
Deep Membrane Proteome Profiling Reveals Overexpression of Prostate-Specific Membrane Antigen (PSMA) in High-Risk Human Paraganglioma and Pheochromocytoma, Suggesting New Theranostic Opportunity

 ,
,
Abstract
:1. Introduction
2. Results
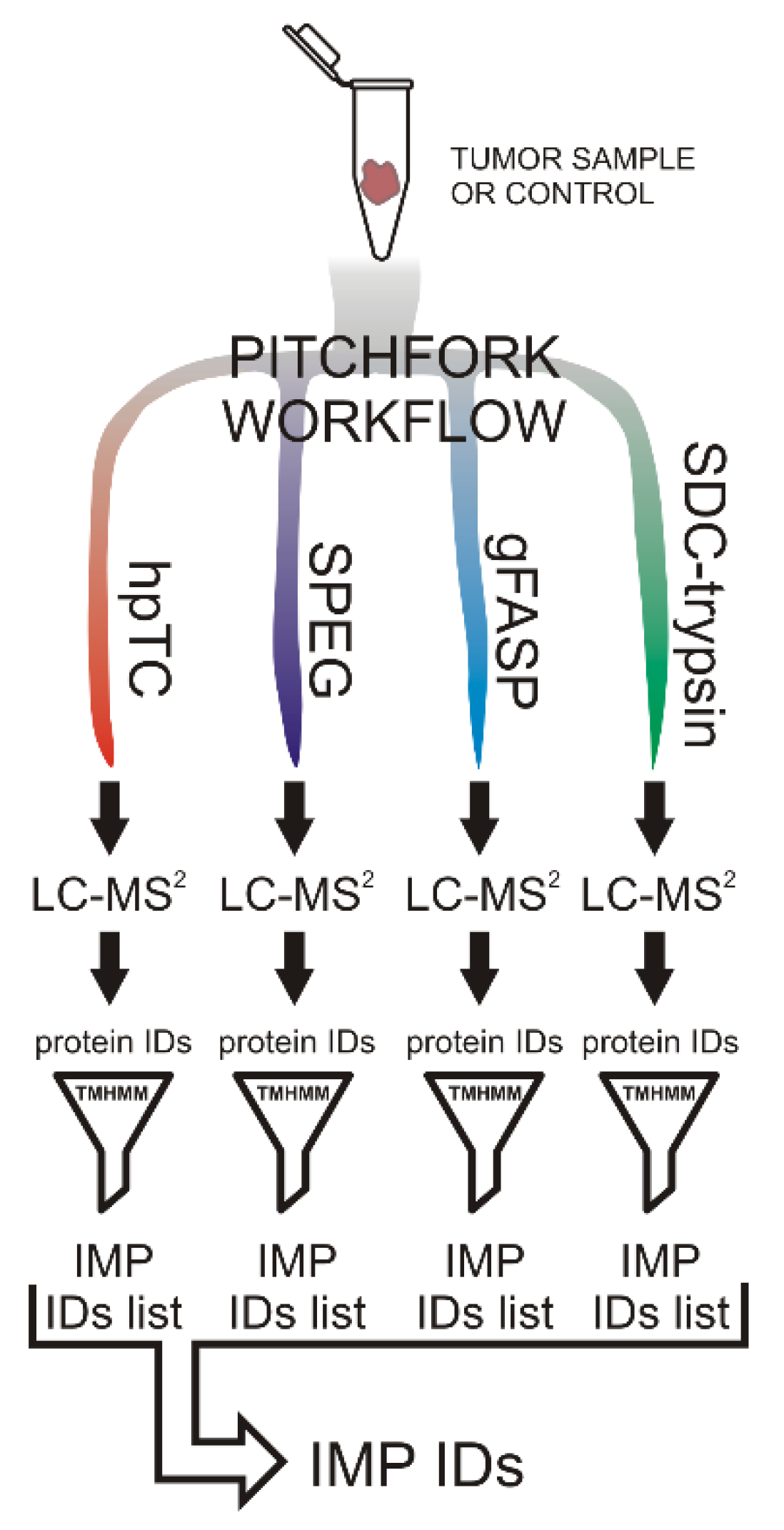
2.1. The Pitchfork Proteomic Analysis
2.2. Verification of PSMA Overexpression
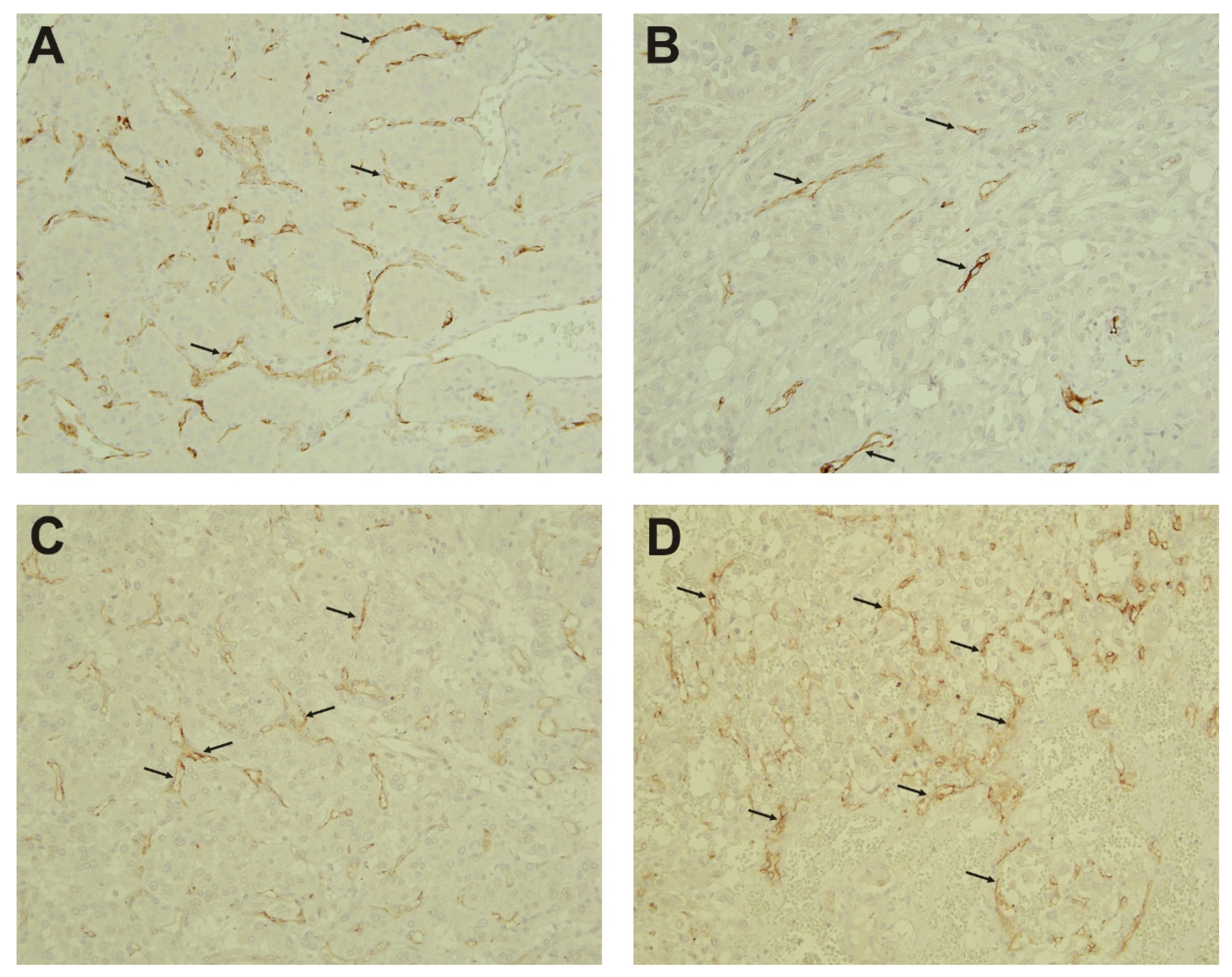
2.3. Immunohistochemistry
3. Discussion
3.1. PSMA—An Effective Theranostic Target
3.2. PSMA Function
3.3. Other Identified Putative Targets Overexpressed in PPGLs
4. Materials and Methods
4.1. Adrenal Medulla Chromaffin Tissue Samples
4.2. Pheochromocytoma and Paraganglioma Samples
4.3. Mutation Screening
4.4. Proteomic Analysis
4.4.1. SPEG
4.4.2. N-glyco-FASP
4.4.3. The High pH Trypsin Cyanogen Bromide (hpTC) Method
4.4.4. SDC-Trypsin
4.4.5. nLC-MS/MS Analysis
4.4.6. Proteomic Data Processing
4.5. SDS-PAGE and Western Blotting
4.6. Immunohistochemistry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Alrezk, R.; Suarez, A.; Tena, I.; Pacak, K. Update of Pheochromocytoma Syndromes: Genetics, Biochemical Evaluation, and Imaging. Front. Endocrinol. 2018, 9, 515. [Google Scholar] [CrossRef]
- Crona, J.; Taïeb, D.; Pacak, K. New Perspectives on Pheochromocytoma and Paraganglioma: Toward a Molecular Classification. Endocr. Rev. 2017, 38, 489–515. [Google Scholar] [CrossRef] [PubMed]
- Hadrava Vanova, K.; Pang, Y.; Krobova, L.; Kraus, M.; Nahacka, Z.; Boukalova, S.; Pack, S.D.; Zobalova, R.; Zhu, J.; Huynh, T.T.; et al. Germline SUCLG2 Variants in Patients with Pheochromocytoma and Paraganglioma. J. Natl. Cancer Inst. 2021, djab158. [Google Scholar] [CrossRef] [PubMed]
- Fishbein, L.; Leshchiner, I.; Walter, V.; Danilova, L.; Robertson, A.G.; Johnson, A.R.; Lichtenberg, T.M.; Murray, B.A.; Ghayee, H.K.; Else, T.; et al. Comprehensive Molecular Characterization of Pheochromocytoma and Paraganglioma. Cancer Cell 2017, 31, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Vit, O.; Petrak, J. Integral membrane proteins in proteomics. How to break open the black box? J. Proteom. 2017, 153, 8–20. [Google Scholar] [CrossRef]
- Vit, O.; Harant, K.; Klener, P.; Man, P.; Petrak, J. A three-pronged “Pitchfork” strategy enables an extensive description of the human membrane proteome and the identification of missing proteins. J. Proteom. 2019, 204, 103411. [Google Scholar] [CrossRef]
- Zielinska, D.F.; Gnad, F.; Wiśniewski, J.R.; Mann, M. Precision mapping of an in vivo N-glycoproteome reveals rigid topological and sequence constraints. Cell 2010, 141, 897–907. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Zhou, Y.; Elliott, S.; Aebersold, R.; Zhang, H. Solid-phase extraction of N-linked glycopeptides. Nat. Protoc. 2007, 2, 334–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vit, O.; Man, P.; Kadek, A.; Hausner, J.; Sklenar, J.; Harant, K.; Novak, P.; Scigelova, M.; Woffendin, G.; Petrak, J. Large-scale identification of membrane proteins based on analysis of trypsin-protected transmembrane segments. J. Proteom. 2016, 149, 15–22. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Waas, M.; Snarrenberg, S.T.; Littrell, J.; Jones Lipinski, R.A.; Hansen, P.A.; Corbett, J.A.; Gundry, R.L. SurfaceGenie: A web-based application for prioritizing cell-type-specific marker candidates. Bioinformatics 2020, 36, 3447–3456. [Google Scholar] [CrossRef] [PubMed]
- Sartor, O.; de Bono, J.; Chi, K.N.; Fizazi, K.; Herrmann, K.; Rahbar, K.; Tagawa, S.T.; Nordquist, L.T.; Vaishampayan, N.; El-Haddad, G.; et al. Lutetium-177-PSMA-617 for Metastatic Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2021, 385, 1091–1103. [Google Scholar] [CrossRef]
- Parsi, M.; Desai, M.H.; Desai, D.; Singhal, S.; Khandwala, P.M.; Potdar, R.R. PSMA: A game changer in the diagnosis and treatment of advanced prostate cancer. Med. Oncol. 2021, 38, 89. [Google Scholar] [CrossRef]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sácha, P.; Zámecník, J.; Barinka, C.; Hlouchová, K.; Vícha, A.; Mlcochová, P.; Hilgert, I.; Eckschlager, T.; Konvalinka, J. Expression of glutamate carboxypeptidase II in human brain. Neuroscience 2007, 144, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Moy, P.; Kim, S.; Xia, Y.; Rajasekaran, A.; Navarro, V.; Knudsen, B.; Bander, N.H. Monoclonal antibodies to the extracellular domain of prostate-specific membrane antigen also react with tumor vascular endothelium. Cancer Res. 1997, 57, 3629–3634. [Google Scholar]
- Chang, S.S.; O’Keefe, D.S.; Bacich, D.J.; Reuter, V.E.; Heston, W.D.; Gaudin, P.B. Prostate-specific membrane antigen is produced in tumor-associated neovasculature. Clin. Cancer Res. 1999, 5, 2674–2681. [Google Scholar]
- Uijen, M.; Derks, Y.; Merkx, R.; Schilham, M.; Roosen, J.; Privé, B.M.; van Lith, S.; van Herpen, C.; Gotthardt, M.; Heskamp, S.; et al. PSMA radioligand therapy for solid tumors other than prostate cancer: Background, opportunities, challenges, and first clinical reports. Eur. J. Nucl. Med. Mol. Imaging 2021. Available online: https://link.springer.com/article/10.1007%2Fs00259-021-05433-w (accessed on 29 October 2021). [CrossRef] [PubMed]
- Van de Wiele, C.; Sathekge, M.; de Spiegeleer, B.; De Jonghe, P.J.; Debruyne, P.R.; Borms, M.; Beels, L.; Maes, A. PSMA expression on neovasculature of solid tumors. Histol. Histopathol. 2020, 35, 919–927. [Google Scholar] [CrossRef]
- Hyväkkä, A.; Virtanen, V.; Kemppainen, J.; Grönroos, T.J.; Minn, H.; Sundvall, M. More Than Meets the Eye: Scientific Rationale behind Molecular Imaging and Therapeutic Targeting of Prostate-Specific Membrane Antigen (PSMA) in Metastatic Prostate Cancer and Beyond. Cancers 2021, 13, 2244. [Google Scholar] [CrossRef]
- Singh, D.; Kumar, R.; Mittal, B.R.; Singh, H.; Bhattacharya, A. 68Ga-Labeled Prostate-Specific Membrane Antigen Uptake in Pheochromocytoma: An Incidental Finding in PET/CT Scan. Clin. Nucl. Med. 2018, 43, 688–690. [Google Scholar] [CrossRef]
- Parihar, A.S.; Vadi, S.K.; Mittal, B.R.; Kumar, R.; Bal, A.; Singh, S.K. 68Ga-PSMA-HBED-CC-Avid Synchronous Urinary Bladder Paraganglioma in a Patient With Metastatic Prostate Carcinoma. Clin. Nucl. Med. 2018, 43, e329–e330. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Singh, S.K.; Narain, T.A.; Singh, H.; Kumar, R.; Vadi, S.K. Synchronous Paraganglioma Masquerading as 68Ga-PSMA PET/CT-Avid Metastasis in Carcinoma Prostate-How Specific Is 68Ga-PSMA PET/CT? Clin. Nucl. Med. 2019, 44, e420–e422. [Google Scholar] [CrossRef]
- Tripathy, S.; Tripathi, M.; Dattagupta, S.; Parida, G.K.; Shamim, S.A. In Vivo PSMA Expression in Head and Neck Paragangliomas on 68Ga PSMA 11 PET/CT. Clin. Nucl. Med. 2019, 44, e398–e400. [Google Scholar] [CrossRef]
- Navalkissoor, S.; Grossman, A. Targeted Alpha Particle Therapy for Neuroendocrine Tumours: The Next Generation of Peptide Receptor Radionuclide Therapy. Neuroendocrinology 2019, 108, 256–264. [Google Scholar] [CrossRef]
- Turkova, H.; Prodanov, T.; Maly, M.; Martucci, V.; Adams, K.; Widimsky, J., Jr.; Chen, C.C.; Ling, A.; Kebebew, E.; Stratakis, C.A.; et al. Characteristics and Outcomes of Metastatic Sdhb and Sporadic Pheochromocytoma/paraganglioma: An National Institutes of Health Study. Endocr. Pract. 2016, 22, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Amar, L.; Baudin, E.; Burnichon, N.; Peyrard, S.; Silvera, S.; Bertherat, J.; Bertagna, X.; Schlumberger, M.; Jeunemaitre, X.; Gimenez-Roqueplo, A.P.; et al. Succinate dehydrogenase B gene mutations predict survival in patients with malignant pheochromocytomas or paragangliomas. J. Clin. Endocrinol. Metab. 2007, 92, 3822–3828. [Google Scholar] [CrossRef]
- Gimenez-Roqueplo, A.P.; Favier, J.; Rustin, P.; Rieubland, C.; Crespin, M.; Nau, V.; Khau Van Kien, P.; Corvol, P.; Plouin, P.F.; Jeunemaitre, X.; et al. Mutations in the SDHB gene are associated with extra-adrenal and/or malignant phaeochromocytomas. Cancer Res. 2003, 63, 5615–5621. [Google Scholar]
- Binderup, M.L.; Jensen, A.M.; Budtz-Jørgensen, E.; Bisgaard, M.L. Survival and causes of death in patients with von Hippel-Lindau disease. J. Med. Genet. 2017, 54, 11–18. [Google Scholar] [CrossRef]
- Janssen, I.; Blanchet, E.M.; Adams, K.; Chen, C.C.; Millo, C.M.; Herscovitch, P.; Taieb, D.; Kebebew, E.; Lehnert, H.; Fojo, A.T.; et al. Superiority of [68Ga]-DOTATATE PET/CT to Other Functional Imaging Modalities in the Localization of SDHB-Associated Metastatic Pheochromocytoma and Paraganglioma. Clin. Cancer Res. 2015, 21, 3888–3895. [Google Scholar] [CrossRef] [Green Version]
- Janssen, I.; Chen, C.C.; Millo, C.M.; Ling, A.; Taieb, D.; Lin, F.I.; Adams, K.T.; Wolf, K.I.; Herscovitch, P.; Fojo, A.T.; et al. PET/CT comparing (68)Ga-DOTATATE and other radiopharmaceuticals and in comparison with CT/MRI for the localization of sporadic metastatic pheochromocytoma and paraganglioma. Eur. J. Nucl. Med. Mol. Imaging 2016, 43, 1784–1791. [Google Scholar] [CrossRef]
- Taïeb, D.; Hicks, R.J.; Hindié, E.; Guillet, B.A.; Avram, A.; Ghedini, P.; Timmers, H.J.; Scott, A.T.; Elojeimy, S.; Rubello, D.; et al. European Association of Nuclear Medicine Practice Guideline/Society of Nuclear Medicine and Molecular Imaging Procedure Standard 2019 for radionuclide imaging of phaeochromocytoma and paraganglioma. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2112–2137. [Google Scholar] [CrossRef]
- Jha, A.; Patel, M.; Carrasquillo, J.A.; Ling, A.; Millo, C.; Saboury, B.; Chen, C.C.; Wakim, P.; Gonzales, M.K.; Meuter, L.; et al. Sporadic Primary Pheochromocytoma: A Prospective Intra-Individual Comparison of Six Imaging Tests (CT, MRI, and PET/CT Using 68Ga-DOTATATE, FDG, 18F-FDOPA, and 18F-FDA). AJR Am. J. Rroentgenol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Mesters, J.R.; Barinka, C.; Li, W.; Tsukamoto, T.; Majer, P.; Slusher, B.S.; Konvalinka, J.; Hilgenfeld, R. Structure of glutamate carboxypeptidase II, a drug target in neuronal damage and prostate cancer. EMBO. J. 2006, 25, 1375–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halsted, C.H. Jejunal brush-border folate hydrolase. A novel enzyme. West. J. Med. 1991, 155, 605–609. [Google Scholar] [PubMed]
- Sheehan, B.; Guo, C.; Neeb, A.; Paschalis, A.; Sandhu, S.; de Bono, J.S. Prostate-specific Membrane Antigen Biology in Lethal Prostate Cancer and its Therapeutic Implications. Eur. Urol. Focus 2021. Available online: https://linkinghub.elsevier.com/retrieve/pii/S2405-4569(21)00168-1 (accessed on 29 October 2021). [CrossRef] [PubMed]
- Yao, V.; Berkman, C.E.; Choi, J.K.; O’Keefe, D.S.; Bacich, D.J. Expression of prostate-specific membrane antigen (PSMA), increases cell folate uptake and proliferation and suggests a novel role for PSMA in the uptake of the non-polyglutamated folate, folic acid. Prostate 2010, 70, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Conway, R.E.; Petrovic, N.; Li, Z.; Heston, W.; Wu, D.; Shapiro, L.H. Prostate-specific membrane antigen regulates angiogenesis by modulating integrin signal transduction. Mol. Cell Biol. 2006, 26, 5310–5324. [Google Scholar] [CrossRef] [Green Version]
- Conway, R.E.; Joiner, K.; Patterson, A.; Bourgeois, D.; Rampp, R.; Hannah, B.C.; McReynolds, S.; Elder, J.M.; Gilfilen, H.; Shapiro, L.H. Prostate specific membrane antigen produces pro-angiogenic laminin peptides downstream of matrix metalloprotease-2. Angiogenesis 2013, 16, 847–860. [Google Scholar] [CrossRef]
- Conway, R.E.; Rojas, C.; Alt, J.; Nováková, Z.; Richardson, S.M.; Rodrick, T.C.; Fuentes, J.L.; Richardson, N.H.; Attalla, J.; Stewart, S.; et al. Prostate-specific membrane antigen (PSMA)-mediated laminin proteolysis generates a pro-angiogenic peptide. Angiogenesis 2016, 19, 487–500. [Google Scholar] [CrossRef]
- Robinson, J.; Whitworth, K.; Jinks, E.; Nagy, Z.; Bicknell, R.; Lee, S.P. An evaluation of the tumour endothelial marker CLEC14A as a therapeutic target in solid tumours. J. Pathol. Clin. Res. 2020, 6, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Riether, C.; Radpour, R.; Kallen, N.M.; Bürgin, D.T.; Bachmann, C.; Schürch, C.M.; Lüthi, U.; Arambasic, M.; Hoppe, S.; Albers, C.E.; et al. Metoclopramide treatment blocks CD93-signaling-mediated self-renewal of chronic myeloid leukemia stem cells. Cell Rep. 2021, 34, 108663. [Google Scholar] [CrossRef] [PubMed]
- Sier, V.Q.; van der Vorst, J.R.; Quax, P.; de Vries, M.R.; Zonoobi, E.; Vahrmeijer, A.L.; Dekkers, I.A.; de Geus-Oei, L.F.; Smits, A.M.; Cai, W.; et al. Endoglin/CD105-Based Imaging of Cancer and Cardiovascular Diseases: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 4804. [Google Scholar] [CrossRef]
- Petrovic, K.; Robinson, J.; Whitworth, K.; Jinks, E.; Shaaban, A.; Lee, S.P. TEM8/ANTXR1-specific CAR T cells mediate toxicity in vivo. PLoS ONE 2019, 14, e0224015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katchman, B.A.; Ocal, I.T.; Cunliffe, H.E.; Chang, Y.H.; Hostetter, G.; Watanabe, A.; LoBello, J.; Lake, D.F. Expression of quiescin sulfhydryl oxidase 1 is associated with a highly invasive phenotype and correlates with a poor prognosis in Luminal B breast cancer. Breast Cancer Res. 2013, 15, R28. [Google Scholar] [CrossRef] [Green Version]
- Baek, J.A.; Song, P.H.; Ko, Y.; Gu, M.J. High expression of QSOX1 is associated with tumor invasiveness and high grades groups in prostate cancer. Pathol. Res. Pract. 2018, 214, 964–967. [Google Scholar] [CrossRef]
- Feldman, T.; Grossman-Haham, I.; Elkis, Y.; Vilela, P.; Moskovits, N.; Barshack, I.; Salame, T.M.; Fass, D.; Ilani, T. Inhibition of fibroblast secreted QSOX1 perturbs extracellular matrix in the tumor microenvironment and decreases tumor growth and metastasis in murine cancer models. Oncotarget 2020, 11, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Fifield, A.L.; Hanavan, P.D.; Faigel, D.O.; Sergienko, E.; Bobkov, A.; Meurice, N.; Petit, J.L.; Polito, A.; Caulfield, T.R.; Castle, E.P.; et al. Molecular Inhibitor of QSOX1 Suppresses Tumor Growth In Vivo. Mol. Cancer Ther. 2020, 19, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crona, J.; Lamarca, A.; Ghosal, S.; Welin, S.; Skogseid, B.; Pacak, K. Genotype-phenotype correlations in pheochromocytoma and paraganglioma: A systematic review and individual patient meta-analysis. Endocr-Relat. Cancer 2019, 26, 539–550. [Google Scholar] [CrossRef]
- Zelinka, T.; Eisenhofer, G.; Pacak, K. Pheochromocytoma as a catecholamine producing tumor: Implications for clinical practice. Stress 2007, 10, 195–203. [Google Scholar] [CrossRef]
- Lenders, J.; Kerstens, M.N.; Amar, L.; Prejbisz, A.; Robledo, M.; Taieb, D.; Pacak, K.; Crona, J.; Zelinka, T.; Mannelli, M.; et al. Genetics, diagnosis, management and future directions of research of phaeochromocytoma and paraganglioma: A position statement and consensus of the Working Group on Endocrine Hypertension of the European Society of Hypertension. J. Hypertens. 2020, 38, 1443–1456. [Google Scholar] [CrossRef]
- Eisenhofer, G.; Lenders, J.W.; Siegert, G.; Bornstein, S.R.; Friberg, P.; Milosevic, D.; Mannelli, M.; Linehan, W.M.; Adams, K.; Timmers, H.J.; et al. Plasma methoxytyramine: A novel biomarker of metastatic pheochromocytoma and paraganglioma in relation to established risk factors of tumour size, location and SDHB mutation status. Eur. J. Cancer 2012, 48, 1739–1749. [Google Scholar] [CrossRef] [Green Version]
- Nölting, S.; Ullrich, M.; Pietzsch, J.; Ziegler, C.G.; Eisenhofer, G.; Grossman, A.; Pacak, K. Current Management of Pheochromocytoma/Paraganglioma: A Guide for the Practicing Clinician in the Era of Precision Medicine. Cancers 2019, 11, 1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, T.; Tomita, M.; Ishihama, Y. Phase transfer surfactant-aided trypsin digestion for membrane proteome analysis. J. Proteome Res. 2008, 7, 731–740. [Google Scholar] [CrossRef]
- Sun, Z.; Qin, H.; Wang, F.; Cheng, K.; Dong, M.; Ye, M.; Zou, H. Capture and dimethyl labeling of glycopeptides on hydrazide beads for quantitative glycoproteomics analysis. Anal. Chem. 2012, 84, 8452–8456. [Google Scholar] [CrossRef]
- Rey, M.; Mrázek, H.; Pompach, P.; Novák, P.; Pelosi, L.; Brandolin, G.; Forest, E.; Havlícek, V.; Man, P. Effective removal of nonionic detergents in protein mass spectrometry, hydrogen/deuterium exchange, and proteomics. Anal. Chem. 2010, 82, 5107–5116. [Google Scholar] [CrossRef] [PubMed]
- Hebert, A.S.; Richards, A.L.; Bailey, D.J.; Ulbrich, A.; Coughlin, E.E.; Westphall, M.S.; Coon, J.J. The one hour yeast proteome. Mol. Cell Proteom. 2014, 13, 339–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient No. | Sex | Tumor Type | PPGL Cluster | Mutated Gene | Used in Proteomics (P) and/or Western Blotting |
|---|---|---|---|---|---|
| 1 | F | PGL | 1 | SDHB | P,W |
| 2 | F | PGL | 1 | SDHB | P,W |
| 3 | M | PGL | 1 | SDHB | P,W |
| 4 | M | PGL | 1 | SDHB | W |
| 5 | M | PGL | 1 | SDHB | W |
| 6 | F | PGL | 1 | SDHB | W |
| 7 | M | PGL | 1 | SDHB | W |
| 8 | F | PGL | 1 | SDHB | W |
| 9 | F | PGL | 1 | SDHB | W |
| 10 | M | PHEO | 1 | VHL | W |
| 11 | F | PHEO | 1 | VHL | W |
| 12 | F | PHEO | 1 | VHL | W |
| 13 | M | PGL | 1 | VHL | P,W |
| 14 | M | PGL | 1 | VHL | P,W |
| 15 | F | PHEO | 1 | EPAS1 | W |
| 16 | M | PHEO | 2 | RET | W |
| 17 | F | PHEO | 2 | RET | W |
| 18 | M | PHEO | 2 | RET | W |
| 19 | F | PHEO | 2 | RET | W |
| 20 | F | PHEO | NA | Sporadic | W |
| 21 | F | PHEO | NA | Sporadic | W |
| 22 | M | PHEO | NA | Sporadic | W |
| Accession | Protein Name | Gene Name | TM Segments | SPC Score |
|---|---|---|---|---|
| A8MWY0 | Endosome/lysosome-associated apoptosis and autophagy regulator family member 2 | ELAPOR2 | 1 | 3 |
| Q86T13 | C-type lectin domain family 14 member A (Epidermal growth factor receptor 5) | CLEC14A | 1 | 3 |
| Q9Y5G0 | Protocadherin gamma-B5 | PCDHGB5 | 1 | 3 |
| Q9UNN8 | Endothelial protein C receptor (CD antigen CD201) | PROCR | 1 | 4 |
| Q04609 | Glutamate carboxypeptidase 2, (Prostate-specific membrane antigen, PSMA) | FOLH1 | 1 | 3 |
| Q96EP9 | Sodium/bile acid cotransporter 4 (Solute carrier family 10 member 4) | SLC10A4 | 7 | 3 |
| Q9ULK0 | Glutamate receptor ionotropic, delta-1 (GluD1) | GRID1 | 3 | 3 |
| Q13936 | Voltage-dependent L-type calcium channel subunit alpha-1C | CACNA1C | 24 | 3 |
| O00391 | Sulfhydryl oxidase 1 | QSOX1 | 1 | 3 |
| P80370 | Protein delta homolog 1 (DLK-1) | DLK1 | 1 | 3 |
| Q9NQX7 | Integral membrane protein 2C (Cerebral protein 14) | ITM2C | 1 | 3 |
| P09619 | Platelet-derived growth factor receptor beta (PDGF-R-beta) (CD antigen CD140b) | PDGFRB | 1 | 4 |
| Q9NPY3 | Complement component C1q receptor (CD antigen CD93) | CD93 | 1 | 4 |
| Q9H6X2 | Anthrax toxin receptor 1 (Tumor endothelial marker 8) | ANTXR1 | 1 | 4 |
| Q9H2H9 | Sodium-coupled neutral amino acid transporter 1 (Solute carrier family 38 member 1) | SLC38A1 | 11 | 4 |
| O75882 | Attractin | ATRN | 1 | 4 |
| P17813 | Endoglin (CD105) | ENG | 1 | 4 |
| Q9HCJ1 | Progressive ankylosis protein homolog (ANK) | ANKH | 8 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vit, O.; Patel, M.; Musil, Z.; Hartmann, I.; Frysak, Z.; Miettinen, M.; Pacak, K.; Petrak, J. Deep Membrane Proteome Profiling Reveals Overexpression of Prostate-Specific Membrane Antigen (PSMA) in High-Risk Human Paraganglioma and Pheochromocytoma, Suggesting New Theranostic Opportunity. Molecules 2021, 26, 6567. https://doi.org/10.3390/molecules26216567
Vit O, Patel M, Musil Z, Hartmann I, Frysak Z, Miettinen M, Pacak K, Petrak J. Deep Membrane Proteome Profiling Reveals Overexpression of Prostate-Specific Membrane Antigen (PSMA) in High-Risk Human Paraganglioma and Pheochromocytoma, Suggesting New Theranostic Opportunity. Molecules. 2021; 26(21):6567. https://doi.org/10.3390/molecules26216567
Chicago/Turabian StyleVit, Ondrej, Mayank Patel, Zdenek Musil, Igor Hartmann, Zdenek Frysak, Markku Miettinen, Karel Pacak, and Jiri Petrak. 2021. "Deep Membrane Proteome Profiling Reveals Overexpression of Prostate-Specific Membrane Antigen (PSMA) in High-Risk Human Paraganglioma and Pheochromocytoma, Suggesting New Theranostic Opportunity" Molecules 26, no. 21: 6567. https://doi.org/10.3390/molecules26216567