
Functional Component Isolated from Phaseolus vulgaris Lectin Exerts In Vitro and In Vivo Anti-Tumor Activity through Potentiation of Apoptosis and Immunomodulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
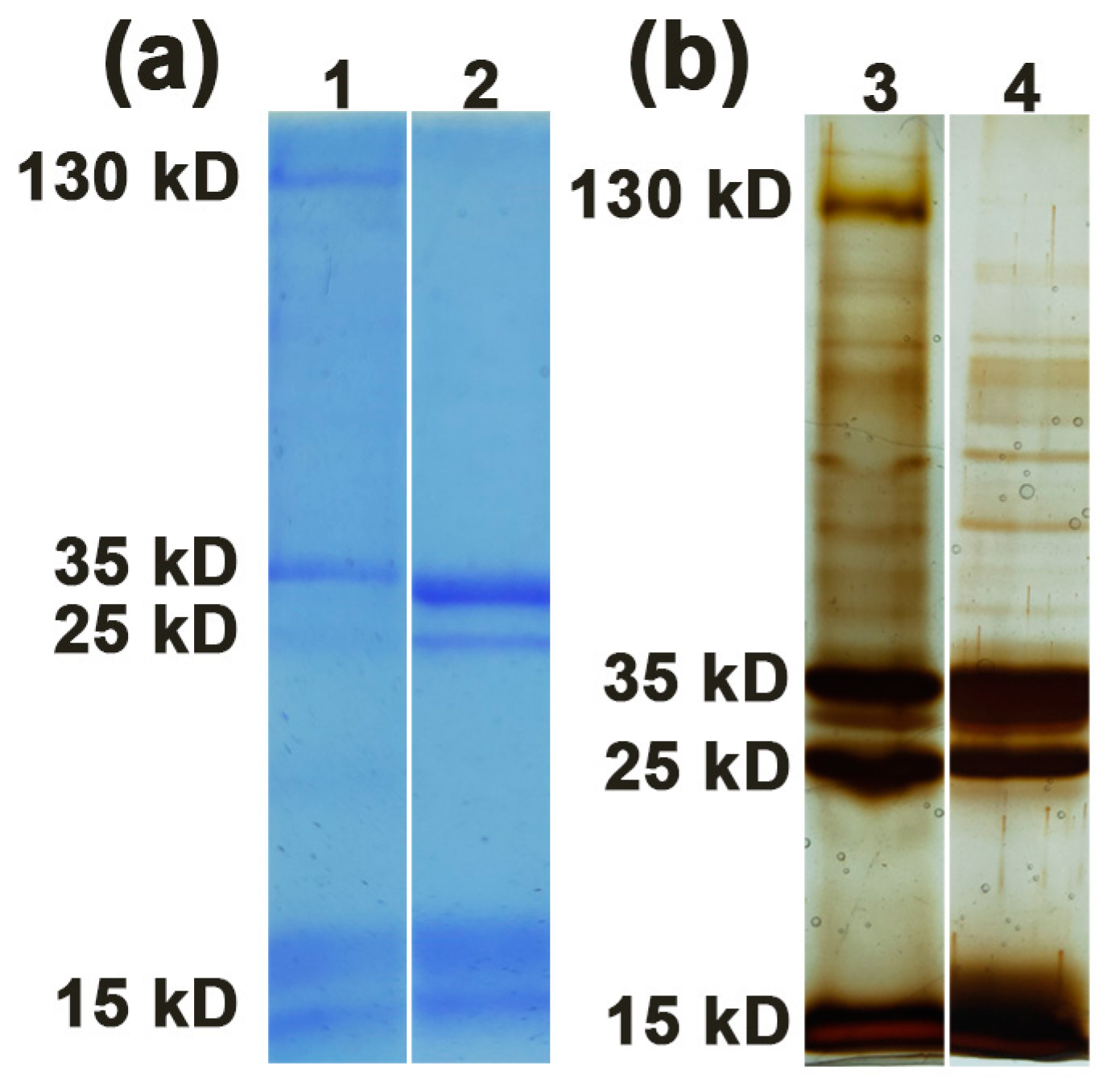
2.1. Protein Profile of PHA-L
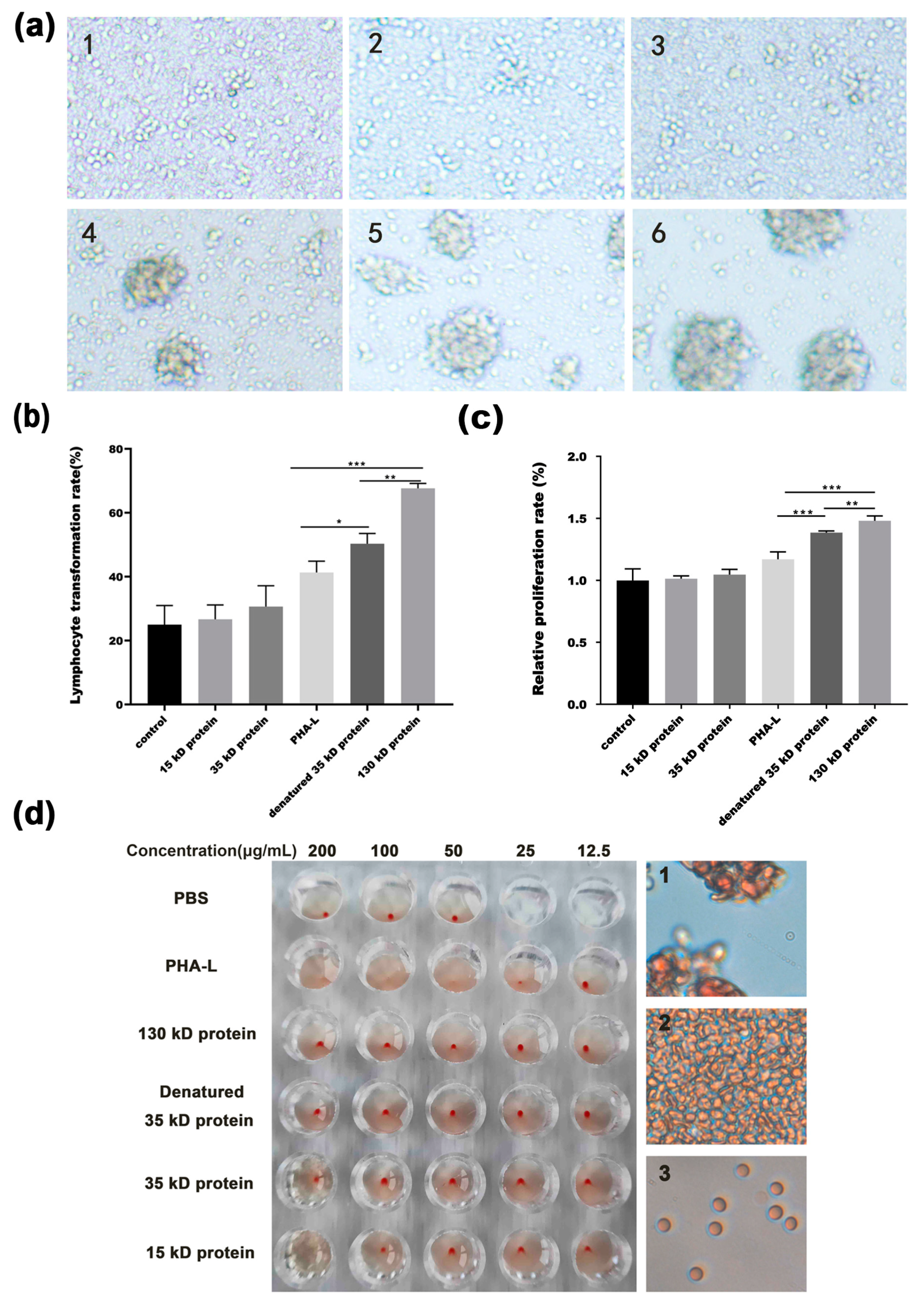
2.2. Characterization of Functional Protein Bands from PHA-L
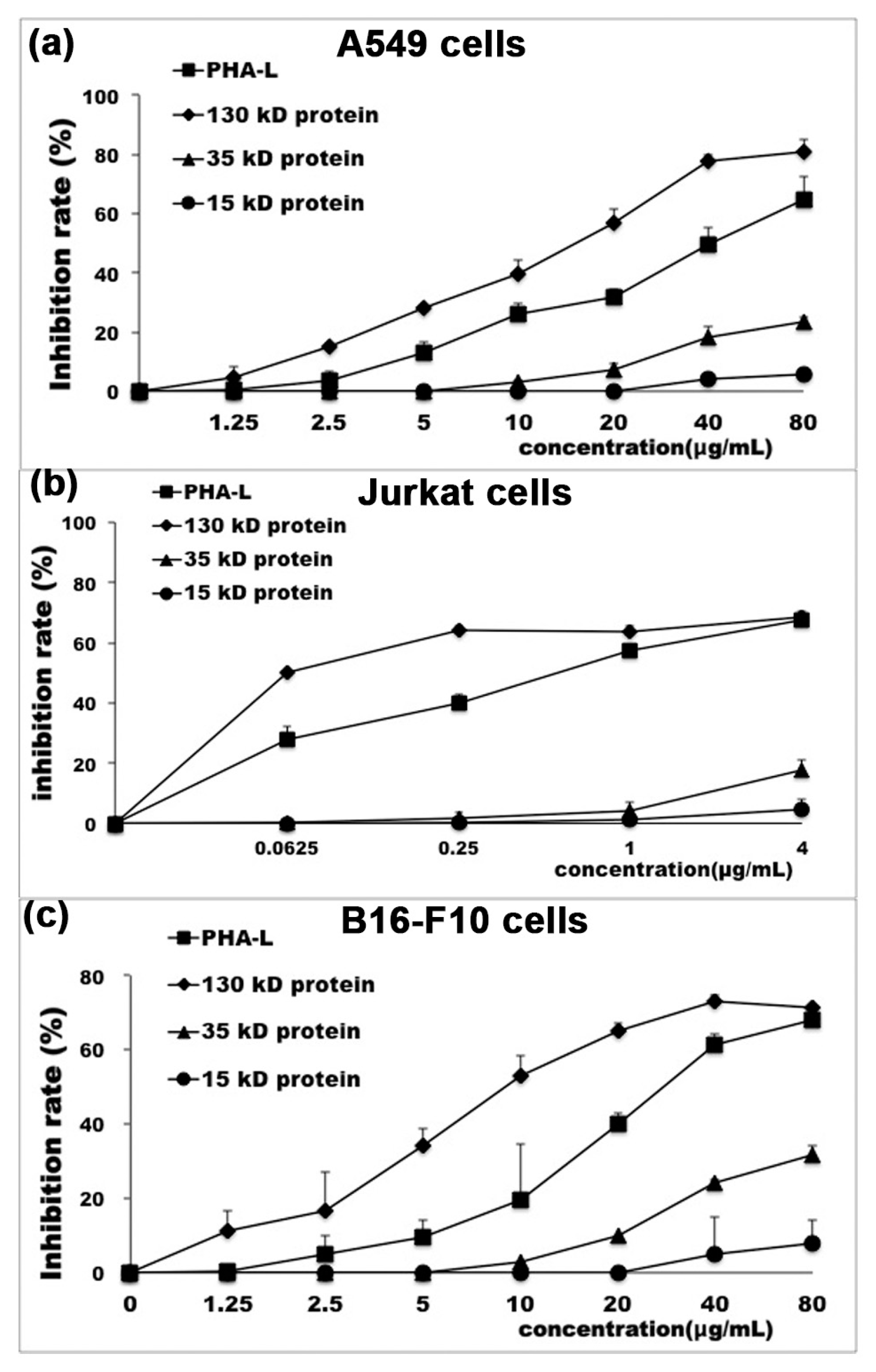
2.3. In Vitro Anti-Tumor Effect of Functional Protein from PHA-L
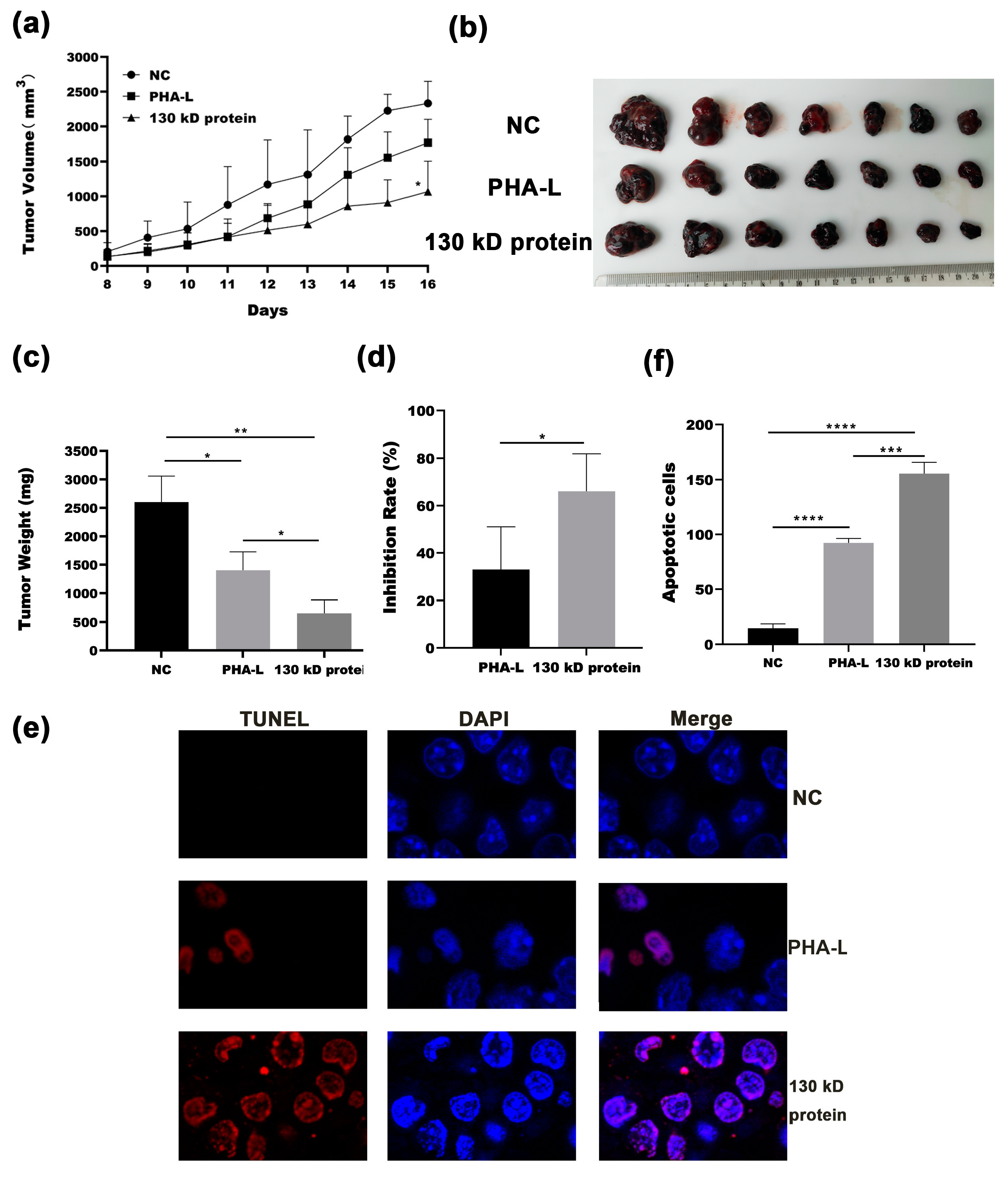
2.4. In Vivo Anti-Tumor Effect of 130 kD Protein from PHA-L
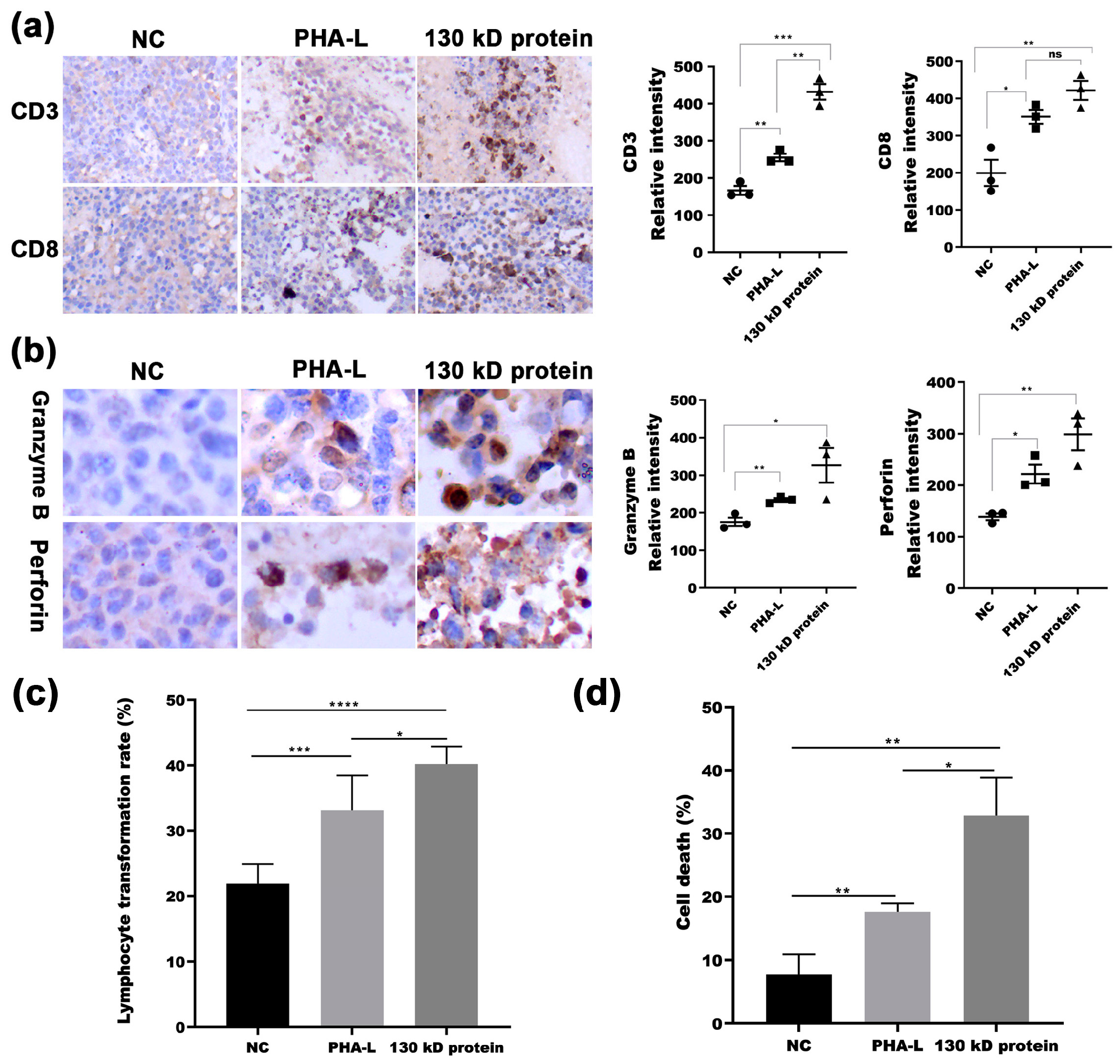
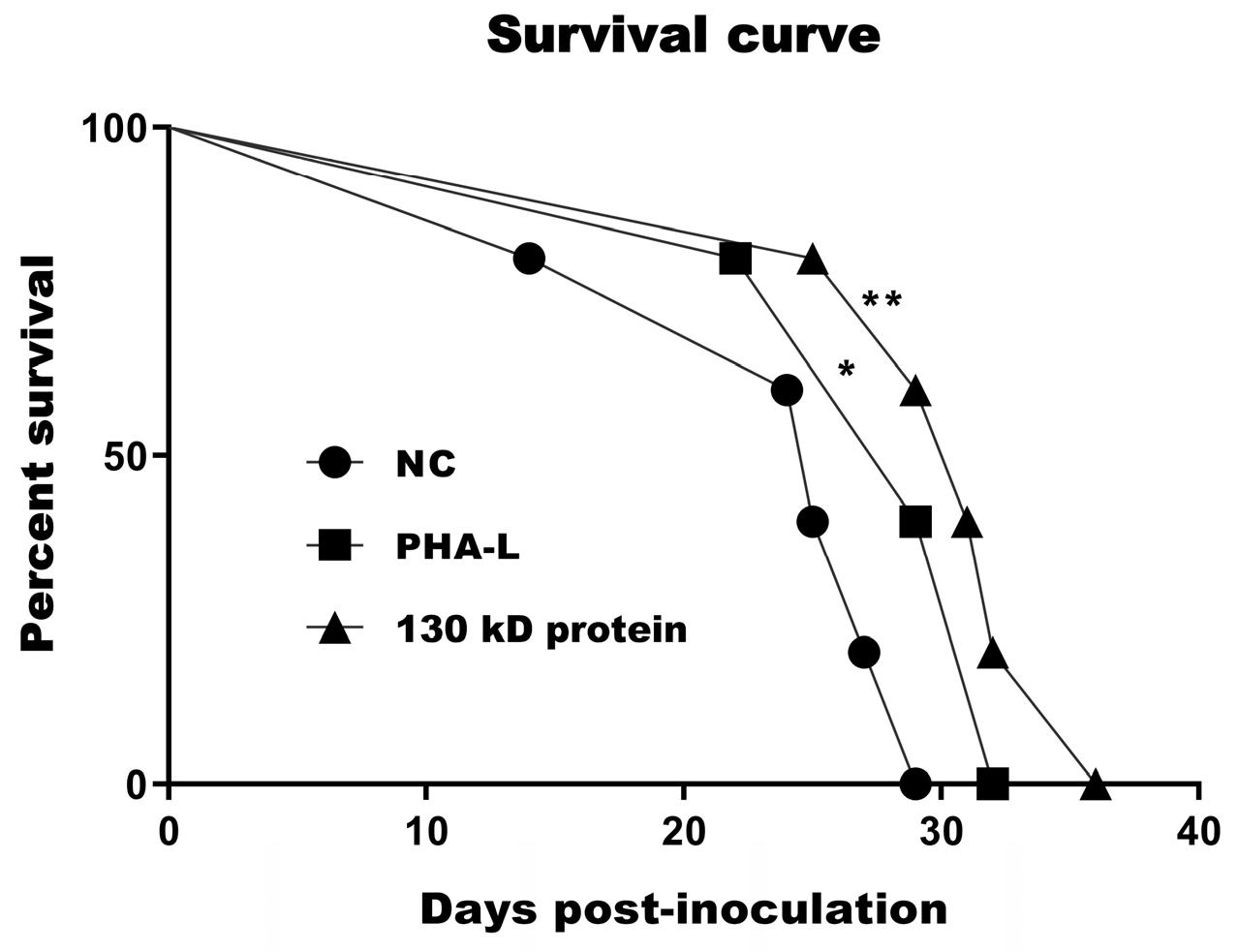
2.5. In Vivo Immunoregulatory Activity of 130 kD Protein from PHA-L
3. Discussion
4. Materials and Methods
4.1. Reagents and Cells
4.2. SDS-PAGE and Silver Staining
4.3. Acquisition of Different Band Proteins of PHA-L
4.4. Lymphocyte Proliferation Assay and Wright’s Staining
4.5. Hemagglutination Assay
4.6. Tumor Cell Culture and Cell Viability Assay
4.7. Cell-Mediated Cytotoxicity Assay
4.8. Murine Tumor Models
4.9. TUNEL Assay and IHC
4.10. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Nowell, P.C. Phytohemagglutinin: An Initiator of Mitosis in Cultures of Normal Human Leukocytes. Cancer Res. 1960, 20, 462–466. [Google Scholar]
- Nathan, S.; Halina, L. History of lectins: From hemagglutinins to biological recognition molecules. Glycobiology 2004, 14, 53R–62R. [Google Scholar]
- Lis, H.; Sharon, N.; Ngadi, M.O.; Ma, Y. Lectins: Carbohydrate-Specific Proteins that Mediate Cellular Recognition. Chem. Rev. 1998, 98, 637–674. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Simpson, B.K.; Ngadi, M.O.; Ma, Y. In vitro studies of the digestibility of lectin from black turtle bean (Phaseolus vulgaris). Food Chem. 2015, 173, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Damme, E.J.M.V.; Peumans, W.J.; Barre, A.; Rougé, P. Plant Lectins: A Composite of Several Distinct Families of Structurally and Evolutionary Related Proteins with Diverse Biological Roles. Crit. Rev. Plant Sci. 1998, 17, 575–692. [Google Scholar] [CrossRef]
- Feng, W.; Jing, Y.; Zhou, Y.; Luo, Y.; Wu, S.; Huang, M.; Yin, B.; Huang, J.; Mao, L.; Sun, Z. The Use of TB-Specific Antigen/Phytohemagglutinin Ratio for Diagnosis and Treatment Monitoring of Extrapulmonary Tuberculosis. Front. Immunol. 2018, 9, 1047. [Google Scholar]
- Loris, R.; Hamelryck, T.; Bouckaert, J.; Wyns, L. Legume lectin structure. Biochim. Biophys. Acta 1998, 1383, 9–36. [Google Scholar] [CrossRef]
- Franz, H.; Ziska, P.; Mohr, J. Lectins—Definition and Classification. Acta Histochem. 1982, 71, 19–21. [Google Scholar] [CrossRef]
- Gupta, G.; Surolia, A.; Sampathkumar, S.-G. Lectin microarrays for glycomic analysis. Omics 2010, 14, 419–436. [Google Scholar] [CrossRef] [Green Version]
- Li, C.Y.; Meng, L.; Liu, B.; Bao, J.-K. Galanthus nivalis Agglutinin (GNA)-Related Lectins: Traditional Proteins, Burgeoning Drugs? Curr. Chem. Biol. 2009, 3, 324–333. [Google Scholar] [CrossRef]
- Kumar, S.; Verma, A.K.; Das, M.; Jain, S.; Dwivedi, P.D. Clinical complications of kidney bean (Phaseolus vulgaris L.) consumption. Nutrition 2013, 29, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Gatehouse, A.M.R.; Dewey, F.M.; Dove, J.; Fenton, K.A.; Pusztai, A. Effect of seed lectins from Phaseolus vulgaris on the development of larvae of Callosobruchus maculatus mechanism of toxicity. J. Sci. Food Agric. 2010, 35, 373–380. [Google Scholar] [CrossRef]
- Antonyuk, V.O. Lectins and Their Stock Sources; Kvart: Lavive, Ukraine, 2005; p. 554. [Google Scholar]
- Bardocz, S.; Grant, G.; Pusztai, A.; Franklin, M.F.; Carvalho, A.D.F.U. The effect of phytohaemagglutinin at different dietary concentrations on the growth, body composition and plasma insulin of the rat. Brit. J. Nutr. 1996, 76, 613–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Verma, A.K.; Sharma, A.; Kumar, D.; Tripathi, A.; Chaudhari, B.; Das, M.; Jain, S.; Dwivedi, P.D. Phytohemagglutinins augment red kidney bean (Phaseolus vulgaris L.) induced allergic manifestations. J. Proteom. 2013, 93, 50–64. [Google Scholar] [CrossRef]
- Grant, G.; Greer, F.; McKenzie, N.H.; Pusztai, A. Nutritional response of mature rats to kidney bean (Phaseolus vulgaris) lectins. J. Sci. Food Agric. 1985, 36, 409–414. [Google Scholar] [CrossRef]
- Vasconcelos, I.M.; José Tadeu, A.O. Antinutritional properties of plant lectins. Toxicon 2004, 44, 385–403. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, A.; Das, M.; Jain, S.K.; Dwivedi, P.D. Leucoagglutinating phytohemagglutinin: Purification, characterization, proteolytic digestion and assessment for allergenicity potential in BALB/c mice. Immunopharmacol. Immunotoxicol. 2014, 36, 138–144. [Google Scholar] [CrossRef]
- Kasera, R.; Singh, A.B.; Lavasa, S.; Nagendra, K.; Arora, N. Purification and Immunobiochemical Characterization of a 31 kDa Cross-Reactive Allergen from Phaseolus vulgaris (Kidney Bean). PLoS ONE 2013, 8, e63063. [Google Scholar] [CrossRef]
- Ang, A.S.W.; Cheung, R.C.F.; Dan, X.; Chan, Y.S.; Pan, W.; Ng, T. Purification and Characterization of a Glucosamine-Binding Antifungal Lectin from Phaseolus vulgaris cv. Chinese Pinto Beans with Antiproliferative Activity Towards Nasopharyngeal Carcinoma Cells. Appl. Biochem. Biotechnol. 2014, 172, 672–686. [Google Scholar] [CrossRef]
- Hou, Y.; Hou, Y.; Yanyan, L.; Qin, G.; Li, J. Extraction and Purification of a Lectin from Red Kidney Bean and Preliminary Immune Function Studies of the Lectin and Four Chinese Herbal Polysaccharides. J. Biomed. Biotechnol. 2010, 217342, 1110–7243. [Google Scholar] [CrossRef]
- Kamemura, K.; Furuichi, Y.; Umekawa, H.; Takahashi, T. Purification and characterization of novel lectins from Great Northern bean, Phaseolus vulgaris L. Biochim. Biophys. Acta 1993, 1158, 181–188. [Google Scholar] [CrossRef]
- Goldstein, I.J.; Hughes, R.C.; Monsigny, M.; Osawa, T.; Sharon, N. What should be called a lectin? Nature 1980, 285, 66. [Google Scholar] [CrossRef]
- Sharon, N.; Lis, H. Lectins as cell recognition molecules. Science 1989, 246, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N. Lectins: Carbohydrate-specific reagents and biological recognition molecules. J. Biol. Chem. 2007, 282, 2753–2764. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Simpson, B.K.; Sun, H.; Ngadi, M.O.; Ma, Y.; Huang, T. Phaseolus vulgaris lectins: A systematic review of characteristics and health implications. Crit. Rev. Food Sci. 2015, 58, 70–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mejía, E.G.; Prisecaru, V.I. Lectins as Bioactive Plant Proteins: A Potential in Cancer Treatment. Crit. Rev. Food Sci. 2005, 45, 425–445. [Google Scholar] [CrossRef]
- Fang, E.F.; Pan, W.L. A new Phaseolus vulgaris lectin induces selective toxicity on human liver carcinoma HepG2 cells. Arch. Toxicol. 2011, 85, 1551–1563. [Google Scholar] [CrossRef]
- Kuo, W.T.; Ho, Y.J.; Kuo, S.M.; Lin, F.H.; Tsai, F.J.; Chen, Y.S.; Dong, G.C.; Yao, C.H. Induction of the Mitochondria Apoptosis Pathway by Phytohemagglutinin Erythroagglutinating in Human Lung Cancer Cells. Ann. Surg. Oncol. 2011, 18, 848–856. [Google Scholar] [CrossRef]
- Lam, S.K.; Ng, T.B. Apoptosis of human breast cancer cells induced by hemagglutinin from Phaseolus vulgaris cv. Legumi secchi. Food Chem. 2011, 126, 595–602. [Google Scholar] [CrossRef]
- Pryme, I.F.; Pusztai, A.; Bardocz, S.; Ewen, S.W. The induction of gut hyperplasia by phytohaemagglutinin in the diet and limitation of tumour growth. Histol. Histopathol. 1998, 13, 575–583. [Google Scholar]
- Pryme, I.F.; Bardocz, S.; Pusztai, A.; Ewen, S.W. The growth of an established murine non-Hodgkin lymphoma tumour is limited by switching to a phytohaemagglutinin-containing diet. Cancer Lett. 1999, 146, 87–91. [Google Scholar] [CrossRef]
- Kochubei, T.O.; Maksymchuk, O.V.; Piven, O.; Lukash, L.L. Isolectins of phytohemagglutinin are able to induce apoptosis in HEp-2 carcinoma cells in vitro. Exp. Oncol. 2015, 37, 116–119. [Google Scholar] [CrossRef]
- Ng, T.B.; Pan, W.; Fei Fang, E.; Pan, W.; Bun Ng, T. A Hemagglutinin from Northeast Red Beans with Immunomodulatory Activity and Anti-proliferative and Apoptosis-inducing Activities toward Tumor Cells. Proteinn Pept. Lett. 2013, 20, 1159–1169. [Google Scholar]
- Hiebert, P.R.; Granville, D.J. Granzyme B in injury, inflammation, and repair. Trends Mol. Med. 2012, 18, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Lostao, L.; Anel, A.; Pardo, J. How Do Cytotoxic Lymphocytes Kill Cancer Cells? Clin. Cancer Res. 2015, 21, 5047–5056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordy, C.; He, Y.W. Endocytosis by target cells: An essential means for perforin- and granzyme-mediated killing. Cell. Mol. Immunol. 2012, 9, 5–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousalova, I.; Krepela, E. Granzyme B-induced apoptosis in cancer cells and its regulation. Int. J. Oncol. 2010, 37, 1361–1378. [Google Scholar]
- Chang, C.P.; Yang, M.C.; Liu, H.S.; Lin, Y.S.; Lei, H.Y. Concanavalin A induces autophagy in hepatoma cells and has a therapeutic effect in a murine in situ hepatoma model. Hepatology 2007, 45, 286–296. [Google Scholar] [CrossRef]
- Augustin, M.; Bock, P.R.; Hanisch, J.; Karasmann, M.; Schneider, B. Safety and efficacy of the long-term adjuvant treatment of primary intermediate to high-risk malignant melanoma (UICC/AJCC stage II and III) with standardized fermented European mistletoe (Viscum album L.) extract. Results from a multicenter, comparative, epidemiological cohort study in Germany and Switzerland. Arzneimittelforschung 2005, 55, 38–49. [Google Scholar]
- Mabed, M.; El-Helw, L.; Shamaa, S. Phase II study of viscum fraxini-2 in patients with advanced hepatocellular carcinoma. Br. J. Cancer 2004, 90, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, S.; Koser, M.L.; Cyr, W.A.; Chopda, G.R.; Tao, J.; Shui, X.; Ying, B.; Chen, D.; Pandya, P.; Chipumuro, E.; et al. Direct Pharmacological Inhibition of β-Catenin by RNA Interference in Tumors of Diverse Origin. Mol. Cancer Ther. 2016, 15, 2143–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, P.; Leng, X.; Duan, J.; Zhu, Y.; Wang, J.; Yan, Z.; Min, S.; Wei, D.; Wang, X. Functional Component Isolated from Phaseolus vulgaris Lectin Exerts In Vitro and In Vivo Anti-Tumor Activity through Potentiation of Apoptosis and Immunomodulation. Molecules 2021, 26, 498. https://doi.org/10.3390/molecules26020498
Wang P, Leng X, Duan J, Zhu Y, Wang J, Yan Z, Min S, Wei D, Wang X. Functional Component Isolated from Phaseolus vulgaris Lectin Exerts In Vitro and In Vivo Anti-Tumor Activity through Potentiation of Apoptosis and Immunomodulation. Molecules. 2021; 26(2):498. https://doi.org/10.3390/molecules26020498
Chicago/Turabian StyleWang, Peipei, Xiaohong Leng, Jiaqi Duan, Yue Zhu, Jue Wang, Zirui Yan, Shitong Min, Dapeng Wei, and Xia Wang. 2021. "Functional Component Isolated from Phaseolus vulgaris Lectin Exerts In Vitro and In Vivo Anti-Tumor Activity through Potentiation of Apoptosis and Immunomodulation" Molecules 26, no. 2: 498. https://doi.org/10.3390/molecules26020498
APA StyleWang, P., Leng, X., Duan, J., Zhu, Y., Wang, J., Yan, Z., Min, S., Wei, D., & Wang, X. (2021). Functional Component Isolated from Phaseolus vulgaris Lectin Exerts In Vitro and In Vivo Anti-Tumor Activity through Potentiation of Apoptosis and Immunomodulation. Molecules, 26(2), 498. https://doi.org/10.3390/molecules26020498