
Characterization of Cross-Linked Enzyme Aggregates of the Y509E Mutant of a Glycoside Hydrolase Family 52 β-xylosidase from G. stearothermophilus

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Optimization of XynB2Y509E CLEAs Preparation
2.1.1. Optimization of Precipitant Type and Precipitation Time
2.1.2. Effect of Concentration and pH in Cross-Linking Reaction
2.2. Biochemical Characterization of Free XynB2Y509E and XynB2Y509E-CLEAs
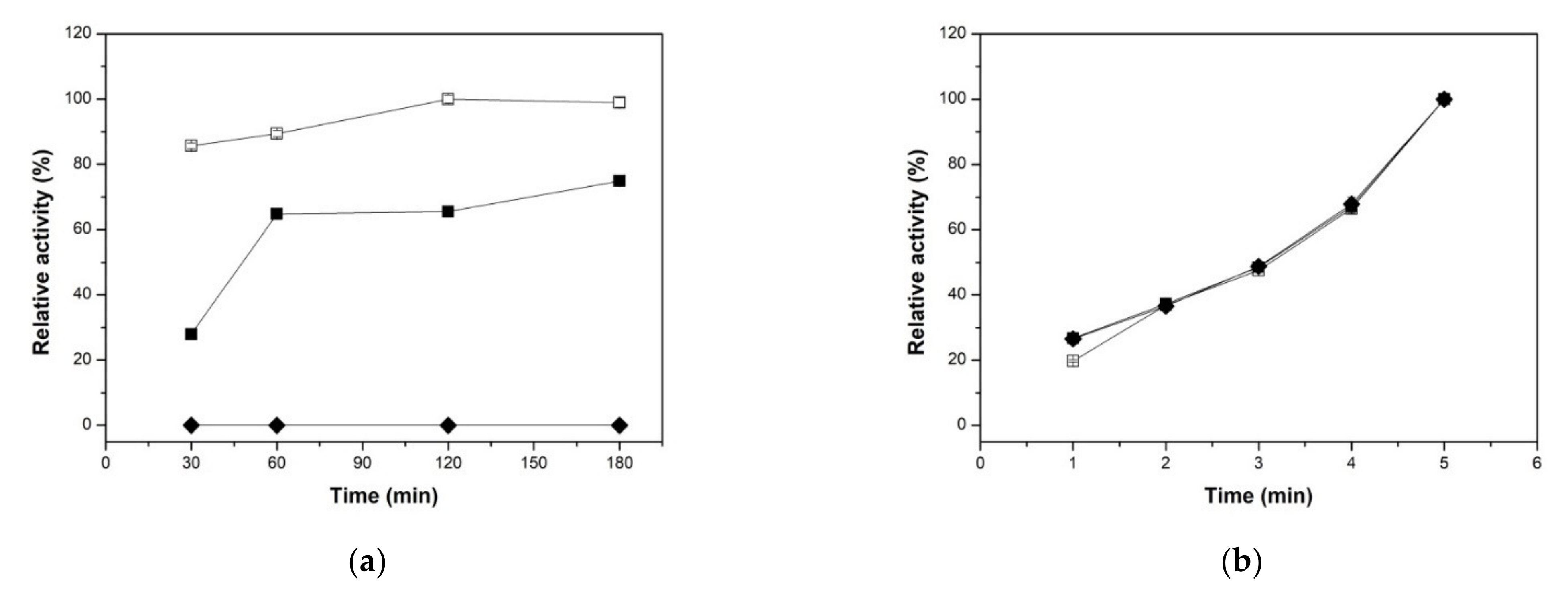
2.2.1. Effect of Reaction Times on both Xylanolytic Activities
2.2.2. Effect of the Temperature and pH on Activity and Stability
2.2.3. Kinetic Parameters for the Free XynB2Y509E and XynB2Y509E-CLEAs
2.2.4. Reusability
3. Materials and Methods
3.1. Materials
3.2. Mutagenesis of the xynB2 Gene
- 5-GCGCGCAACAATTTAGAGTTGACAGGAAAAT-3′
- 5-ATTTTCCTGTCAACTCTAAATTGTTGCGCGC-3′
3.3. Overexpression and Partial Purification of XynB2Y509E
3.4. Enzymatic Assays
3.5. Preparation of XynB2Y509E-CLEAs
3.6. Optimization of XynB2Y509E-CLEAs Preparation
3.7. Scanning Electron Microscopy of XynB2Y509E-CLEAs
3.8. Biochemical Characterization of XynB2Y509E-CLEAs
3.8.1. Optimal Temperature and Thermal Stability of Free and XynB2Y509E-CLEAs
3.8.2. Effect of pH on Activity
3.8.3. Kinetic Parameters of Free and XynB2Y509E-CLEAs
3.8.4. Recyclability of XynB2Y509E-CLEAs
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Xu, M.Q.; Wang, S.S.; Li, L.N.; Gao, J.; Zhang, Y.W. Combined cross-linked enzyme aggregates as biocatalysts. Catalysts 2018, 8, 460. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, R.A. Cross-linked enzyme aggregates (CLEA® s): Stable and recyclable biocatalysts. Biochem. Soc. Trans. 2007, 35, 1583–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheldon, R.A. Characteristic features and biotechnological applications of cross-linked enzyme aggregates (CLEAs). Appl. Microbiol. Biotechnol. 2011, 92, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talekar, S.; Joshi, A.; Joshi, G.; Kamat, P.; Haripurkar, R.; Kambale, S. Parameters in preparation and characterization of cross linked enzyme aggregates (CLEAs). RSC Adv. 2013, 3, 12485–12511. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Kiyota, Y.; Miyazaki, M. Techniques for preparation of cross-linked enzyme aggregates and their applications in bioconversions. Catalysts 2018, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, R.A. CLEAs, combi-CLEAs and ‘smart’ magnetic CLEAs: Biocatalysis in a bio-based economy. Catalysts 2019, 9, 261. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hu, Y.D.; Li, N.; Zong, M.H. Cross-linked Enzyme Aggregates of β-glucosidase from Prunus domestica seeds. Biotechnol. Lett. 2012, 34, 1673–1678. [Google Scholar] [CrossRef]
- Li, L.; Li, G.; Cao, L.C.; Ren, G.H.; Kong, W.; Wang, S.D.; Guo, G.S.; Liu, Y.H. Characterization of the cross-linked enzyme aggregates of a novel β-galactosidase, a potential catalyst for the synthesis of galacto-oligosaccharides. J. Agric. Food Chem. 2015, 63, 894–901. [Google Scholar] [CrossRef]
- Hero, J.S.; Romero, C.M.; Pisa, J.H.; Perotti, N.I.; Olivaro, C.; Martinez, M.A. Designing cross-linked xylanase aggregates for bioconversion of agroindustrial waste biomass towards potential production of nutraceuticals. Int. J. Biol. Macromol. 2018, 111, 229–236. [Google Scholar] [CrossRef]
- Verma, R.; Kumar, A.; Kumar, S. Synthesis and characterization of cross-linked enzyme aggregates (CLEAs) of thermostable xylanase from Geobacillus thermodenitrificans X1. Process. Biochem. 2019, 80, 72–79. [Google Scholar] [CrossRef]
- Periyasamy, K.; Santhalembi, L.; Mortha, G.; Aurousseau, M.; Subramanian, S. Carrier-free co-immobilization of xylanase, cellulase and β-1,3-glucanase as combined cross-linked enzyme aggregates (combi-CLEAs) for one-pot saccharification of sugarcane bagasse. RSC Adv. 2016, 6, 32849–32857. [Google Scholar] [CrossRef]
- Xu, M.Q.; Li, F.L.; Yu, W.Q.; Li, R.F.; Zhang, Y.W. Combined cross-linked enzyme aggregates of glycerol dehydrogenase and NADH oxidase for high efficiency in situ NAD+ regeneration. Int J. Biol Macromol. 2020, 144, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Bravman, T.; Zolotnitsky, G.; Shulami, S.; Belakhov, V.; Solomon, D.; Baasov, T.; Shoham, G.; Shoham, Y. Stereochemistry of family 52 glycosyl hydrolases: A β-xylosidase from Bacillus stearothermophilus T-6 is a retaining enzyme. FEBS Lett. 2001, 495, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Bravman, T.; Zolotnitsky, G.; Belakhov, V.; Shoham, G.; Henrissat, B.; Baasov, T.; Shoham, Y. Detailed kinetic analysis of a family 52 glycoside hydrolase: A β-xylosidase from Geobacillus stearothermophilus. Biochemistry 2003, 42, 10528–10536. [Google Scholar] [CrossRef] [PubMed]
- Bravman, T.; Belakhov, V.; Solomon, D.; Shoham, G.; Henrissat, B.; Baasov, T.; Shoham, Y. Identification of the catalytic residues in family 52 glycoside hydrolase, a β-xylosidas from Geobacillus stearothermophilus T-6. J. Biol. Chem. 2003, 278, 26742–26749. [Google Scholar] [CrossRef] [Green Version]
- Contreras, L.M.; Gómez, J.; Prieto, J.; Clemente-Jiménez, J.M.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F.; Blanco, F.; Neira, J.L. The family 52 β-xylosidase from Geobacillus stearothermophilus is a dimer: Structural and biophysical characterization of a glycoside hydrolase. Biochim. Biophys. Acta. 2008, 1784, 1924–1934. [Google Scholar] [CrossRef]
- Kurz, L.; García, V.; Wilkesman, J.; Contreras, L.M. Enzymatic characterization of the recombinant beta-xylosidase XynB2. J. Medical Biol. Sci. 2014, 1, 14–19. [Google Scholar]
- Ben-David, A.; Bravman, T.; Balazs, Y.S.; Czjzek, M.; Schomburg, D.; Shoham, G.; Shoham, Y. Glycosynthase activity of Geobacillus stearothermophilus GH52 b-xylosidase: Efficient synthesis of xylooligosaccharides from alpha-D-xylopyranosyl fluoride through a conjugated reaction. Chembiochem 2007, 8, 2145–2151. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, X.; Zhang, S.; Liu, Z. GH52 xylosidase from Geobacillus stearothermophilus: Characterization and introduction of xylanase activity by site-directed mutagenesis of Tyr509. J. Ind. Microbiol. Biotechnol. 2014, 41, 65–74. [Google Scholar] [CrossRef]
- Zhen, Q.; Wang, M.; Qi, W.; Su, R.; He, Z. Preparation of β-mannanase CLEAs using macromolecular cross-linkers. Catal. Sci. Technol. 2013, 3, 1937–1941. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Pletschke, B.I. Magnetic cross-linked enzyme aggregates (CLEAs): A novel concept towards carrier free immobilization of lignocellulolytic enzymes. Enzyme Microb. Technol. 2014, 61–62, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Pletschke, B.I. Strategic optimization of xylanase–mannanase combi-CLEAs for synergistic and efficient hydrolysis of complex lignocellulosic substrates. J. Mol. Catal. B Enzym. 2015, 115, 140–150. [Google Scholar] [CrossRef]
- Schoevaart, R.; Wolbers, M.W.; Golubovic, M.; Ottens, M.; Kieboom, A.P.G.; Van Rantwijk, F.; van der Wielen, L.A.M.; Sheldon, R.A. Preparation, optimization, and structures of cross-linked enzyme aggregates (CLEAs). Biotechnol. Bioeng. 2004, 87, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Hwangbo, M.; Tran, J.L.; Chu, K.H. Effective one-step saccharification of lignocellulosic biomass using magnetite-biocatalysts containing saccharifying enzymes. Sci. Total Environ. 2019, 647, 806–813. [Google Scholar] [CrossRef]
- Migneault, I.; Dartiguenave, C.; Bertrand, M.J.; Waldron, K.C. Glutaraldehyde: Behavior in aqueous solution, reaction with proteins, and application to enzyme crosslinking. Biotechniques 2004, 37, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.W.; Chen, H.; Wang, X.; Yang, Y.Y.; Ching, C.B. Cross-linked enzyme aggregates (CLEAs) with controlled particles: Application to Candida rugosa lipase. J. Mol. Catal. B Enzym. 2006, 43, 124–127. [Google Scholar] [CrossRef]
- Rojas, M.J.; Amaral-Fonseca, M.; Zanin, G.M.; Fernandez-Lafuente, R.; Giordano, R.L.C.; Tardioli, P.W. Preparation of Crosslinked Enzyme Aggregates of a Thermostable Cyclodextrin Glucosyltransferase from Thermoanaerobacter sp. Critical Effect of the Crosslinking Agent. Catalysts 2019, 9, 120. [Google Scholar] [CrossRef] [Green Version]
- Bibi, Z.; Qader, S.A.U.; Aman, A. Calcium alginate matrix increases the stability and recycling capability of immobilized endo-β-1,4-xylanase from Geobacillus stearothermophilus KIBGE-IB29. Extremophiles 2015, 19, 819–827. [Google Scholar] [CrossRef]
- Hormigo, D.; García-Hidalgo, J.; Acebal, C.; de la Mata, I.; Arroyo, M. Preparation and characterization of cross-linked enzyme aggregates (CLEAs) of recombinant poly-3-hydroxybutyrate depolymerase from Streptomyces exfoliatus. Bioresour. Technol. 2012, 115, 177–182. [Google Scholar] [CrossRef]
- Vršanská, M.; Voběrková, S.; Jiménez Jiménez, A.M.; Strmiska, V.; Adam, V. Preparation and optimisation of cross-linked enzyme aggregates using native isolate white rot fungi Trametes versicolor and Fomes fomentarius for the decolourisation of synthetic dyes. Int. J. Environ. Res. Public Health. 2018, 15, 23. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.Y.; Yang, Y.; Yang, Z. Activity and stability of cross-linked tyrosinase aggregates in aqueous and nonaqueous media. J. Biotechnol. 2011, 152, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.D.; Zhang, S.; Sun, L.M. Cross-linked enzyme aggregates of phenylalanine ammonia lyase: Novel biocatalysts for synthesis of L-phenylalanine. Appl. Biochem. Biotechnol. 2012, 167, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Zhao, L.; Huang, Y.; Tan, X. Preparation of cross-linked aggregates of aminoacylase from Aspergillus melleus by using bovine serum albumin as an inert additive. Bioresour. Technol. 2010, 101, 6569–6571. [Google Scholar] [CrossRef] [PubMed]
- Montoro-García, S.; Gil-Ortiz, F.; Navarro-Fernández, J.; Rubio, V.; García-Carmona, F.; Sánchez-Ferrer, Á. Improved cross-linked enzyme aggregates for the production of desacetyl β-lactam antibiotics intermediates. Bioresour. Technol. 2010, 101, 331–336. [Google Scholar] [CrossRef]
- Zhao, L.; Zheng, L.; Gao, G.; Jia, F.; Cao, S. Resolution of N-(2-ethyl-6-methylphenyl) alanine via cross-linked aggregates of Pseudomonas sp. lipase. J. Mol. Catal. B Enzym. 2008, 54, 7–12. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Km (mM) | Vmax (µmol/min) |
|---|---|---|
| Free XynB2Y509E | 0.9 ± 0.1 | 1.6 ± 0.2 |
| XynB2Y509E-CLEAs | 1.2 ± 0.2 | 1.18 ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero, G.; Contreras, L.M.; Aguirre, C.; Wilkesman, J.; Clemente-Jiménez, J.M.; Rodríguez-Vico, F.; Las Heras-Vázquez, F.J. Characterization of Cross-Linked Enzyme Aggregates of the Y509E Mutant of a Glycoside Hydrolase Family 52 β-xylosidase from G. stearothermophilus. Molecules 2021, 26, 451. https://doi.org/10.3390/molecules26020451
Romero G, Contreras LM, Aguirre C, Wilkesman J, Clemente-Jiménez JM, Rodríguez-Vico F, Las Heras-Vázquez FJ. Characterization of Cross-Linked Enzyme Aggregates of the Y509E Mutant of a Glycoside Hydrolase Family 52 β-xylosidase from G. stearothermophilus. Molecules. 2021; 26(2):451. https://doi.org/10.3390/molecules26020451
Chicago/Turabian StyleRomero, Gabriela, Lellys M. Contreras, Carolina Aguirre, Jeff Wilkesman, Josefa María Clemente-Jiménez, Felipe Rodríguez-Vico, and Francisco Javier Las Heras-Vázquez. 2021. "Characterization of Cross-Linked Enzyme Aggregates of the Y509E Mutant of a Glycoside Hydrolase Family 52 β-xylosidase from G. stearothermophilus" Molecules 26, no. 2: 451. https://doi.org/10.3390/molecules26020451