
Naturally Available Flavonoid Aglycones as Potential Antiviral Drug Candidates against SARS-CoV-2

 ,
, 
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of the Isolated Compounds
2.2. Docking Studies
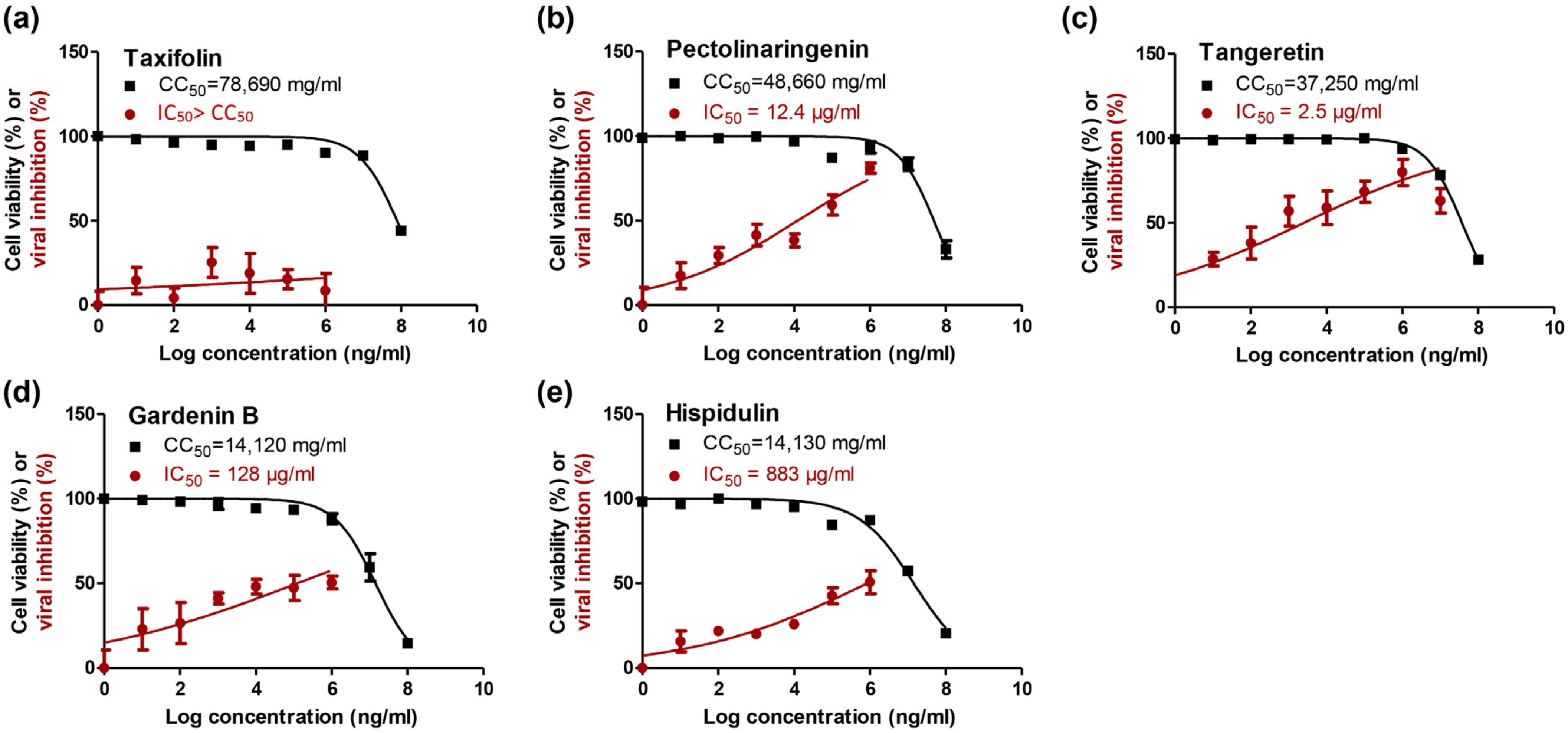
2.3. In Vitro Validation
3. Material and Methods
3.1. Plant Material
3.2. Extraction, Isolation and Structure Elucidation
3.2.1. Taxifolin (Dihydroquercetin) (1)
3.2.2. Pectolinarigenin (Scutellarein 4′,6-Dimethyl Ether) (2)
3.2.3. Tangeretin (4′,5,6,7,8-Pentamethoxyflavone) (3)
3.2.4. Gardenin B = Demethyltangeretin (5-Hydroxy 6,7,8,4′-Tetra Methoxy Flavone) (4)
3.2.5. Hispidulin (5)
3.3. Molecular Docking Study
3.3.1. Preparation of the Isolated and Identified Five Flavonoids (1–5)
3.3.2. Target Mpro of SARS-CoV-2 Preparation
3.3.3. Docking of the Database Compounds (1–5) to the Dimer Mpro of SARS-CoV-2
3.4. In Vitro Anti-SARS-CoV-2 Activity
3.4.1. Cytotoxicity (CC50) Determination
3.4.2. Inhibitory Concentration 50 (IC50) Determination
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kamps, B.S.; Christia, H. COVID Reference; Steinhauser Verlag: Wuppertal, Germany, 2021; Available online: https://amedeo.com/CovidReference06.pdf (accessed on 10 June 2021).
- Elebeedy, D. Anti-SARS-CoV-2 activities of tanshinone IIA, carnosic acid, rosmarinic acid, salvianolic acid, baicalein, and glycyrrhetinic acid between computational and in vitro insights. RSC Adv. 2021, 11, 29267–29286. [Google Scholar] [CrossRef]
- Kandeil, A. Immunogenicity and Safety of an Inactivated SARS-CoV-2 Vaccine: Preclinical Studies. Vaccines 2021, 9, 214. [Google Scholar] [CrossRef] [PubMed]
- Shehata, M.M. In Silico and In Vivo Evaluation of SARS-CoV-2 Predicted Epitopes-Based Candidate Vaccine. Molecules 2021, 26, 6182. [Google Scholar] [CrossRef]
- El Gizawy, H.A. Pimenta dioica (L.) Merr. Bioactive Constituents Exert Anti-SARS-CoV-2 and Anti-Inflammatory Activities: Molecular Docking and Dynamics, In Vitro, and In Vivo Studies. Molecules 2021, 26, 5844. [Google Scholar] [CrossRef]
- El-Demerdash, A. Investigating the structure–activity relationship of marine natural polyketides as promising SARS-CoV-2 main protease inhibitors. RSC Adv. 2021, 11, 31339–31363. [Google Scholar] [CrossRef]
- Mahmoud, D.B. Delineating a potent antiviral activity of Cuphea ignea extract loaded nano-formulation against SARS-CoV-2: In silico and in vitro studies. J. Drug Deliv. Sci. Technol. 2021, 66, 102845. [Google Scholar] [CrossRef]
- Roviello, V.; Roviello, G.N. Less COVID-19 deaths in southern and insular Italy explained by forest bathing, Mediterranean environment, and antiviral plant volatile organic compounds. Environ. Chem. Lett. 2021, 1–11. [Google Scholar] [CrossRef]
- Vicidomini, C.; Roviello, V.; Roviello, G.N. In Silico Investigation on the Interaction of Chiral Phytochemicals from Opuntia ficus-indica with SARS-CoV-2 Mpro. Symmetry 2021, 13, 1041. [Google Scholar] [CrossRef]
- Muhammad, I. Screening of potent phytochemical inhibitors against SARS-CoV-2 protease and its two Asian mutants. Comput. Biol. Med. 2021, 133, 104362. [Google Scholar] [CrossRef]
- Ibrahim, M.A. Blue Biotechnology: Computational Screening of Sarcophyton Cembranoid Diterpenes for SARS-CoV-2 Main Protease Inhibition. Mar. Drugs 2021, 19, 391. [Google Scholar] [CrossRef] [PubMed]
- Oesch, F.; Oesch-Bartlomowicz, B.; Efferth, T. Toxicity as prime selection criterion among SARS-active herbal medications. Phytomedicine 2021, 85, 153476. [Google Scholar] [CrossRef]
- Vicidomini, C.; Roviello, V.; Roviello, G.N. Molecular Basis of the Therapeutical Potential of Clove (Syzygium aromaticum L.) and Clues to Its Anti-COVID-19 Utility. Molecules 2021, 26, 1880. [Google Scholar] [CrossRef] [PubMed]
- Calland, N. Hepatitis C virus and natural compounds: A new antiviral approach? Viruses 2012, 4, 2197–2217. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.-F. Antiviral phenolic compounds from Arundina gramnifolia. J. Nat. Prod. 2013, 76, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Shoala, T. Nanobiotechnological Approaches to Enhance Potato Resistance against Potato Leafroll Virus (PLRV) Using Glycyrrhizic Acid Ammonium Salt and Salicylic Acid Nanoparticles. Horticulturae 2021, 7, 402. [Google Scholar] [CrossRef]
- Ngwa, W. Potential of flavonoid-inspired phytomedicines against COVID-19. Molecules 2020, 25, 2707. [Google Scholar] [CrossRef]
- Jo, S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzym. Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Abo Elmaaty, A. Computational Insights on the Potential of Some NSAIDs for Treating COVID-19: Priority Set and Lead Optimization. Molecules 2021, 26, 3772. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A. Telaprevir is a potential drug for repurposing against SARS-CoV-2: Computational and in vitro studies. Heliyon 2021, 7, e07962. [Google Scholar] [CrossRef] [PubMed]
- Al-Karmalawy, A.A.; Eissa, I.H. Molecular docking and dynamics simulations reveal the potential of anti-HCV drugs to inhibit COVID-19 main protease. Pharm. Sci. 2021, 27. [Google Scholar] [CrossRef]
- Sarhan, A.A.; Ashour, N.A.; Al-Karmalawy, A.A. The journey of antimalarial drugs against SARS-CoV-2: Review article. Inform. Med. Unlocked 2021, 24, 100604. [Google Scholar] [CrossRef]
- Zaki, A.A. Calendulaglycoside A Showing Potential Activity Against SARS-CoV-2 Main Protease: Molecular Docking, Molecular Dynamics, and SAR Studies. J. Tradit. Complement. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Elmaaty, A.A. In a search for potential drug candidates for combating COVID-19: Computational study revealed salvianolic acid B as a potential therapeutic targeting 3CLpro and spike proteins. J. Biomol. Struct. Dyn. 2021, 30, 1–28. [Google Scholar] [CrossRef]
- Soltane, R. Strong Inhibitory Activity and Action Modes of Synthetic Maslinic Acid Derivative on Highly Pathogenic Coronaviruses: COVID-19 Drug Candidate. Pathogens 2021, 10, 623. [Google Scholar] [CrossRef] [PubMed]
- Elmaaty, A.A. Revisiting activity of some glucocorticoids as a potential inhibitor of SARS-CoV-2 main protease: Theoretical study. RSC Adv. 2021, 11, 10027–10042. [Google Scholar] [CrossRef]
- Soltan, M.A. In Silico Prediction of a Multitope Vaccine against Moraxella catarrhalis: Reverse Vaccinology and Immunoinformatics. Vaccines 2021, 9, 669. [Google Scholar] [CrossRef] [PubMed]
- Soltan, M.A. Proteome Based Approach Defines Candidates for Designing a Multitope Vaccine against the Nipah Virus. Int. J. Mol. Sci. 2021, 22, 9330. [Google Scholar] [CrossRef]
- Brogi, S. Computational approaches for drug discovery. Molecules 2019, 24, 3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Karmalawy, A.A.; Khattab, M.J. Molecular modelling of mebendazole polymorphs as a potential colchicine binding site inhibitor. New J. Chem. 2020, 44, 13990–13996. [Google Scholar] [CrossRef]
- Khattab, M.; Al-Karmalawy, A.A. Revisiting Activity of Some Nocodazole Analogues as a Potential Anticancer Drugs Using Molecular Docking and DFT Calculations. Front. Chem. 2021, 9, 92. [Google Scholar] [CrossRef]
- Mahmoud, A. In vitro and in silico characterization of alkaline serine protease from Bacillus subtilis D9 recovered from Saudi Arabia. Heliyon 2021, 7, e08148. [Google Scholar] [CrossRef]
- Gogoi, N. Computational guided identification of a citrus flavonoid as potential inhibitor of SARS-CoV-2 main protease. Mol. Divers. 2021, 25, 1745–1759. [Google Scholar] [CrossRef]
- Da Rocha, M.N. Virtual screening of citrus flavonoid tangeretin: A promising pharmacological tool for the treatment and prevention of Zika fever and COVID-19. J. Comput. Biophys. Chem. 2021, 20, 2150013. [Google Scholar] [CrossRef]
- Omar, S.; Bouziane, I.; Bouslama, Z.; Djemel, A. In-Silico Identification of Potent Inhibitors of COVID-19 Main Protease (Mpro) and Angiotensin Converting Enzyme 2 (ACE2) from Natural Products: Quercetin, Hispidulin, and Cirsimaritin Exhibited Better Potential Inhibition than Hydroxy-Chloroquine Against COVID-19 Main Protease Active Site and ACE2. ChemRxiv 2020. [Google Scholar] [CrossRef]
- Kandeil, A. Bioactive Polyphenolic Compounds Showing Strong Antiviral Activities against Severe Acute Respiratory Syndrome Coronavirus 2. Pathogens 2021, 10, 758. [Google Scholar] [CrossRef] [PubMed]
- Alnajjar, R. Molecular docking, molecular dynamics, and in vitro studies reveal the potential of angiotensin II receptor blockers to inhibit the COVID-19 main protease. Heliyon 2020, 6, e05641. [Google Scholar] [CrossRef]
- Al-Karmalawy, A.A. Molecular Docking and Dynamics Simulation Revealed the Potential Inhibitory Activity of ACEIs Against SARS-CoV-2 Targeting the hACE2 Receptor. Front. Chem. 2021, 9, 227. [Google Scholar]
- Zaki, A.A. Molecular docking reveals the potential of Cleome amblyocarpa isolated compounds to inhibit COVID-19 virus main protease. New J. Chem. 2020, 44, 16752–16758. [Google Scholar] [CrossRef]
- Lee, S. Pectolinarigenin, an aglycone of pectolinarin, has more potent inhibitory activities on melanogenesis than pectolinarin. Biochem. Biophys. Res. Commun. 2017, 493, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Chen, X.; Wu, M. Antidiabetic effect of flavones from Cirsium japonicum DC in diabetic rats. Arch. Pharm. Res. 2010, 33, 353–362. [Google Scholar] [CrossRef]
- Yoo, Y.-M. Pectolinarin and pectolinarigenin of Cirsium setidens prevent the hepatic injury in rats caused by D-galactosamine via an antioxidant mechanism. Biol. Pharm. Bull. 2008, 31, 760–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tundis, R.; Deguin, B.; Loizzo, M.R.; Bonesi, M.; Statti, G.A.; Tillequin, F.; Menichini, F. Potential antitumor agents: Flavones and their derivatives from Linaria reflexa Desf. Bioorg. Med. Chem. Lett. 2005, 15, 4757–4760. [Google Scholar] [CrossRef]
- Cabrera, J. Gardenin B-induced cell death in human leukemia cells involves multiple caspases but is independent of the generation of reactive oxygen species. Chem.-Biol. Interact. 2016, 256, 220–227. [Google Scholar] [CrossRef]
- Parmar, V.S. Highly oxygenated bioactive flavones from Tamarix. Phytochemistry 1994, 36, 507–511. [Google Scholar] [CrossRef]
- Harborne, J.B. The systematic identification of flavonoids: By T.J. Mabry, K.R. Markham and M.B. Thomas, Springer-Verlag, Berlin-Heidelberg-New York, 1970, pp. xii + 354, price DM.98.00, £11.75, $27.00. J. Mol. Struct. 1971, 10, 320. [Google Scholar]
- Yoshikawa, M. Anastatins A and B, new skeletal flavonoids with hepatoprotective activities from the desert plant Anastatica hierochuntica. Bioorg. Med. Chem. Lett. 2003, 13, 1045–1049. [Google Scholar] [CrossRef]
- Farid, M.M. Isoscutellarein 8,4′-dimethyl ether glycosides as cytotoxic agents and chemotaxonomic markers in Kickxia aegyptiaca. Biocatal. Agric. Biotechnol. 2019, 22, 101431. [Google Scholar] [CrossRef]
- Huang, X. Inhibitory mechanisms and interaction of tangeretin, 5-demethyltangeretin, nobiletin, and 5-demethylnobiletin from citrus peels on pancreatic lipase: Kinetics, spectroscopies, and molecular dynamics simulation. Int. J. Biol. Macromol. 2020, 164, 1927–1938. [Google Scholar] [CrossRef] [PubMed]
- Seghiri, R. Phenolic compounds from Centaurea africana. Chem. Nat. Compd. 2006, 42, 610–611. [Google Scholar] [CrossRef]
- Chemical Computing Group Inc. Molecular Operating Environment (MOE), 2016; Chemical Computing Group Inc.: Montreal, QC, Canada, 2021. [Google Scholar]
- Ibrahim, M. Design, synthesis, molecular docking and biological evaluation of some novel quinazoline-4 (3H)-one derivatives as anti-inflammatory agents. Eur. J. Med. Chem. 2012, 46, 185–203. [Google Scholar]
- Ghanem, A. Tanshinone IIA synergistically enhances the antitumor activity of doxorubicin by interfering with the PI3K/AKT/mTOR pathway and inhibition of topoisomerase II: In vitro and molecular docking studies. New J. Chem. 2020, 44, 17374–17381. [Google Scholar] [CrossRef]
- Abdallah, A.E. Design and synthesis of new 4-(2-nitrophenoxy)benzamide derivatives as potential antiviral agents: Molecular modeling and in vitro antiviral screening. New J. Chem. 2021, 45, 16557–16571. [Google Scholar] [CrossRef]
- El-Shershaby, M.H. From triazolophthalazines to triazoloquinazolines: A bioisosterism-guided approach toward the identification of novel PCAF inhibitors with potential anticancer activity. Bioorg. Med. Chem. 2021, 42, 116266. [Google Scholar] [CrossRef] [PubMed]
- Taher, R.F. Two new flavonoids and anticancer activity of Hymenosporum flavum: In vitro and molecular docking studies. J. Herbmed Pharmacol. 2021, 10, 443–458. [Google Scholar]
- Zhang, L. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samra, R.M. Bioassay-guided isolation of a new cytotoxic ceramide from Cyperus rotundus L. S. Afr. J. Bot. 2021, 139, 210–216. [Google Scholar] [CrossRef]
- Al-Karmalawy, A.A.; Elshal, M.F. Concanavalin-A shows synergistic cytotoxicity with tamoxifen via inducing apoptosis in estrogen receptor-positive breast cancer: In vitro and molecular docking studies. Pharm. Sci. 2021. [Google Scholar] [CrossRef]
- Gaber, A.A. Pharmacophore-linked pyrazolo [3,4-d]pyrimidines as EGFR-TK inhibitors: Synthesis, anticancer evaluation, pharmacokinetics, and in silico mechanistic studies. Arch. Pharm. 2021, 10, e2100258. [Google Scholar]
- Ibrahim, M.K.; El-Adl, K.; Al-Karmalawy, A.A. Design, synthesis, molecular docking and anticonvulsant evaluation of novel 6-iodo-2-phenyl-3-substituted-quinazolin-4 (3H)-ones. Bull. Fac. Pharm. Cairo Univ. 2015, 53, 101–116. [Google Scholar] [CrossRef] [Green Version]
- Eliaa, S.G. Empagliflozin and Doxorubicin Synergistically Inhibit the Survival of Triple-Negative Breast Cancer Cells via Interfering with the mTOR Pathway and Inhibition of Calmodulin: In Vitro and Molecular Docking Studies. ACS Pharmacol. Transl. Sci. 2020, 3, 1330–1338. [Google Scholar] [CrossRef]
- Al-Karmalawy, A. Design and Synthesis of New Quinoxaline Derivatives as Potential Histone Deacetylase Inhibitors Targeting Hepatocellular Carcinoma: In Silico, In Vitro, and SAR Studies. Front. Chem. 2021, 9, 725135. [Google Scholar]
- El-Shershaby, M.H. The antimicrobial potential and pharmacokinetic profiles of novel quinoline-based scaffolds: Synthesis and in silico mechanistic studies as dual DNA gyrase and DHFR inhibitors. New J. Chem. 2021, 45, 13986–14004. [Google Scholar] [CrossRef]
- Zaki, A.A. Isolation of cytotoxic active compounds from Reichardia tingitana with investigation of apoptosis mechanistic induction: In silico, in vitro, and SAR studies. S. Afr. J. Bot. 2022, 144, 115–123. [Google Scholar] [CrossRef]
- Khattab, M.; Al-Karmalawy, A.A. Computational repurposing of benzimidazole anthelmintic drugs as potential colchicine binding site inhibitors. Future Med. Chem. 2021, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Alesawy, M.S. Design and discovery of new 1,2,4-triazolo[4,3-c] quinazolines as potential DNA intercalators and topoisomerase II inhibitors. Arch. Pharm. 2020, 3, e2000237. [Google Scholar]
- Feoktistova, M.; Geserick, P.; Leverkus, M. Crystal violet assay for determining viability of cultured cells. Cold Spring Harb. Protoc. 2016, 2016, 087379. [Google Scholar] [CrossRef]
- Mostafa, A. FDA-Approved Drugs with Potent In Vitro Antiviral Activity against Severe Acute Respiratory Syndrome Coronavirus 2. Pharmaceuticals 2020, 13, 443. [Google Scholar] [CrossRef]
- Ahmed Kandeil, A.M. Coding-complete genome sequences of two SARS-CoV-2 isolates from Egypt. Microbiol. Resour. Announc. 2020, 9, 22. [Google Scholar]


{kind=link}
{kind=link}
| No. | Isolated Compound | S a | RMSD b | Interactions | Distance (Å) |
|---|---|---|---|---|---|
| KI | α-Ketoamide inhibitor | −8.17 | 1.64 | Glu166/H-donor Glu166/H-acceptor Glu166/H-donor Gly143/pi-H | 2.89 3.10 3.42 3.70 |
| 1 | Taxifolin | −6.50 | 1.58 | Arg188/H-donor Glu166/H-donor Cys145/H-donor His41/H-pi | 2.85 3.16 3.60 3.44 |
| 2 | Pectolinarigenin | −5.74 | 1.72 | Glu166/pi-H Met165/pi-H | 4.19 4.47 |
| 3 | Tangeretin | −6.61 | 1.17 | Glu166/pi-H Glu166/pi-H | 4.09 4.19 |
| 4 | Gardenin B | −6.48 | 0.74 | Glu166/pi-H Glu166/pi-H | 4.10 4.28 |
| 5 | Hispidulin | −5.85 | 1.14 | His41/H-pi Glu166/pi-H His41/pi-H | 3.83 3.87 4.32 |
| Isolated Comp. | 3D Binding | 3D Positioning |
|---|---|---|
| α-Ketoamide Inhibitor (KI) |  |  |
| Taxifolin (1) |  |  |
| Pectolinarigenin (2) | 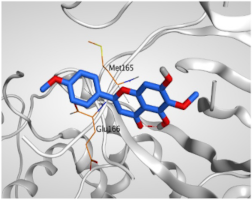 |  |
| Tangeretin (3) |  | 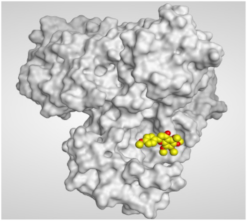 |
| Gardenin B (4) |  |  |
| Hispidulin (5) | 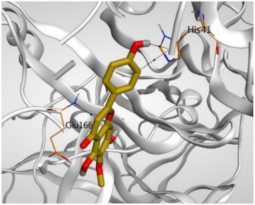 |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Karmalawy, A.A.; Farid, M.M.; Mostafa, A.; Ragheb, A.Y.; H. Mahmoud, S.; Shehata, M.; Shama, N.M.A.; GabAllah, M.; Mostafa-Hedeab, G.; Marzouk, M.M. Naturally Available Flavonoid Aglycones as Potential Antiviral Drug Candidates against SARS-CoV-2. Molecules 2021, 26, 6559. https://doi.org/10.3390/molecules26216559
Al-Karmalawy AA, Farid MM, Mostafa A, Ragheb AY, H. Mahmoud S, Shehata M, Shama NMA, GabAllah M, Mostafa-Hedeab G, Marzouk MM. Naturally Available Flavonoid Aglycones as Potential Antiviral Drug Candidates against SARS-CoV-2. Molecules. 2021; 26(21):6559. https://doi.org/10.3390/molecules26216559
Chicago/Turabian StyleAl-Karmalawy, Ahmed A., Mai M. Farid, Ahmed Mostafa, Alia Y. Ragheb, Sara H. Mahmoud, Mahmoud Shehata, Noura M. Abo Shama, Mohamed GabAllah, Gomaa Mostafa-Hedeab, and Mona M. Marzouk. 2021. "Naturally Available Flavonoid Aglycones as Potential Antiviral Drug Candidates against SARS-CoV-2" Molecules 26, no. 21: 6559. https://doi.org/10.3390/molecules26216559
APA StyleAl-Karmalawy, A. A., Farid, M. M., Mostafa, A., Ragheb, A. Y., H. Mahmoud, S., Shehata, M., Shama, N. M. A., GabAllah, M., Mostafa-Hedeab, G., & Marzouk, M. M. (2021). Naturally Available Flavonoid Aglycones as Potential Antiviral Drug Candidates against SARS-CoV-2. Molecules, 26(21), 6559. https://doi.org/10.3390/molecules26216559