
Nano Milk Protein-Mucilage Complexes: Characterization and Anticancer Effect

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
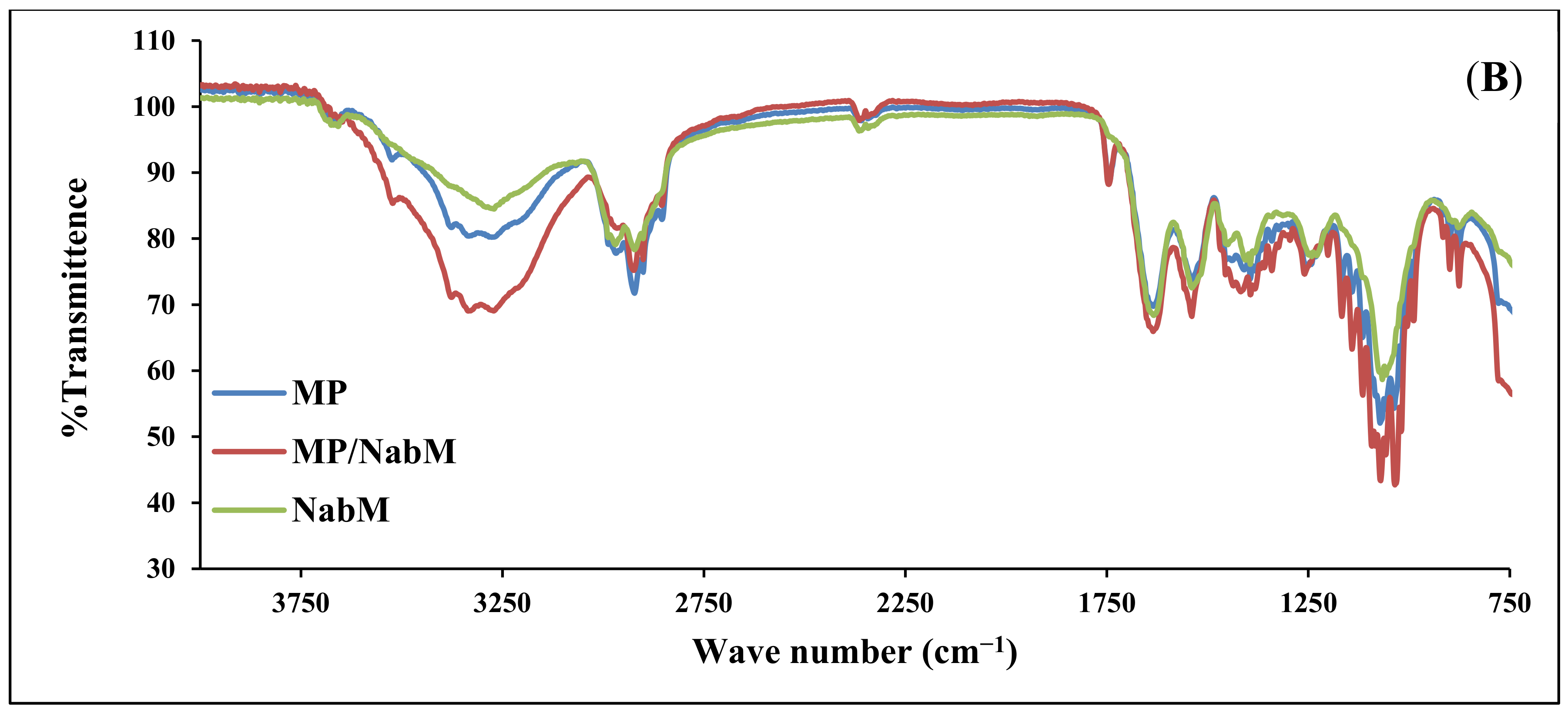
2.1. Fourier-Transform Infrared (FTIR) Spectroscopy
2.2. Physicochemical and Functional Properties of MP and MPMC
2.3. Bioactive Components
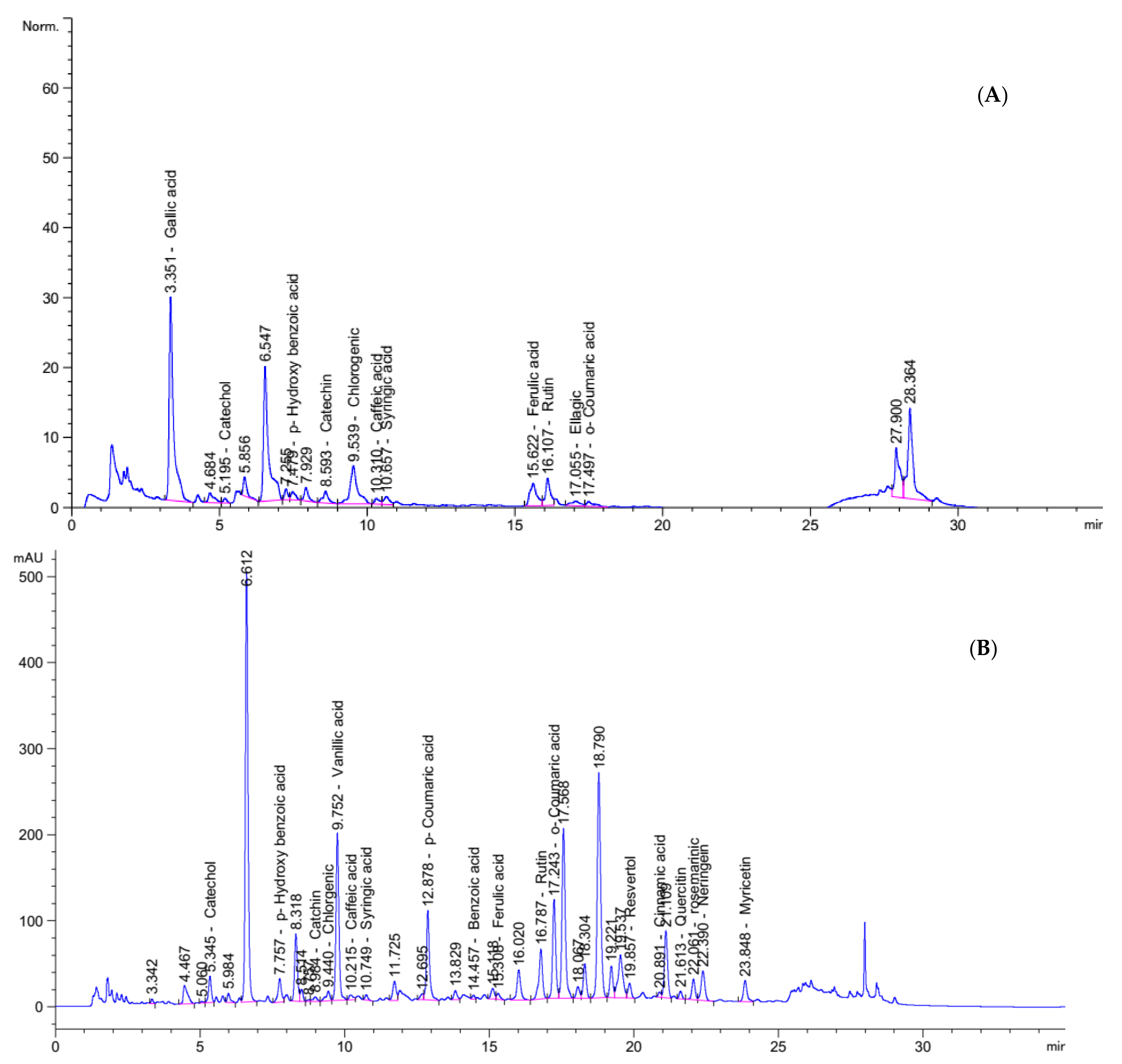
2.3.1. Phenolic Compounds of PHM and NabM
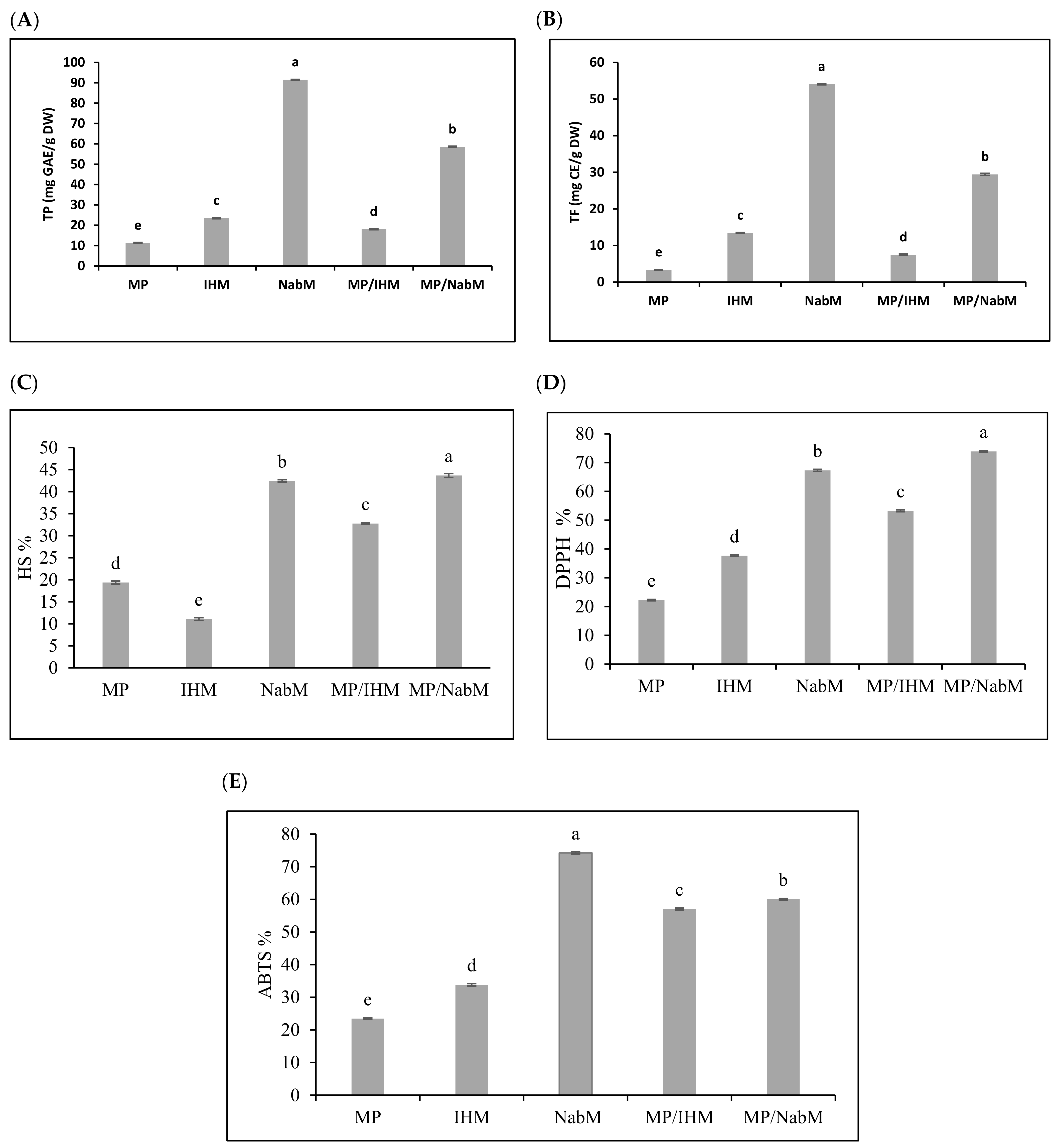
2.3.2. Total Phenolic, Flavonoid, and Antioxidant Activity of MP and MPMC
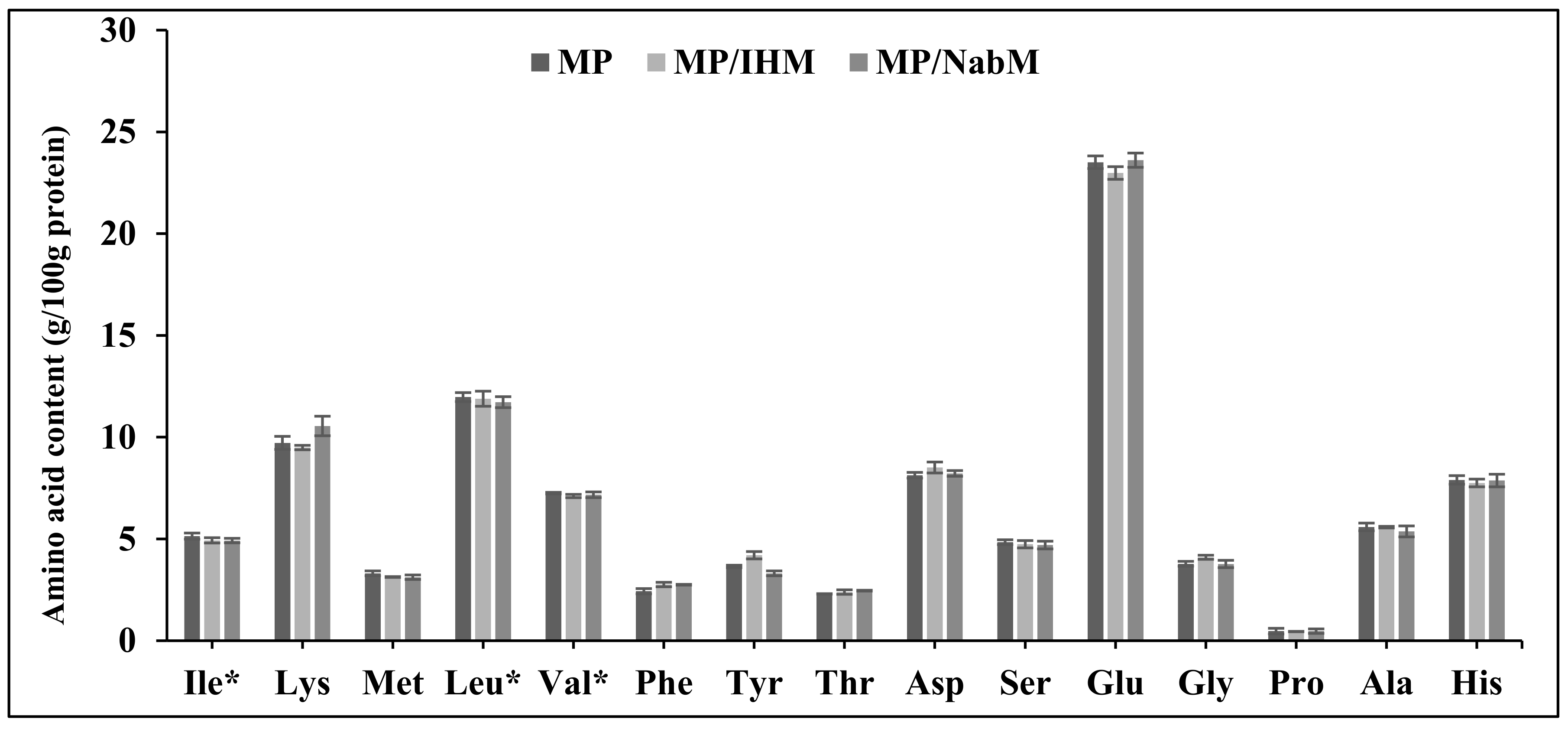
2.4. Amino Acid Profile of MP and MPMC
2.5. Rheological Properties of MP and MPMCs
2.5.1. Apparent Viscosity
2.5.2. Flow Behavior
2.6. Differential Scanning Calorimetry of MP and MPMC
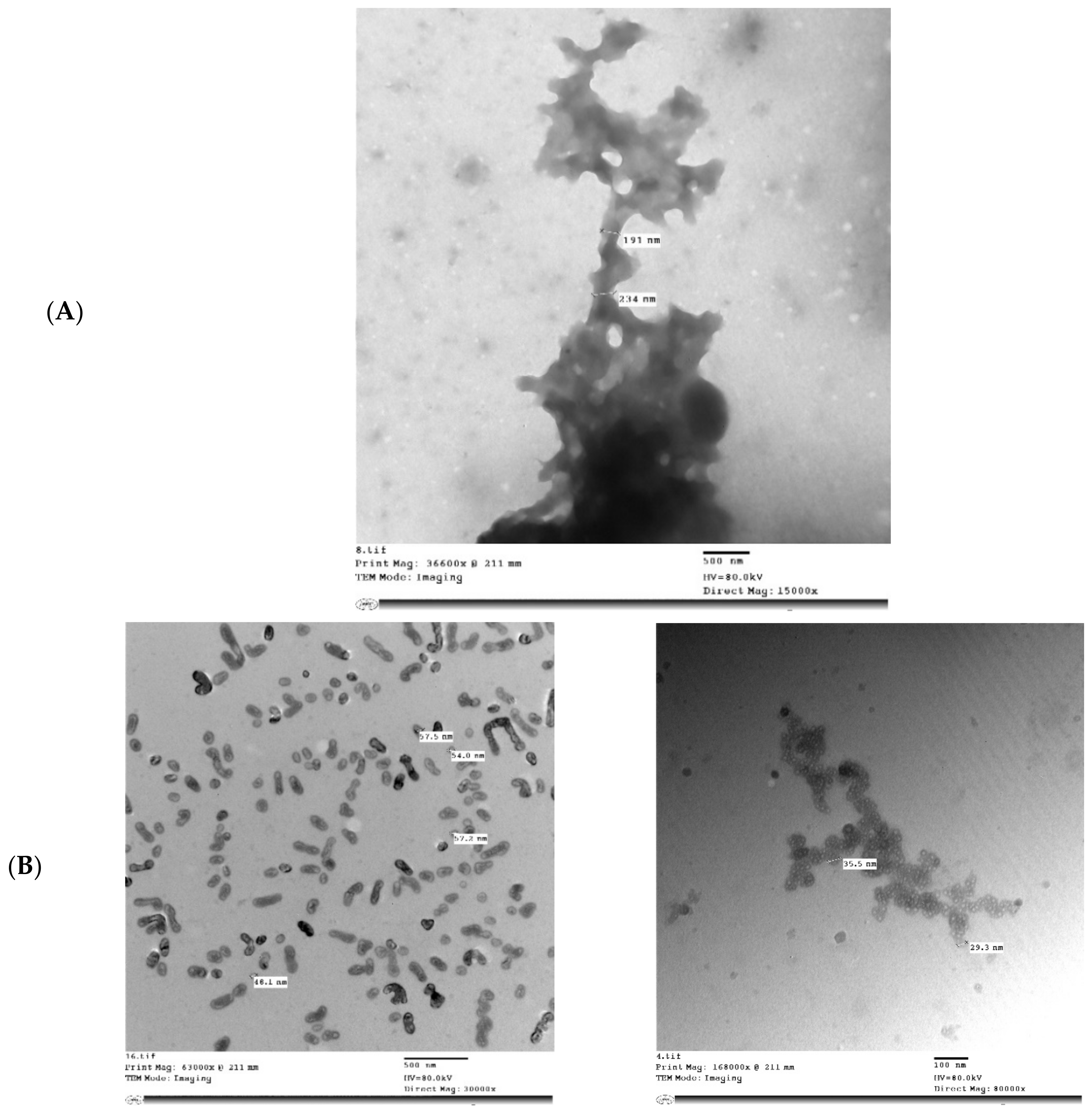
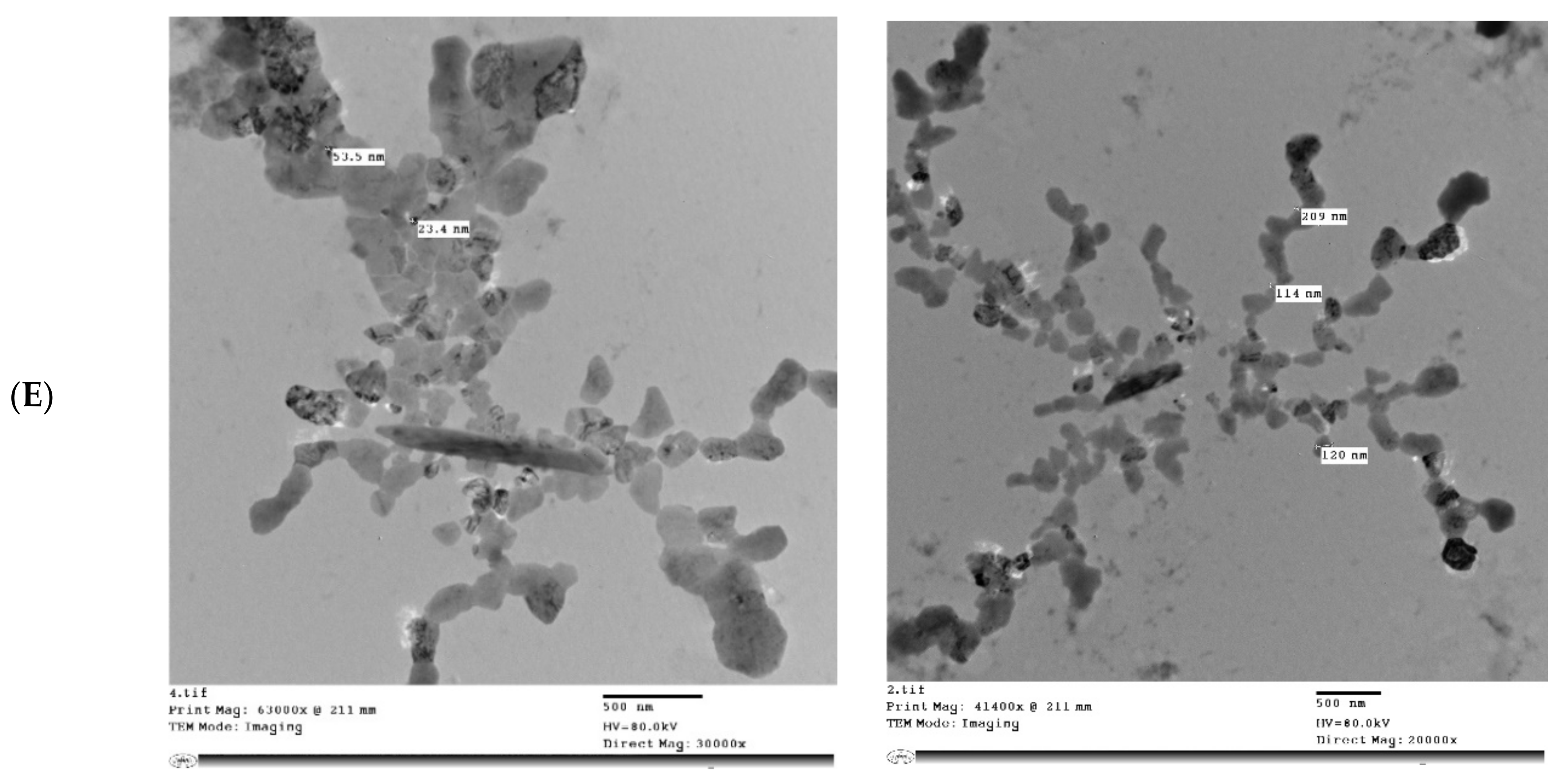
2.7. Transmission Electron Microscopy (TEM) of MP and MPMC
2.8. Biological Studies on MP and MPMC
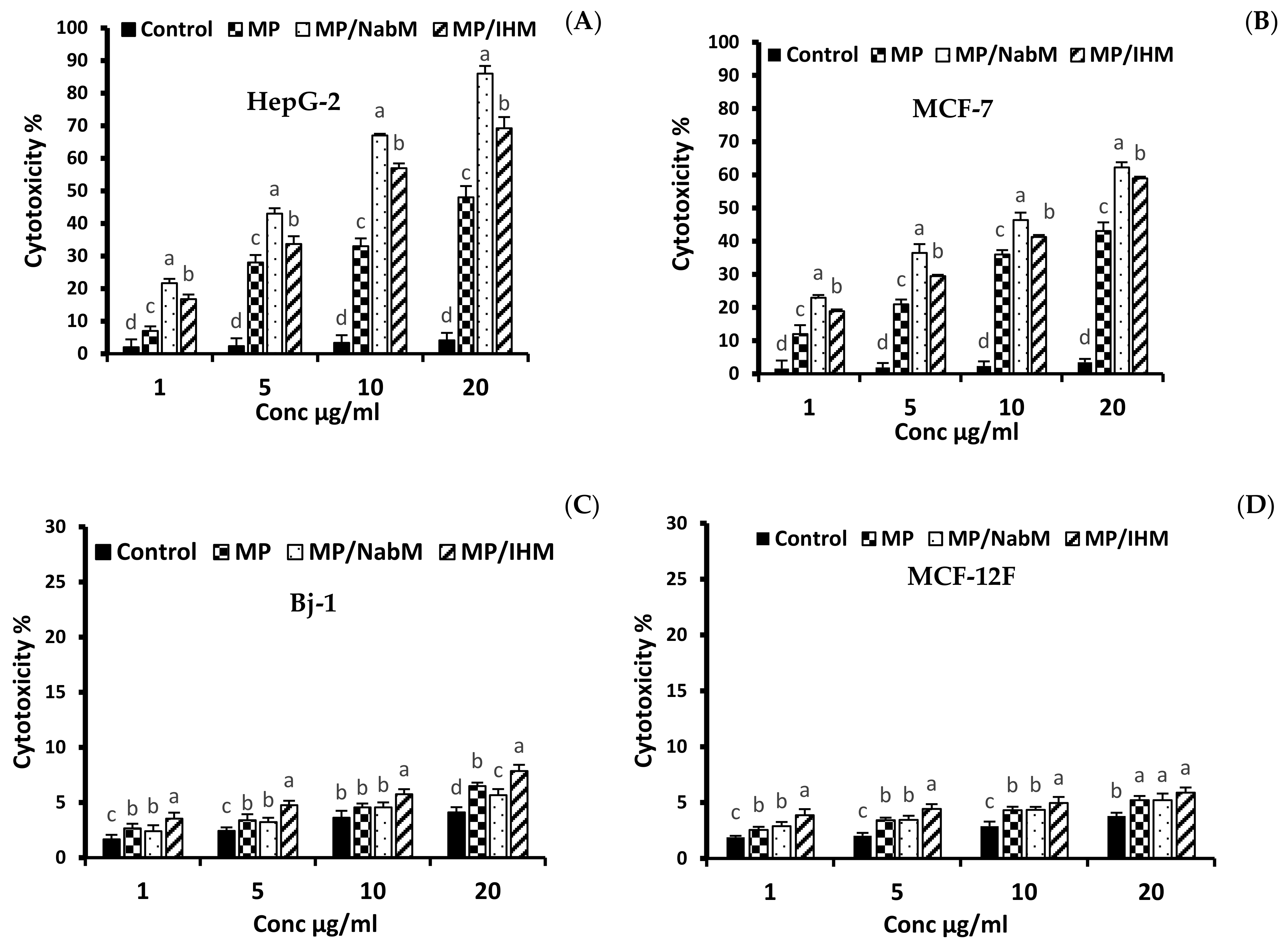
2.8.1. Anticancer Activity of MP and MPMC
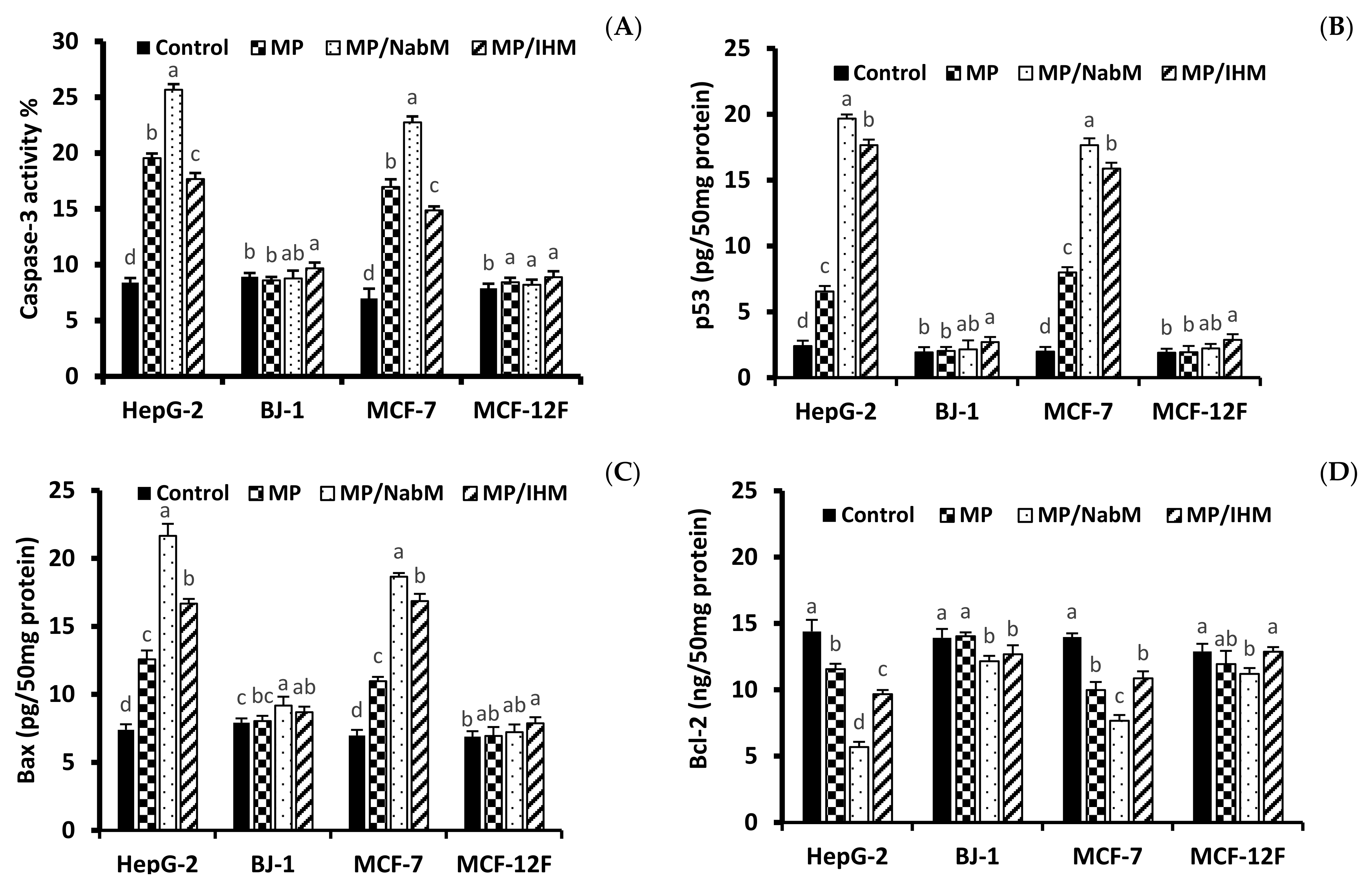
2.8.2. Determination of p53, Bax, Caspase-3, and Bcl-2 Proteins Level
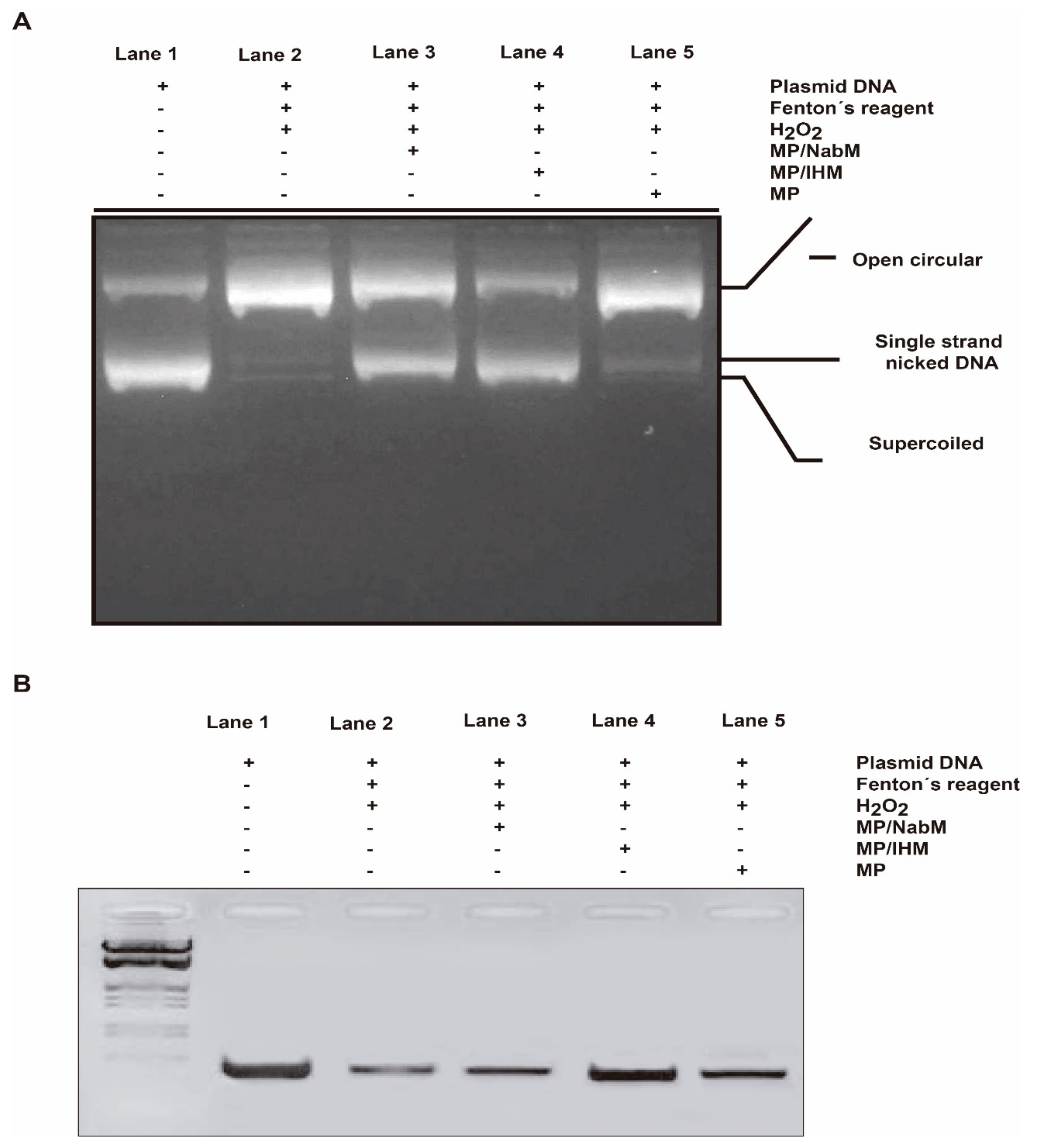
2.8.3. DNA Damage Protection by MPMC against Oxidative Cleavage of RNH1 Plasmid
2.8.4. DNA Damage Protection by MPMC against Oxidative Cleavage of Genomic DNA
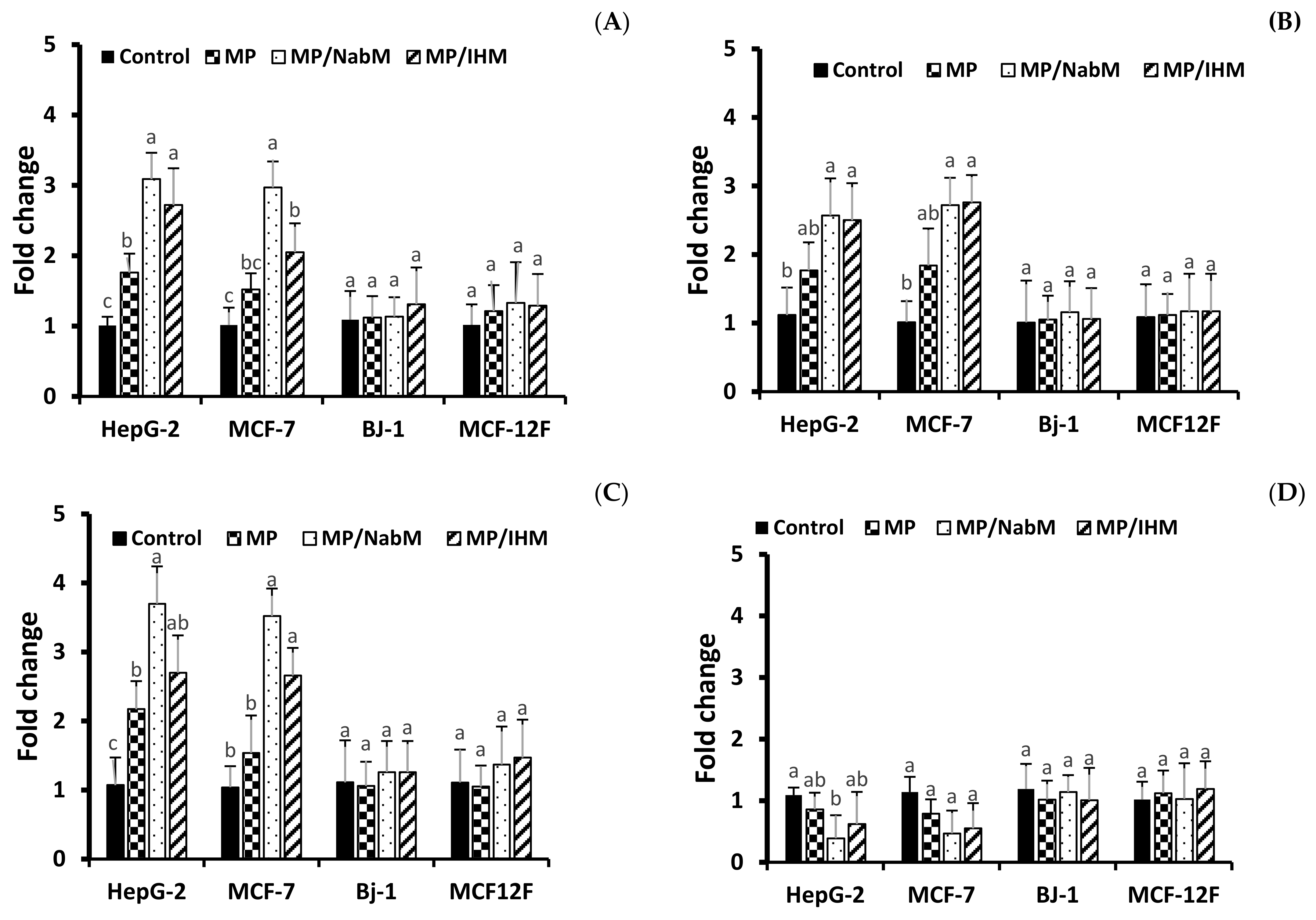
2.8.5. Gene Expression
3. Materials and Methods
3.1. Materials
3.2. Preparation of Isabgol Husk Mucilage (IHM) and Nabeq Mucilage (NabM)
3.3. Preparation of Milk Protein-Mucilage Complexes (MPMC)
3.4. Physicochemical and Functional Characteristics
3.4.1. pH Measurement
3.4.2. Determination of Bulk Density (BD), Tapped Density (TD), and Carr’s Index
3.4.3. Water Absorption Index (WAI) and Water Solubility Index (WSI)
3.4.4. Milk Protein Digestibility
3.5. Phenolic Fractions of PHM and NabM
3.6. Total Phenolic and Flavonoid Content of MP and MPMC
3.7. Antioxidant Activity of MP and MPMC
3.8. Amino Acids Profile of MP and MPMC
3.9. Rheological Properties
3.10. Differential Scanning Calorimetry (DSC)
3.11. Fourier-Transform Infrared Spectroscopy (FTIR)
3.12. Transmission Electron Microscopy
3.13. Biological Studies on MP and MPMC
3.13.1. In Vitro Anticancer Activity
3.13.2. Determination of p53, Bax, Caspase-3, and Bcl-2 Proteins Level
3.13.3. DNA Damage Induced by Oxidative Stress Protection
3.13.4. PCR-Based Genomic DNA Damage Assay
3.13.5. Gene Expression Profiling by RT-qPCR
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GLOBOCAN 2020: New Global Cancer Data. Available online: https://www.uicc.org/news/globocan-2020-new-global-cancer-data (accessed on 15 February 2021).
- Yu, Y.; Shen, M.; Song, Q.; Xie, J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohydr. Polym. 2018, 183, 91–101. [Google Scholar] [CrossRef]
- Elshaghabee, F.M.; El-Maksoud, A.A.; Alharbi, S.A.; Alfarraj, S.; Mohamed, M.S. Fortification of Acidophilus-bifidus-thermophilus (ABT) Fermented Milk with Heat-Treated Industrial Yeast Enhances Its Selected Properties. Molecules 2021, 26, 3876. [Google Scholar] [CrossRef]
- Xie, J.H.; Tang, W.; Jin, M.L.; Li, J.E.; Xie, M.Y. Recent advances in bioactive polysaccharides from Lycium barbarum L., Zizyphus jujuba Mill, Plantago Spp., and Morus Spp.: Structures and functionalities. Food Hydrocoll. 2016, 60, 148–160. [Google Scholar] [CrossRef]
- El-Maksoud, A.A.A.; Korany, R.M.; El-Ghany, I.H.A.; El-Beltagi, H.S.; Ambrósio, F.; de Gouveia, G.M. Dietary solutions to dyslipidemia: Milk protein–polysaccharide conjugates as liver biochemical enhancers. J. Food Biochem. 2020, 44, e13142. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Bhattacharya, A. Isabgol husk: A herbal remedy for human health. J. Pharm. Res. 2009, 2, 296–301. [Google Scholar]
- Hussein, A.S. Ziziphus spina-christi: Analysis of Bioactivities and Chemical Composition. In Wild Fruits: Composition, Nutritional Value and Products; Springer: Cham, Switzerland, 2019; pp. 175–197. [Google Scholar]
- Barras, D.; Widmann, C. Promises of Apoptosis-Inducing Peptides in Cancer Therapeutics. Curr. Pharm. Biotechnol. 2011, 12, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, D.; Sakakibara, H.; Fukuno, H.; Kimura, K.; Hori, A.; Nara, T.; Kato, K.; Sakono, M. Whey protein-hydrolyzed peptides diminish hepatic lipid levels in rats consuming high-sucrose diets. Food Sci. Technol. Res. 2016, 22, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Maksoud, A.A.; Abd El-Ghany, I.H.; El-Beltagi, H.S.; Anankanbil, S.; Banerjee, C.; Petersen, S.V.; Pérez, B.; Guo, Z. Adding functionality to milk-based protein: Preparation, and physico-chemical characterization of β-lactoglobulin-phenolic conjugates. Food Chem. 2018, 241, 281–289. [Google Scholar] [CrossRef]
- Sah, B.N.P.; Vasiljevic, T.; McKechnie, S.; Donkor, O.N. Identification of anticancer peptides from bovine milk proteins and their potential roles in management of cancer: A critical review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 123–138. [Google Scholar] [CrossRef]
- Pepe, G.; Tenore, G.C.; Mastrocinque, R.; Stusio, P.; Campiglia, P. Potential anticarcinogenic peptides from bovine milk. J. Amino Acids 2013, 2013, 1–7. [Google Scholar] [CrossRef]
- Ausar, S.F.; Bianco, I.D.; Badini, R.G.; Castagna, L.F.; Modesti, N.M.; Landa, C.A.; Beltramo, D.M. Characterization of casein micelle precipitation by chitosans. J. Dairy Sci. 2001, 84, 361–369. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Bandyopadhyay, P. Polysaccharide-protein interactions and their relevance in food colloids. Complex World Polysacch. 2012, 14, 395–406. [Google Scholar]
- Højgaard, C.; Kofoed, C.; Espersen, R.; Johansson, K.E.; Villa, M.; Willemoës, M.; Lindorff-Larsen, K.; Teilum, K.; Winther, J.R. A soluble, folded protein without charged amino acid residues. Biochemistry 2016, 55, 3949–3956. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Sun, S.Q.; Yu, L.; Xu, C.H.; Noda, I.; Zhang, X.R. Sequential changes of main components in different kinds of milk powders using two-dimensional infrared correlation analysis. J. Mol. Struct. 2006, 799, 77–84. [Google Scholar] [CrossRef]
- Vukic, V.R.; Hrnjez, D.V.; Kanuric, K.G.; Milanovic, S.D.; Iličic, M.D.; Torbica, A.M.; Tomic, J.M. The effect of kombucha starter culture on the gelation process, microstructure and rheological properties during milk fermentation. J. Texture Stud. 2014, 45, 261–273. [Google Scholar] [CrossRef]
- Farahnaky, A.; Askari, H.; Majzoobi, M.; Mesbahi, G. The impact of concentration, temperature and pH on dynamic rheology of psyllium gels. J. Food Eng. 2010, 100, 294–301. [Google Scholar] [CrossRef]
- Ganguly, S.; Mondal, S.; Das, P.; Bhawal, P.; Maity, P.P.; Ghosh, S.; Dhara, S.; Das, N.C. Design of psyllium-g-poly(acrylic acid-co-sodium acrylate)/cloisite 10A semi-IPN nanocomposite hydrogel and its mechanical, rheological and controlled drug release behaviour. Int. J. Biol. Macromol. 2018, 111, 983–998. [Google Scholar] [CrossRef] [PubMed]
- Qaisrani, T.B.; Qaisrani, M.M.; Qaisrani, T.M. Arabinoxylans from psyllium husk: A review. J. Environ. Agric. Sci. 2016, 6, 33–39. [Google Scholar]
- Müllertz, A.; Ogbonna, A.; Ren, S.; Rades, T. New perspectives on lipid and surfactant based drug delivery systems for oral delivery of poorly soluble drugs. J. Pharm. Pharmacol. 2010, 62, 1622–1636. [Google Scholar] [CrossRef]
- Phillips, G.O.; Williams, P.A. Handbook of Hydrocolloids; Woodhead Publishing Limited: Cambridge, UK, 2009; pp. 10–90. [Google Scholar]
- Singh, V.; Guizani, N.; Essa, M.M.; Rahman, M.S.; Selvaraju, S. In vitro antioxidant activities of Ziziphus spina-christi fruits (red date) grown in Oman. Biotechnology 2012, 11, 209–216. [Google Scholar] [CrossRef]
- Li, X.; Lin, J.; Gao, Y.; Han, W.; Chen, D. Antioxidant activity and mechanism of Rhizoma Cimicifugae. Chem. Centr. J. 2012, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Koşar, M. Relationship between Structure of Phenolics and Anticancer Activity. Proceedings 2017, 1, 978. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Ray, S.; Nagarajan, K. Glutamic acid as anticancer agent: An overview. Saudi Pharm. J. 2013, 21, 337–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.H.; Chen, W.Y. Rheological properties of the water-soluble mucilage of a green laver, Monostroma nitidium. J. Appl. Phycol. 2001, 13, 481–488. [Google Scholar] [CrossRef]
- Kale, M.S.; Yadav, M.P.; Hanah, K.A. Suppression of psyllium husk suspension viscosity by addition of water soluble polysaccharides. J. Food Sci. 2016, 81, E2476–E2483. [Google Scholar] [CrossRef]
- Thanatcha, R.; Pranee, A.J.I.F.R.J. Extraction and characterization of mucilage in Ziziphus mauritiana Lam. Int. Food Res. J. 2011, 18, 201–212. [Google Scholar]
- Ali, H.; Alli, I.; Ismail, A.; Kermasha, S. Protein-phenolic interactions in food. Eurasian J. Anal. Chem. 2012, 7, 123–133. [Google Scholar]
- Krešic, G.; Lelas, V.; Herceg, Z.; Režek, A. Effects of high pressure on functionality of whey protein concentrate and whey protein isolate. Lait 2006, 86, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Ye, A. Complexation between milk proteins and polysaccharides via electrostatic interaction: Principles and applications—A review. Int. J. Food Sci. Technol. 2008, 43, 406–415. [Google Scholar] [CrossRef]
- Goh, K.K.; Teo, A.; Sarkar, A.; Singh, H. Milk protein-polysaccharide interactions. In Milk Proteins; Academic Press: Cambridge, MA, USA, 2020; pp. 499–535. [Google Scholar]
- Jeong, J.E.; Hong, Y.H. Anticancer activity of bovine α-lactalbumin treated with microbial transglutaminase. Int. J. Dairy Technol. 2012, 65, 212–216. [Google Scholar] [CrossRef]
- Yarramala, D.S.; Prakash, P.; Ranade, D.S.; Doshi, S.; Kulkarni, P.P.; Bhaumik, P.; Rao, C.P. Cytotoxicity of apo bovine α-lactalbumin complexed with La 3+ on cancer cells supported by its high resolution crystal structure. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lima, C.F.; Rodrigues, L.R. Invitro evaluation of bovine lactoferrin potential as an anticancer agent. Int. Dairy J. 2015, 40, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Leischner, C.; Egert, S.; Burkard, M.; Venturelli, S. Potential Protective Protein Components of Cow’s Milk against Certain Tumor Entities. Nutrients 2021, 13, 1974. [Google Scholar] [CrossRef]
- Elzoghby, A.O.; Helmy, M.W.; Samy, W.M.; Elgindy, N.A. Novel ionically crosslinked casein nanoparticles for flutamide delivery: Formulation, characterization, and in vivo pharmacokinetics. Int. J. Nanomed. 2013, 8, 1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimamura, M.; Yamamoto, Y.; Ashino, H.; Oikawa, T.; Hazato, T.; Tsuda, H.; Iigo, M. Bovine lactoferrin inhibits tumor-induced angiogenesis. Int. J. Cancer 2004, 111, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochem. Biophys. Acta-Biomembr. 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sienkiewicz-Szłapka, E.; Jarmołowska, B.; Krawczuk, S.; Kostyra, E.; Kostyra, H.; Bielikowicz, K. Transport of bovine milk-derived opioid peptides across a Caco-2 monolayer. Int. Dairy J. 2009, 10, 252–257. [Google Scholar] [CrossRef]
- Farmani, F.; Moein, M.; Amanzadeh, A.; Kandelous, H.M.; Ehsanpour, Z.; Salimi, M. Antiproliferative evaluation and apoptosis induction in MCF-7 cells by Ziziphus spina christi leaf extracts. Asian Pac. J. Cancer Prev. 2016, 17, 315–321. [Google Scholar] [CrossRef] [Green Version]
- Abo-Salem, H.M.; Nassrallah, A.; Soliman, A.A.; Ebied, M.S.; Elawady, M.E.; Abdelhamid, S.A.; El-Sawy, E.R.; Al-Sheikh, Y.A.; Aboul-Soud, M.A. Synthesis and Bioactivity Assessment of Novel Spiro Pyrazole-Oxindole Congeners Exhibiting Potent and Selective in vitro Anticancer Effects. Molecules 2020, 25, 1124. [Google Scholar] [CrossRef] [Green Version]
- Ashour, A.E.; Ahmed, A.F.; Kumar, A.; Zoheir, K.M.; Aboul-Soud, M.A.; Ahmad, S.F.; Attia, S.M.; Abd-Allah, A.R.A.; Cheryan, V.T.; Rishi, A.K. Thymoquinone inhibits growth of human medulloblastoma cells by inducing oxidative stress and caspase-dependent apoptosis while suppressing NF-κB signaling and IL-8 expression. Mol. Cell. Biochem. 2016, 416, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Morr, C.V. Functionality of Heated Milk Proteins in Dairy and Related Foods. J. Dairy Sci. 1985, 68, 2773–2781. [Google Scholar] [CrossRef]
- Deshpande, H.W.; Poshadri, A. Physical and sensory characteristics of extruded snacks prepared from Foxtail millet based composite flours. Int. Food Res. J. 2011, 18, 751–756. [Google Scholar]
- Yaǧci, S.; Göǧüş, F. Response surface methodology for evaluation of physical and functional properties of extruded snack foods developed from food-by-products. J. Food Eng. 2008, 86, 122–132. [Google Scholar] [CrossRef]
- Abd El-Ghany, I.H.; El-Asser, M.A.; Nagy, K.S.; Abd El-Maksoud, A.A. Effect of milk proteins on physical and chemical characteristics of crispy puff snacks. J. Agric. Sci. Technol. A 2013, 3, 633–645. [Google Scholar]
- Elsayed, N.; El-Din, H.S.; Altemimi, A.B.; Ahmed, H.Y.; Pratap-Singh, A.; Abedelmaksoud, T.G. In Vitro Antimicrobial, Antioxidant and Anticancer Activities of Egyptian Citrus Beebread. Molecules 2021, 26, 2433. [Google Scholar] [CrossRef]
- Peksa, A.; Kita, A.; Jariene, E.; Danilcenko, H.; Gryszkin, A.; Figiel, A.; Kulaitiene, J.; Cerniauskiene, J.; Aniolowska, M. Amino acid improving and physical qualities of extruded corn snacks using flours made from Jerusalem artichoke (Helianthus tuberosus), amaranth (Amaranthus cruentus L.) and pumpkin (Cucurbita maxima L.). J. Food Qual. 2016, 39, 580–589. [Google Scholar] [CrossRef]
- Barnes, H.A. An examination of the use of rotational viscometers for the quality control of non-Newtonian liquid products in factories. Appl. Rheol. 2001, 11, 89–101. [Google Scholar] [CrossRef]
- Sinaki, N.Y.; Scanlon, M.G. Shear Flow Behavior of Aqueous Suspensions of Potato Parenchyma Powder. Food Biophys. 2018, 13, 432–439. [Google Scholar] [CrossRef]
- Abd El-Maksoud, A.A.; Anankanbil, S.; Zhou, Y.; Abd El-Ghany, I.H.; El-Beltagi, H.S.; Banerjee, C.; Petersen, S.V.; Guo, Z. Grafting phenolics onto milk protein via conjugated polymerization for delivery of multiple functionalities: Synthesis and characterization. Food Chem. 2019, 301, 125298. [Google Scholar] [CrossRef]
- Gallagher, J.R.; Kim, A.J.; Gulati, N.M.; Harris, A.K. Negative-Stain Transmission Electron Microscopy of Molecular Complexes for Image Analysis by 2D Class Averaging. Curr. Prot. Microbiol. 2019, 54, e90. [Google Scholar] [CrossRef] [Green Version]
- Repetto, G.; Del Peso, A.; Zurita, J.L. Neutral Red Uptake Assay for the Estimation of Cell Viability/Cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hirao, A.; Kong, Y.Y.; Matsuoka, S.; Wakeham, A.; Ruland, J.; Yoshida, H.; Liu, D.; Elledge, S.J.; Mak, T.W. DNA damage-induced activation of p53 by the checkpoint kinase Chk2. Science 2000, 287, 1824–1827. [Google Scholar] [CrossRef] [PubMed]
- Aboul-Soud, M.A.M.; Ashour, A.E.; Challis, J.K.; Ahmed, A.F.; Kumar, A.; Nassrallah, A.; Alahmari, T.A.; Saquib, Q.; Siddiqui, M.A.; Al-Sheikh, Y.; et al. Biochemical and Molecular Investigation of in Vitro Antioxidant and Anticancer Activity Spectrum of Crude Extracts of Willow Leaves Salix safsaf. Plants 2020, 9, 1295. [Google Scholar] [CrossRef] [PubMed]
- El-Hallouty, S.M.; Soliman, A.A.; Nassrallah, A.; Salamatullah, A.; Alkaltham, M.S.; Kamal, K.Y.; Hanafy, E.A.; Gaballa, H.S.; Aboul-Soud, M.A. Crude Methanol Extract of Rosin Gum Exhibits Specific Cytotoxicity against Human Breast Cancer Cells via Apoptosis Induction. Curr. Med. Chem. Anti-Cancer Agents 2020, 20, 1028–1036. [Google Scholar] [CrossRef]
- Leba, L.J.; Brunschwig, C.; Saout, M.; Martial, K.; Vulcain, E.; Bereau, D.; Robinson, J.C. Optimization of a DNA nicking assay to evaluate Oenocarpus bataua and Camellia sinensis antioxidant capacity. Int. J. Mol. Sci. 2014, 15, 18023–18039. [Google Scholar] [CrossRef] [Green Version]
- Sadat Asadi, N.; Heidari, M.M.; Khatami, M. Protective effect of Berberis vulgaris on Fenton reaction-induced DNA cleavage. Avicenna J. Phytomed. 2019, 9, 213–220. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IHM | NabM | MP | MP/IHM | MP/NabM | |
|---|---|---|---|---|---|
| pH | 7.79 ± 0.21 a | 6.42 ± 0.61 b | 4.7 ± 0.21 d | 5.1 ± 0.1 c | 4.6 ± 0.22 d |
| Bulk density (g/cm3) | 0.34 ± 0.04 c | 1.25 ± 0.07 a | 1.18 ± 0.05 a | 0.55 ± 0.01 b | 1.23 ± 0.01 a |
| True density (g/cm3) | 0.53 ± 0.02 e | 1.42 ± 0.04 a | 1.25 ± 0.03 c | 0.91 ± 0.01 d | 1.29 ± 0.01 b |
| Carr’s Index (%) | 57.58 ± 0.14 a | 53.97 ± 0.11 b | 30.6 ± 0.10 e | 39.56 ± 0.12 c | 33.65 ± 0.18 d |
| WSI (%) | 71.35 ± 1.52 a | 33.52 ± 0.24 c | 15.87 ± 1.04 e | 36.67 ± 0.16 b | 21.42 ± 0.13 d |
| WAI (g/g) | 21.42 ± 0.12 a | 8.13 ± 0.15 c | 3.54 ± 0.04 e | 16.63 ± 0.53 b | 5.50 ± 0.29 d |
| Protein digestibility (%) | 79.44 ± 0.5 a | 81.63 ± 1.22 a | 75.63 ± 1.22 b |
| Samples | MP | MP/IHM | MP/NabM |
|---|---|---|---|
| Denaturation onset (°C) | 50.01 | 138.12 | 62.89 |
| Protein denaturation (°C) | 101.44 | 148.24 | 121.4 |
| Degradation degree (°C) | 302.3 | - | - |
| ΔH (J/g) | 98.63 | 553.08 | 121.82 |
| Cell Lines | IC50 µg/mL | |||
|---|---|---|---|---|
| MP | MP/NabM | MP/IHM | DOX | |
| HepG2 | 18.2 ± 1.86 a | 5.13 ± 0.31 b | 8.89 ± 0.15 b | 14.87 ± 0.19 c |
| MCF-7 | 22.5 ± 2.75 a | 10.07 ± 0.72 c | 12.3 ± 0.018 b | 0.74 ± 1.18 b |
| MCF7-12F | 198.4 ± 6.23 c | 256.47 ± 7.25 a | 215.32 ± 4.34 b | 24.16 ± 7.23 b |
| Bj-1 | 203.34 ± 2.45 c | 238.56 ± 3.54 a | 236.67 ± 4.78 b | 36.23 ± 5.64 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Maksoud, A.A.A.; Makhlouf, A.I.A.; Altemimi, A.B.; El-Ghany, I.H.A.; Nassrallah, A.; Cacciola, F.; Abedelmaksoud, T.G. Nano Milk Protein-Mucilage Complexes: Characterization and Anticancer Effect. Molecules 2021, 26, 6372. https://doi.org/10.3390/molecules26216372
El-Maksoud AAA, Makhlouf AIA, Altemimi AB, El-Ghany IHA, Nassrallah A, Cacciola F, Abedelmaksoud TG. Nano Milk Protein-Mucilage Complexes: Characterization and Anticancer Effect. Molecules. 2021; 26(21):6372. https://doi.org/10.3390/molecules26216372
Chicago/Turabian StyleEl-Maksoud, Ahmed Ali Abd, Amal I. A. Makhlouf, Ammar B. Altemimi, Ismail H. Abd El-Ghany, Amr Nassrallah, Francesco Cacciola, and Tarek Gamal Abedelmaksoud. 2021. "Nano Milk Protein-Mucilage Complexes: Characterization and Anticancer Effect" Molecules 26, no. 21: 6372. https://doi.org/10.3390/molecules26216372
APA StyleEl-Maksoud, A. A. A., Makhlouf, A. I. A., Altemimi, A. B., El-Ghany, I. H. A., Nassrallah, A., Cacciola, F., & Abedelmaksoud, T. G. (2021). Nano Milk Protein-Mucilage Complexes: Characterization and Anticancer Effect. Molecules, 26(21), 6372. https://doi.org/10.3390/molecules26216372