg kg−1 BW | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| DEN + DMH | 17.45 ± 3.97 * | 1.84 ± 0.74 * | 169.56 ± 70.97 * | 4.45 ± 1.00 * |
| DEN + DMH + PRHE 500 mg kg−1 BW | 11.06 ± 4.89 ** | 1.37 ± 0.81 | 226.43 ± 112.61 | 4.68 ± 0.55 |
| Treatment | PCNA Labeling Index (%) | Apoptotic Labeling Index (%) | ||
|---|---|---|---|---|
| GST-P-Positive Foci | Surrounding Area | GST-P-Positive Foci | Surrounding Area | |
| NSS | 0.00 ± 0.00 | 2.81 ± 0.52 | 0.00 ± 0.00 | 0.30 ± 0.19 |
| NSS + PRHE 500 mg kg−1 BW | 0.00 ± 0.00 | 2.07 ± 0.55 | 0.00 ± 0.00 | 0.34 ± 0.13 |
| DEN + DMH | 6.67 ± 2.81 * | 4.20 ± 1.39 | 0.42 ± 0.37 | 0.40 ± 0.15 |
| DEN + DMH + PRHE 500 mg kg−1 BW | 3.38 ± 1.71 ** | 2.50 ± 0.90 ** | 0.72 ± 0.44 | 0.99 ± 0.58 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Punvittayagul, C.; Chariyakornkul, A.; Sankam, P.; Wongpoomchai, R. Inhibitory Effect of Thai Purple Rice Husk Extract on Chemically Induced Carcinogenesis in Rats. Molecules 2021, 26, 360. https://doi.org/10.3390/molecules26020360
Punvittayagul C, Chariyakornkul A, Sankam P, Wongpoomchai R. Inhibitory Effect of Thai Purple Rice Husk Extract on Chemically Induced Carcinogenesis in Rats. Molecules. 2021; 26(2):360. https://doi.org/10.3390/molecules26020360
Chicago/Turabian StylePunvittayagul, Charatda, Arpamas Chariyakornkul, Paweena Sankam, and Rawiwan Wongpoomchai. 2021. "Inhibitory Effect of Thai Purple Rice Husk Extract on Chemically Induced Carcinogenesis in Rats" Molecules 26, no. 2: 360. https://doi.org/10.3390/molecules26020360