Novel Methods to Manipulate Autolysis in Sparkling Wine: Effects on Yeast

 ,
,  ,
,
Abstract
1. Introduction
2. Results
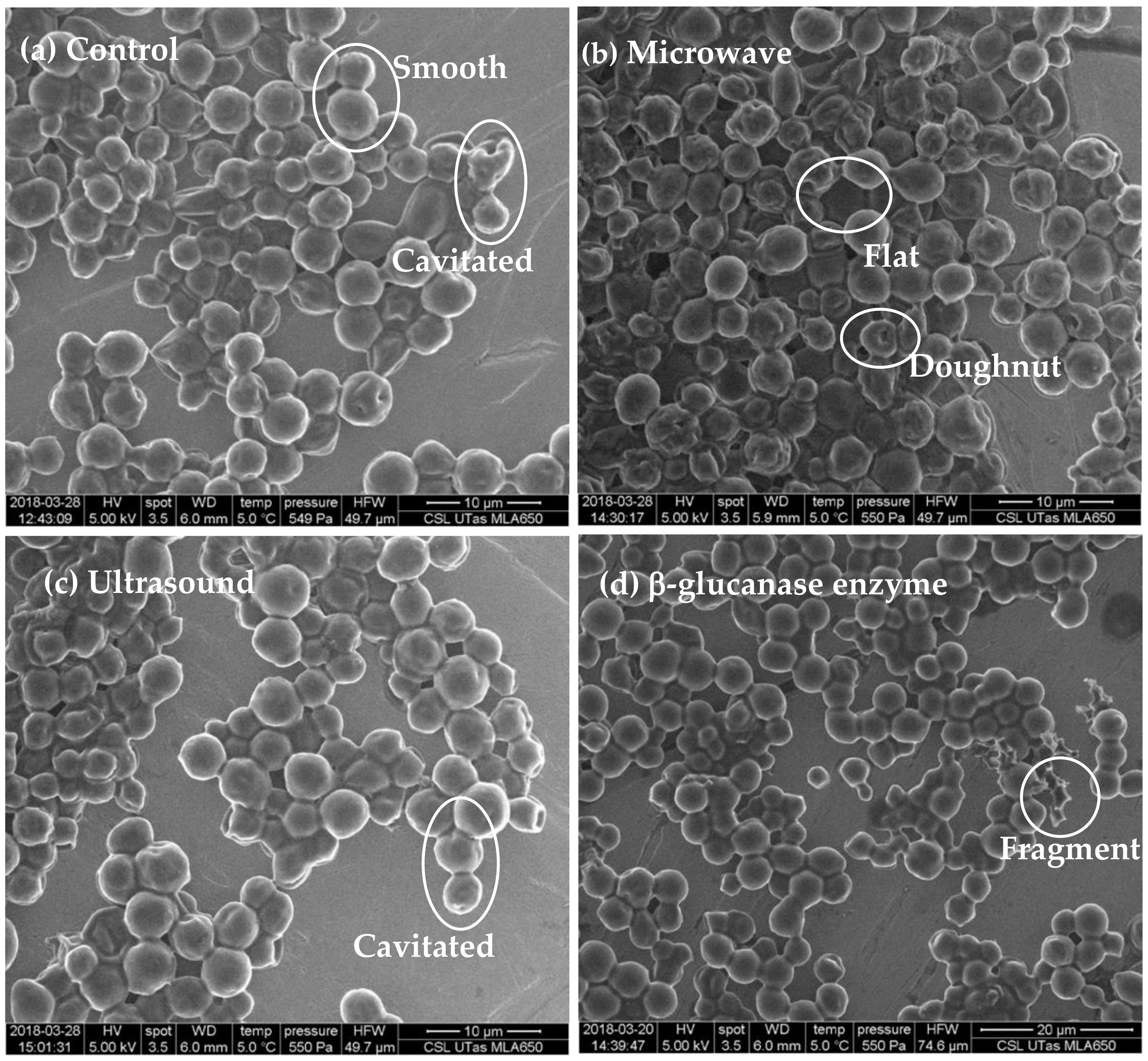
2.1. Scanning Electron Microscopy Observation of Yeast Cells
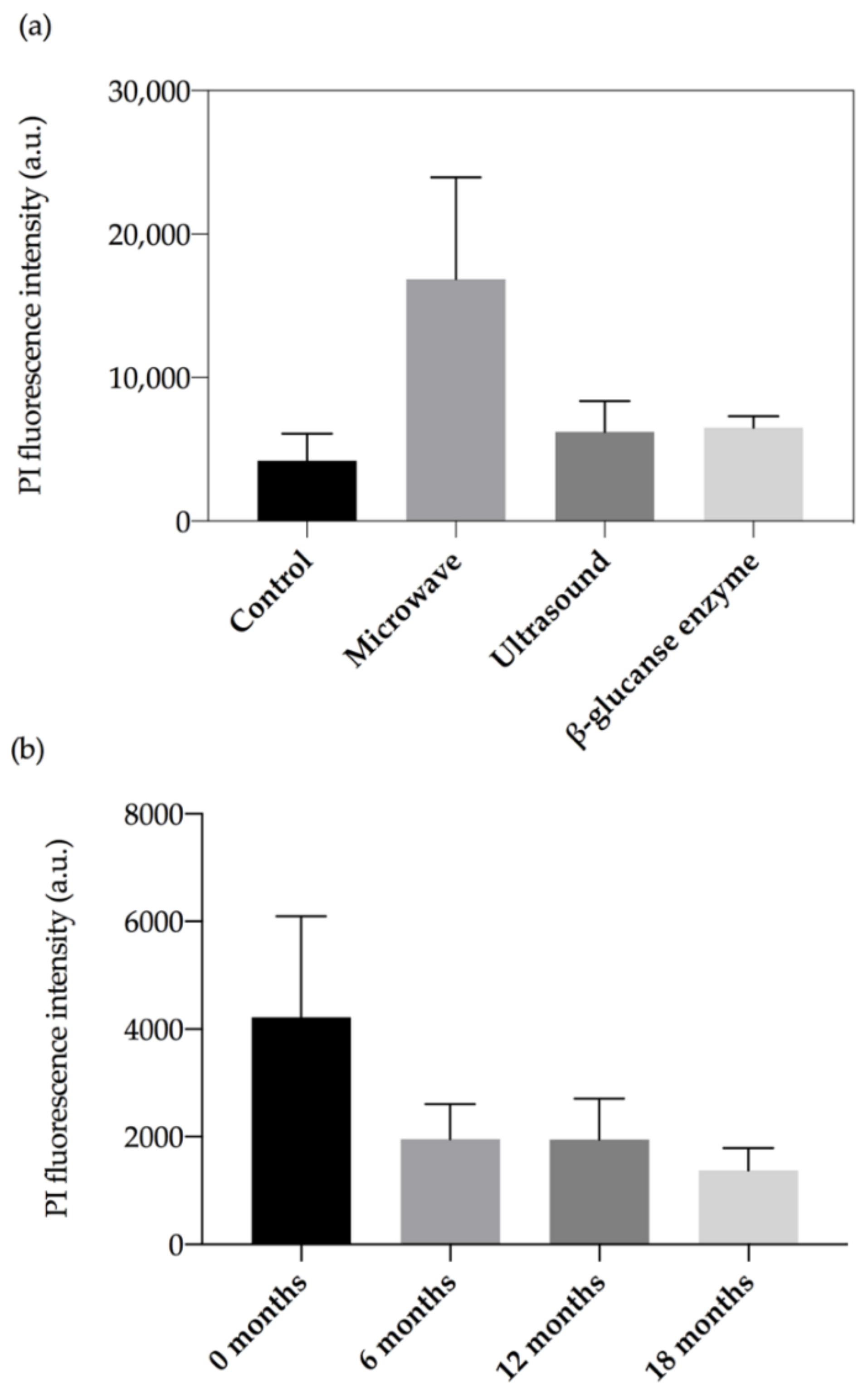
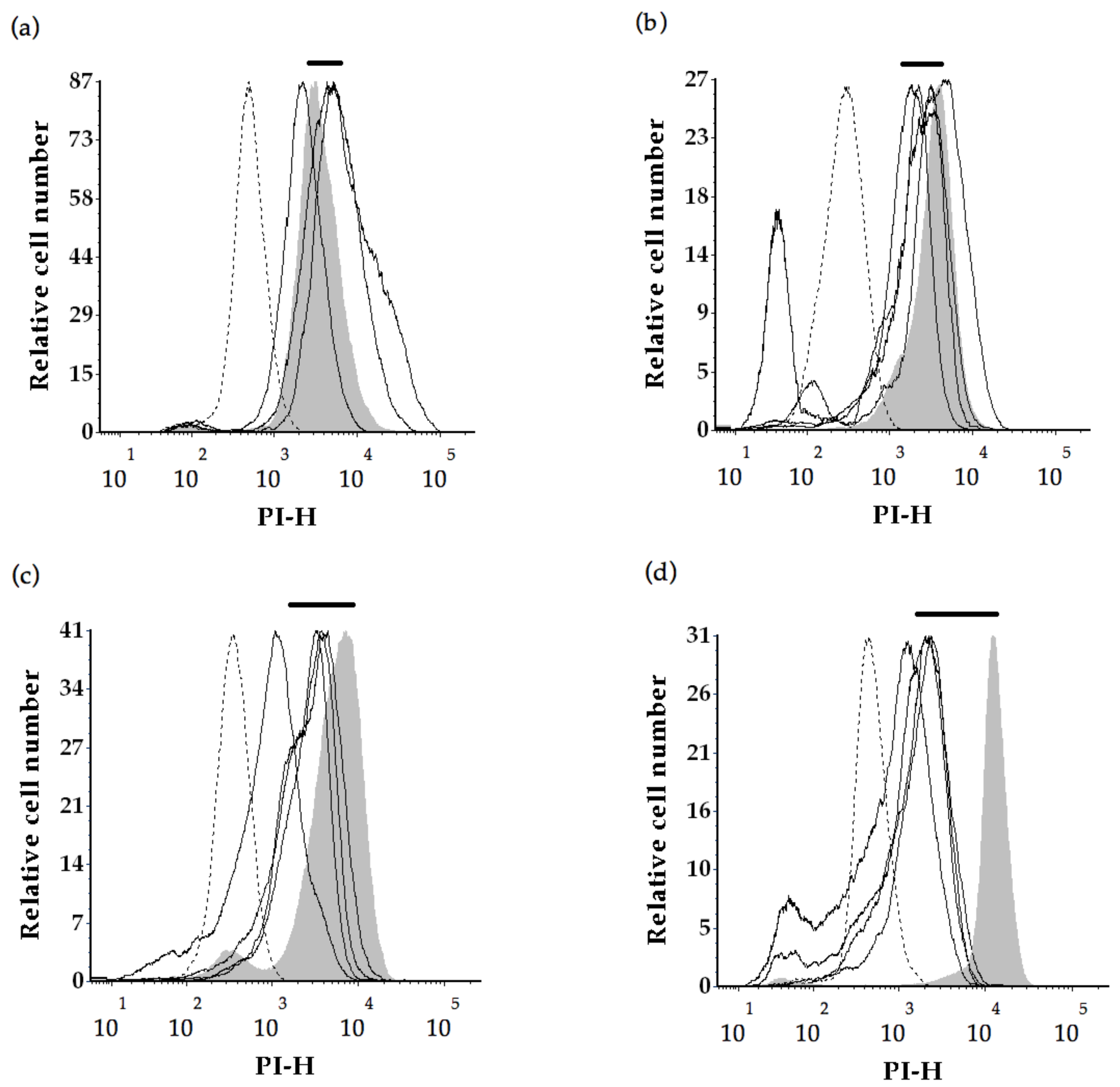
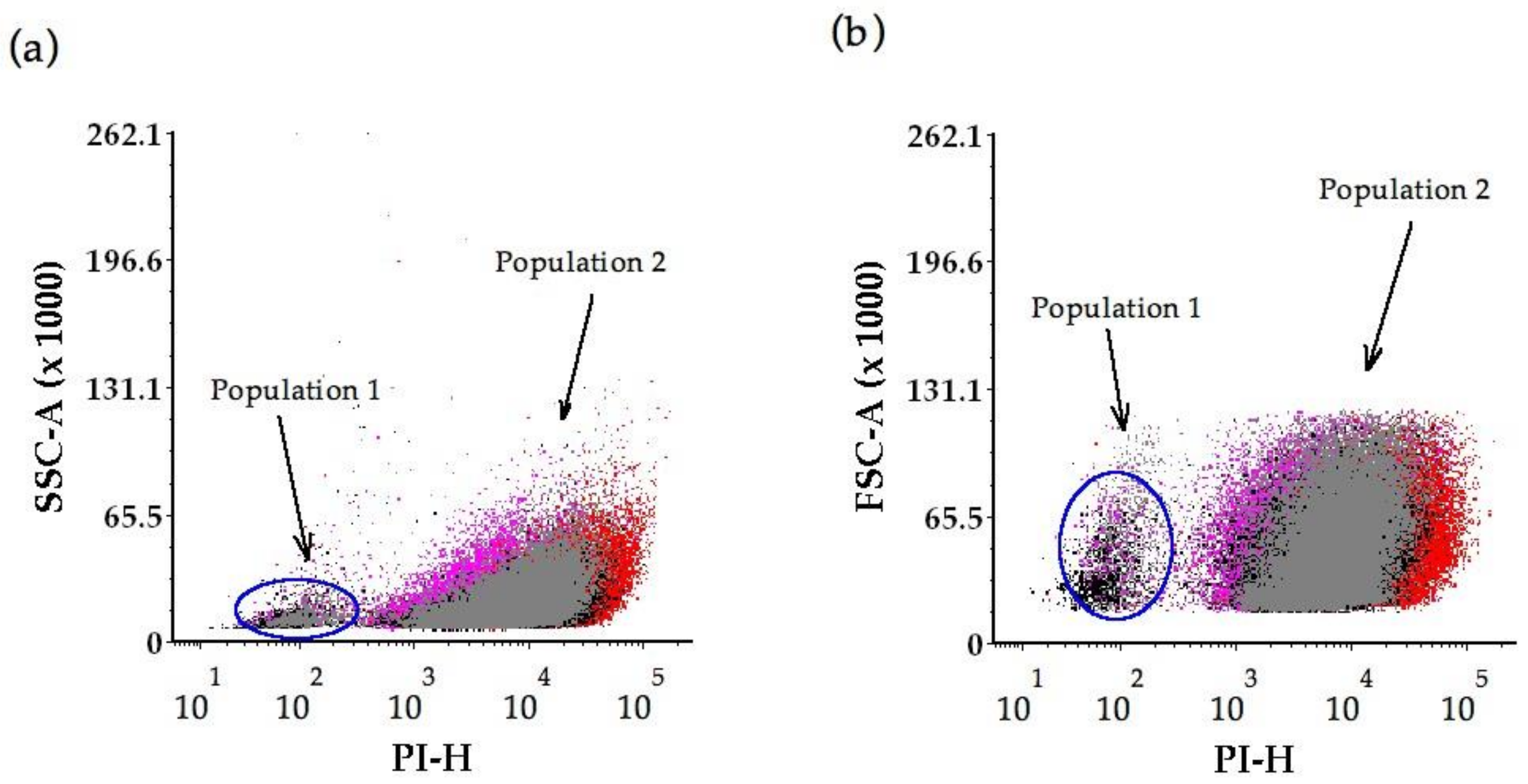
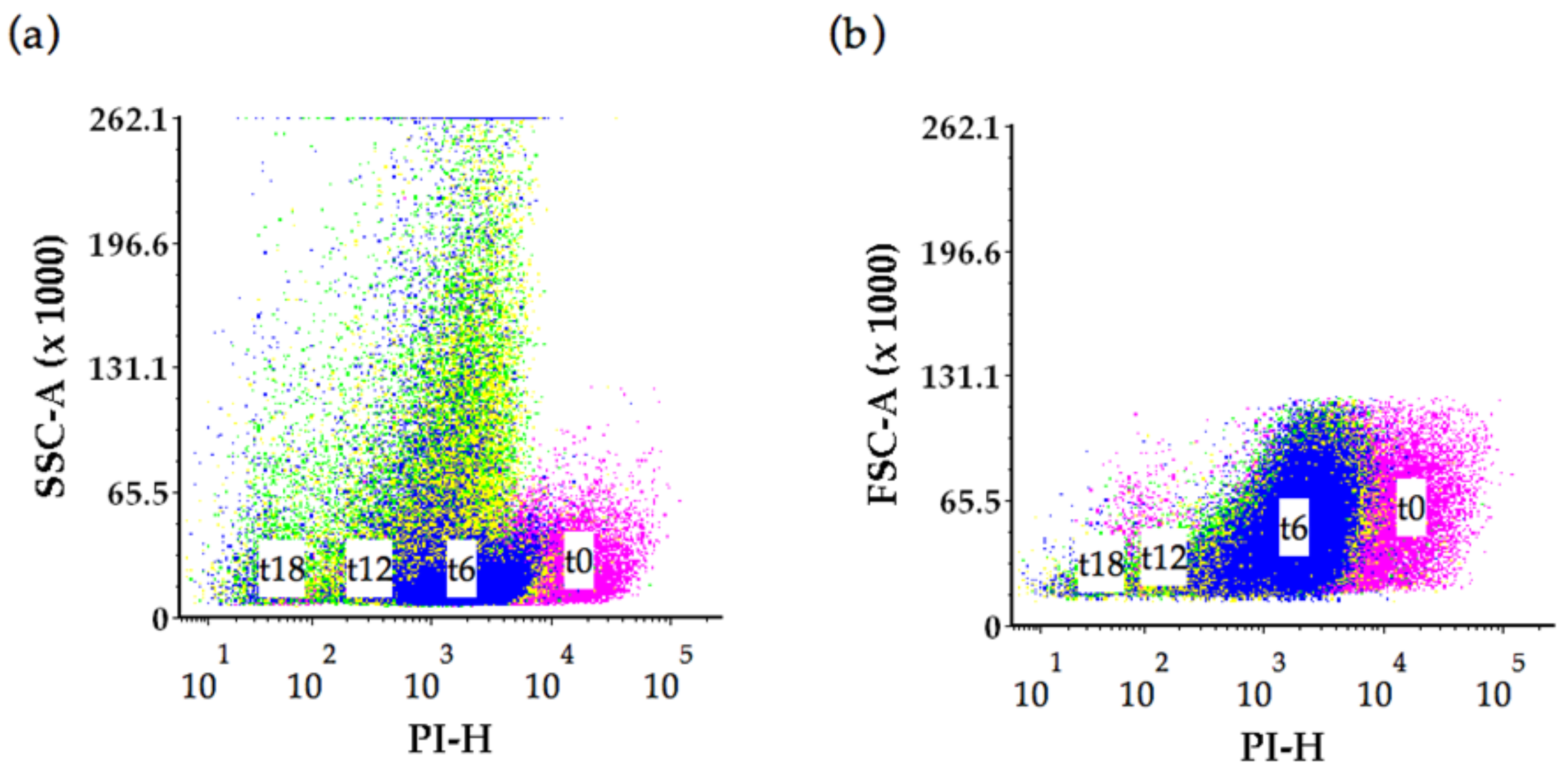
2.2. Flow Cytometry Analysis
3. Discussion
3.1. Morphological Features of Yeast Cells
3.2. Changes in Yeast Morphological Features During Secondary Fermentation with aging of Sparkling Wine
3.3. Effects of Novel Treatments on Yeast Morphological Features
3.4. Yeast Cell Integrity as Measured by PI Permeability
4. Materials and Methods
4.1. Sparkling Wine Production
4.1.1. Yeast Culture
4.1.2. Yeast Treatments
4.1.3. Preparation of Sparkling Wines
4.2. Scanning Electron Microscopy Analyses
4.3. Image Analyses
4.4. Flow Cytometry Analyses
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Charpentier, C.; Feuillat, M. Yeast autolysis. In The Yeasts; Rose, A.H., Harrison, J.S., Eds.; Academic Press: New York, NY, USA, 1992; Volume III, pp. 315–386. [Google Scholar]
- Moreno-Arribas, M.V.; Pueyo, E.; Nieto, F.J.; Martin-Avarez, P.J.; Polo, M.C. Influence of the polysaccharides and the nitrogen compounds on foaming properties of sparkling wines. Food Chem. 2000, 70, 309–317. [Google Scholar] [CrossRef]
- Nunez, Y.P.; Carrascosa, A.V.; Gonzalez, R.; Polo, M.C.; Martinez-Rodriguez, A.J. Effect of accelerated autolysis of yeast on the composition and foaming properties of sparkling wines elaborated by a champenoise method. J. Agric. Food Chem. 2005, 53, 7232–7237. [Google Scholar] [CrossRef]
- Alexandre, H. Yeast and sparkling wine production. In Yeasts in the Production of Wine; Springer: Berlin, Germany, 2019; pp. 395–432. [Google Scholar]
- Alexandre, H.; Guilloux-Benatier, M. Yeast autolysis in sparkling wine-a review. Aust. J. Grape Wine Res. 2006, 12, 119–127. [Google Scholar] [CrossRef]
- Todd, B.E.N.; Fleet, G.H.; Henschke, P.A. Promotion of autolysis through the interaction of killer and sensitive yeasts: Potential application in sparkling wine production. Am. J. Enol. Vitic. 2000, 51, 65–72. [Google Scholar]
- Leroy, M.J.; Charpentier, M.; Duteurtre, B.; Feuillat, M.; Charpentier, C. Yeast Autolysis during Champagne Aging. Am. J. Enol. Vitic. 1990, 41, 21–28. [Google Scholar]
- Babayan, T.L.; Bezrukov, M.G. Autolysis in Yeasts. Acta Biotechnol. 1985, 5, 129–136. [Google Scholar] [CrossRef]
- Marti-Raga, M.; Martin, V.; Gil, M.; Sancho, M.; Zamora, F.; Mas, A.; Beltran, G. Contribution of yeast and base wine supplementation to sparkling wine composition. J. Sci. Food Agric. 2016, 96, 4962–4972. [Google Scholar] [CrossRef]
- Martinez-Lapuente, L.; Guadalupe, Z.; Ayestaran, B.; Ortega-Heras, M.; Perez-Magarino, S. Changes in polysaccharide composition during sparkling wine making and aging. J. Agric. Food Chem. 2013, 61, 12362–12373. [Google Scholar] [CrossRef]
- Cebollero, E.; Carrascosa, A.V.; Gonzalez, R. Evidence for yeast autophagy during simulation of sparkling wine aging: A reappraisal of the mechanism of yeast autolysis in wine. Biotechnol. Prog. 2005, 21, 614–616. [Google Scholar] [CrossRef]
- Charpentier, C.; Aussenac, J.; Charpentier, M.; Prome, J.C.; Duteurtre, B.; Feuillat, M. Release of nucleotides and nucleosides during yeast autolysis: Kinetics and potential impact on flavor. J. Agric. Food Chem. 2005, 53, 3000–3007. [Google Scholar] [CrossRef]
- Perpetuini, G.; Di Gianvito, P.; Arfelli, G.; Schirone, M.; Corsetti, A.; Tofalo, R.; Suzzi, G. Biodiversity of autolytic ability in flocculent Saccharomyces cerevisiae strains suitable for traditional sparkling wine fermentation. Yeast 2016, 33, 303–312. [Google Scholar] [CrossRef]
- Lombardi, S.J.; De Leonardis, A.; Lustrato, G.; Testa, B.; Iorizzo, M. Yeast Autolysis in Sparkling Wine Aging: Use of Killer and Sensitive Saccharomyces cerevisiae Strains in Co-Culture. Recent Pat. Biotechnol. 2016, 9, 223–230. [Google Scholar] [CrossRef]
- La Gatta, B.; Picariello, G.; Rutigliano, M.; Lopriore, G.; Petrella, G.; Rusco, G.; Tremonte, P. Di Luccia, A. Addition of lees from base wine in the production of Bombino sparkling wine. Eur. Food Res. Technol. 2016, 242, 1307–1317. [Google Scholar] [CrossRef]
- Comuzzo, P.; Calligaris, S.; Iacumin, L.; Ginaldi, F.; Voce, S.; Zironi, R. Application of multi-pass high pressure homogenization under variable temperature regimes to induce autolysis of wine yeasts. Food Chem. 2017, 224, 105–113. [Google Scholar] [CrossRef]
- Voce, S.; Calligaris, S.; Comuzzo, P. Effect of a yeast autolysate produced by high pressure homogenization on white wine evolution during ageing. J. Food Sci. Technol. 2020, 1–10. [Google Scholar] [CrossRef]
- Khoddami, A.; Wilkes, M.A.; Roberts, T.H. Techniques for analysis of plant phenolic compounds. Molecules 2013, 18, 2328–2375. [Google Scholar] [CrossRef]
- Liu, L.; Loira, I.; Morata, A.; Suárez-Lepe, J.A.; González, M.C.; Rauhut, D. Shortening the ageing on lees process in wines by using ultrasound and microwave treatments both combined with stirring and abrasion techniques. Eur. Food Res. Technol. 2016, 242, 559–569. [Google Scholar] [CrossRef]
- Colagrande, O.; Silva, A.; Fumi, M.D. Recent Applications of Biotechnology in Wine Production. Biotechnol. Prog. 1994, 10, 2–18. [Google Scholar] [CrossRef]
- Aguilar, T.; Loyola, C.; De Bruijn, J.; Bustamante, L.; Vergara, C.; Von Baer, D.; Mardones, C.; Serra, I. Effect of thermomaceration and enzymatic maceration on phenolic compounds of grape must enriched by grape pomace, vine leaves and canes. Eur. Food Res. Technol. 2015, 242, 1149–1158. [Google Scholar] [CrossRef]
- Rodriguez-Nogales, J.M.; Fernández-Fernández, E.; Vila-Crespo, J. Effect of the addition of β-glucanases and commercial yeast preparations on the chemical and sensorial characteristics of traditional sparkling wine. Eur. Food Res. Technol. 2012, 235, 729–744. [Google Scholar] [CrossRef]
- Rodriguez-Nogales, J.M.; Fernandez-Fernandez, E.; Gomez, M.; Vila-Crespo, J. Antioxidant properties of sparkling wines produced with beta-glucanases and commercial yeast preparations. J. Food Sci. 2012, 77, 1005–1010. [Google Scholar] [CrossRef]
- Torresi, S.; Frangipane, M.T.; Garzillo, A.M.V.; Massantini, R.; Contini, M. Effects of a β-glucanase enzymatic preparation on yeast lysis during aging of traditional sparkling wines. Food Res. Int. 2014, 55, 83–92. [Google Scholar] [CrossRef]
- Feuillat, M. Yeast macromolecules: Origin, composition, and enological interest. Am. J. Enol. Vitic. 2003, 54, 211–213. [Google Scholar]
- Palomero, F.; Morata, A.; Benito, S.; Gonzalez, M.C.; Suarez-Lepe, J.A. Conventional and enzyme-assisted autolysis during ageing over lees in red wines: Influence on the release of polysaccharides from yeast cell walls and on wine monomeric anthocyanin content. Food Chem. 2007, 105, 838–846. [Google Scholar] [CrossRef]
- Martínez-Rodríguez, A.J.; Pueyo, E. Sparkling wines and yeast autolysis. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, M.C., Eds.; Springer: New York, NY, USA, 2009; pp. 61–80. [Google Scholar]
- Martı́nez-Rodrı́guez, A.J.; Polo, M.C.; Carrascosa, A.V. Structural and ultrastructural changes in yeast cells during autolysis in a model wine system and in sparkling wines. Int. J. Food Microbiol. 2001, 71, 45–51. [Google Scholar] [CrossRef]
- Martinez-Rodriguez, A.J.; Gonzalez Garcia, R.; Carrascosa, A.V. Morphological Changes in Autolytic Wine Yeast during Aging in Two Model Systems. J. Food Sci. 2004, 69, 233–239. [Google Scholar] [CrossRef]
- Tudela, R.; Gallardo-Chacon, J.J.; Rius, N.; Lopez-Tamames, E.; Buxaderas, S. Ultrastructural changes of sparkling wine lees during long-term aging in real enological conditions. FEMS Yeast Res. 2012, 12, 466–476. [Google Scholar] [CrossRef]
- Piton, F.; Charpentier, M.; Troton, D. Cell Wall and Lipid Changes in Saccharomyces cerevisiae during Aging of Champagne Wine. Am. J. Enol. Vitic. 1988, 39, 221–226. [Google Scholar]
- Prudencio, C.; Sansonetty, F. Corte-Real, M. Flow cytometric assessment of cell structural and functional changes induced by acetic acid in the yeasts Zygosaccharomyces bailii and Saccharomyces cerevisiae. Cytometry 1998, 31, 307–313. [Google Scholar] [CrossRef]
- Branco, P.; Monteiro, M.; Moura, P.; Albergaria, H. Survival rate of wine-related yeasts during alcoholic fermentation assessed by direct live/dead staining combined with fluorescence in situ hybridization. Int. J. Food Microbiol. 2012, 158, 49–57. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W. Definition, classification and nomenclature of the yeasts. In The Yeasts, A Taxonomic Study, 4th ed.; Kurtzman, C.P., Fell, J.W., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Garcia Martin, J.F.; Guillemet, L.; Feng, C.; Sun, D.W. Cell viability and proteins release during ultrasound-assisted yeast lysis of light lees in model wine. Food Chem. 2013, 141, 934–939. [Google Scholar] [CrossRef]
- Feuillat, M.; Charpentier, C. Autolysis of Yeasts in Champagne. Am. J. Enol. Vitic. 1982, 33, 6–13. [Google Scholar]
- Charpentier, C.; Freyssinet, M. The mechanism of autolysis in wine. Yeast 1989, 5, 181–186. [Google Scholar]
- Mehmood, N.; Husson, E.; Jacquard, C.; Wewetzer, S.; Buchs, J.; Sarazin, C.; Gosselin, I. Impact of two ionic liquids, 1-ethyl-3-methylimidazolium acetate and 1-ethyl-3-methylimidazolium methylphosphonate, on Saccharomyces cerevisiae: Metabolic, physiologic, and morphological investigations. Biotechnol. Biofuels Biomed. Cent. 2015, 8, 1–17. [Google Scholar] [CrossRef]
- Tang, S.Y.; Zhang, W.; Soffe, R.; Nahavandi, S.; Shukla, R.; Khoshmanesh, K. High resolution scanning electron microscopy of cells using dielectrophoresis. PLoS ONE 2014, 9, e104109. [Google Scholar] [CrossRef]
- Vincent, M.; Suali, L.; Azahari, A.; Rowena, P.; Baran, M.; Nillian, E.; Bilung, L.M. Growth Kinetics of Ethidium Bromide Mutagenized Lipomyces starkeyi Strains. J. Adv. Res. Appl. Sci. Eng. Technol. 2018, 11, 47–54. [Google Scholar]
- Guzzon, R.; Larcher, R. The application of flow cytometry in microbiological monitoring during winemaking: Two case studies. Ann. Microbiol. 2015, 65, 1865–1878. [Google Scholar] [CrossRef]
- Longin, C.; Petitgonnet, C.; Guilloux-Benatier, M.; Rousseaux, S.; Alexandre, H. Application of flow cytometry to wine microorganisms. Food Microbiol. 2017, 62, 221–231. [Google Scholar] [CrossRef]
- Laurent, M.; Valade, M. La preparation du levain de tirage: A partir de levures seches actives. Le Vign. Champen. 2007, 128, 74–95. [Google Scholar]
- Cacciola, V.; Batllò, I.F.; Ferraretto, P.; Vincenzi, S.; Celotti, E. Study of the ultrasound effects on yeast lees lysis in winemaking. Eur. Food Res. Technol. 2012, 236, 311–317. [Google Scholar] [CrossRef]
- Kirk, S.E.; Skepper, J.N.; Donald, A.M. Application of environmental scanning electron microscopy to determine biological surface structure. J. Microsc. 2009, 233, 205–224. [Google Scholar] [CrossRef]
- Arena, E.T.; Rueden, C.T.; Hiner, M.C.; Wang, S.; Yuan, M.; Eliceiri, K.W. Quantitating the cell: Turning images into numbers with ImageJ. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6, 1–18. [Google Scholar] [CrossRef]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Description |
|---|---|
| Smooth | Whole-cells with smooth external surfaces, mother-daughter cells, and budding, generally 1 to 5 μm size diameter |
| Cavitated | Modification of the cell surface to varying degrees creating an uneven cell surface |
| Flat | Yeast cells positioned flat on the SEM sample holder. Such cells were not classified into the ‘smooth’ or ‘cavitated’ or ‘doughnut’-shaped categories |
| Pitted/Porous | Indentations in the cell surface giving an impression of holes in the surface |
| Doughnut-shaped | Whole-cells where the centre appears to have been removed, a cell shaped like a doughnut or bagel |
| Fragments | Irregular small features thought to result from breakages or remains of cells that are smaller than whole-cells, generally less than 1 μm size diameter |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gnoinski, G.B.; Schmidt, S.A.; Close, D.C.; Goemann, K.; Pinfold, T.L.; Kerslake, F.L. Novel Methods to Manipulate Autolysis in Sparkling Wine: Effects on Yeast. Molecules 2021, 26, 387. https://doi.org/10.3390/molecules26020387
Gnoinski GB, Schmidt SA, Close DC, Goemann K, Pinfold TL, Kerslake FL. Novel Methods to Manipulate Autolysis in Sparkling Wine: Effects on Yeast. Molecules. 2021; 26(2):387. https://doi.org/10.3390/molecules26020387
Chicago/Turabian StyleGnoinski, Gail B., Simon A. Schmidt, Dugald C. Close, Karsten Goemann, Terry L. Pinfold, and Fiona L. Kerslake. 2021. "Novel Methods to Manipulate Autolysis in Sparkling Wine: Effects on Yeast" Molecules 26, no. 2: 387. https://doi.org/10.3390/molecules26020387
APA StyleGnoinski, G. B., Schmidt, S. A., Close, D. C., Goemann, K., Pinfold, T. L., & Kerslake, F. L. (2021). Novel Methods to Manipulate Autolysis in Sparkling Wine: Effects on Yeast. Molecules, 26(2), 387. https://doi.org/10.3390/molecules26020387