
Diterpenes/Diterpenoids and Their Derivatives as Potential Bioactive Leads against Dengue Virus: A Computational and Network Pharmacology Study

 ,
,  ,
, 
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Attribution of Proteins’ Active Sites and Validation
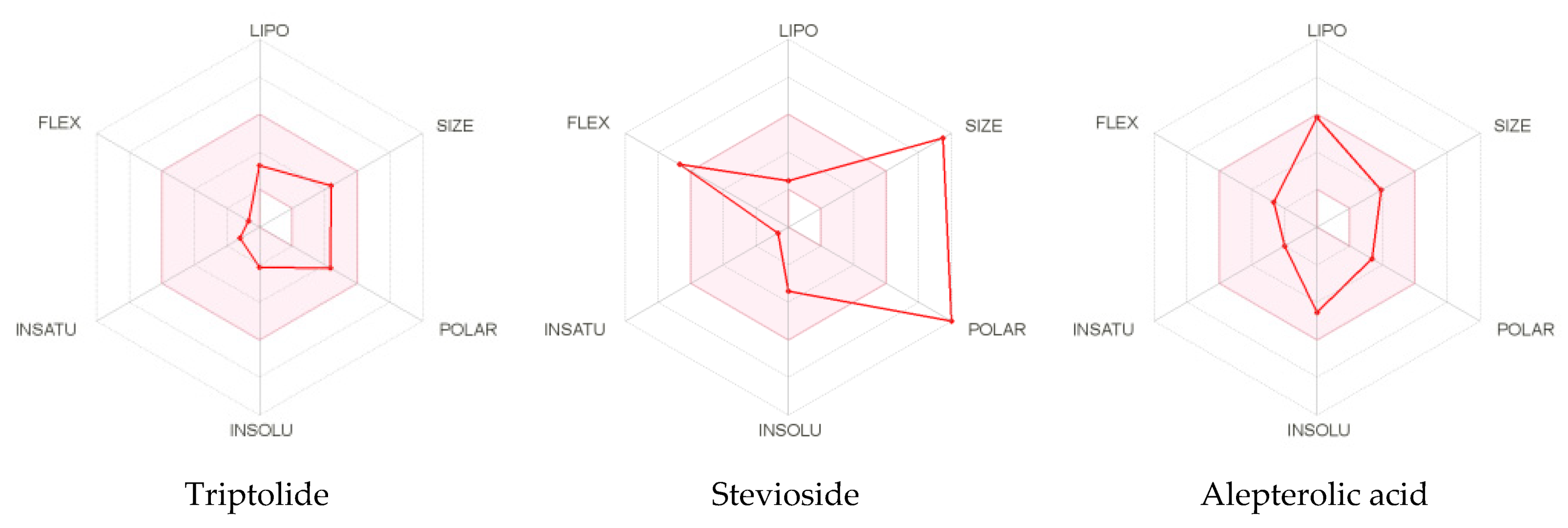
2.2. Computational Virtual Screening of Diterpenoids and Their Derivatives
ADMET Analysis
2.3. Molecular Docking
2.3.1. Docking Approach of Natural Bioactive against DENV Receptor Proteins
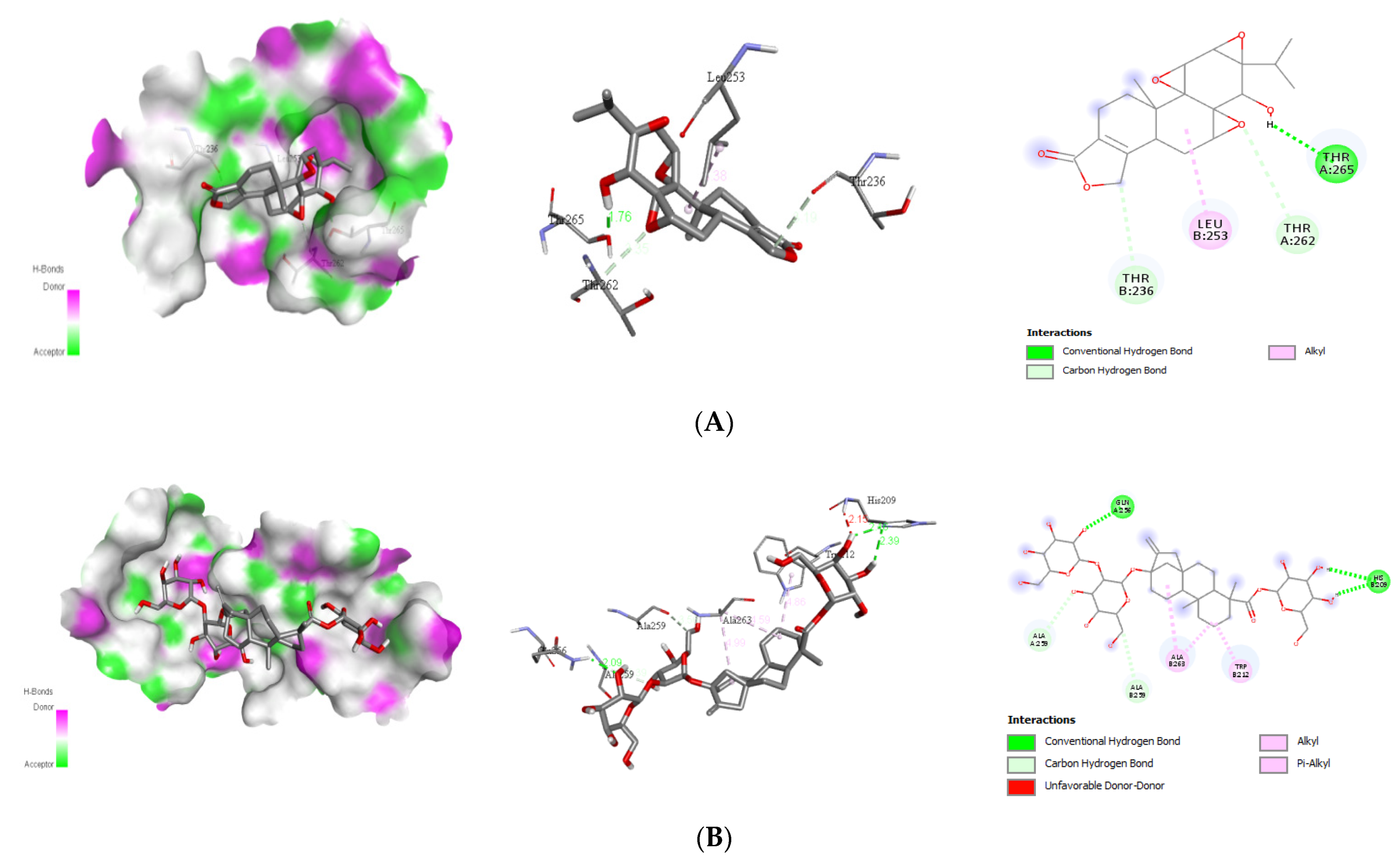
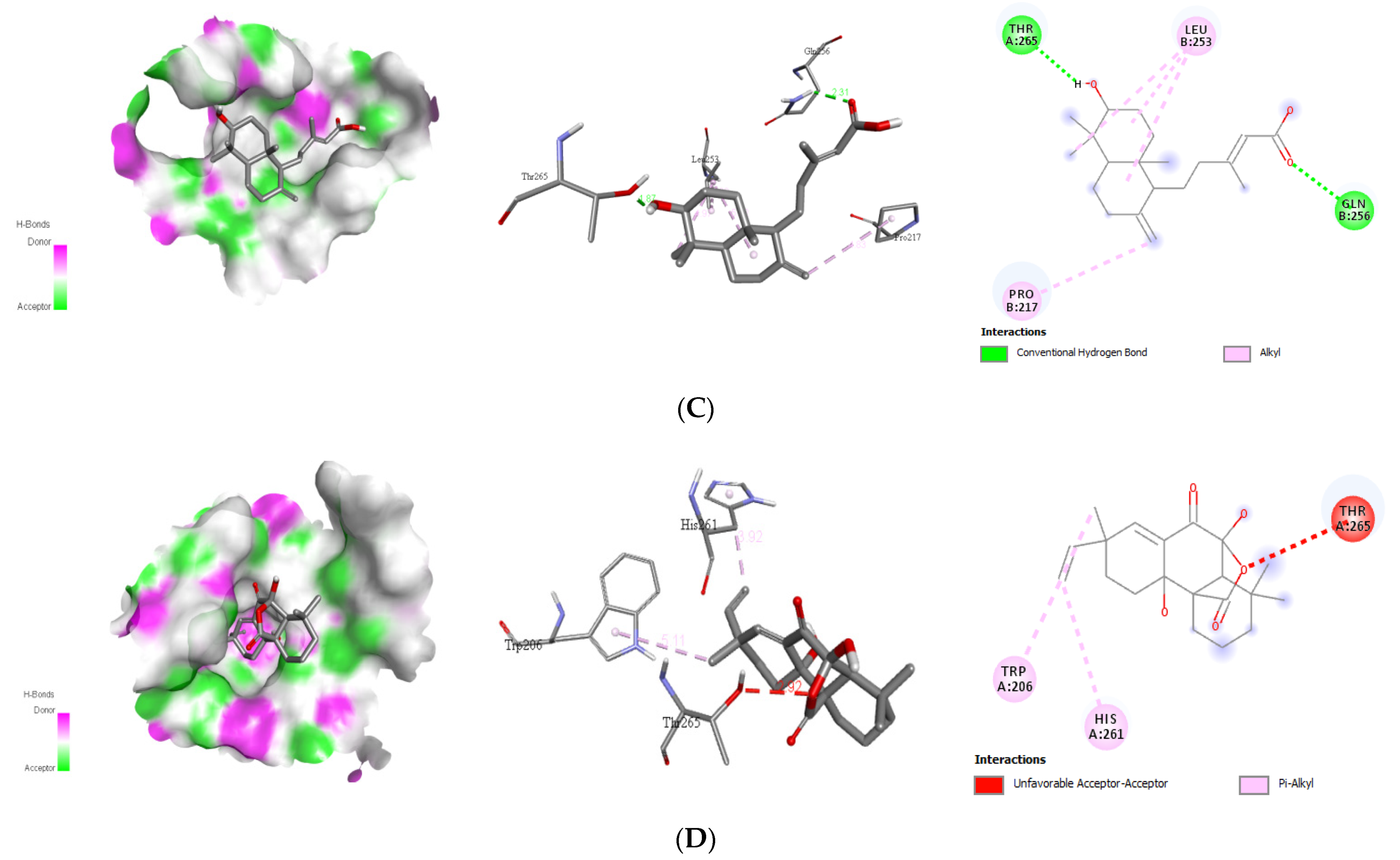
Interaction with Viral Envelope (E) Protein
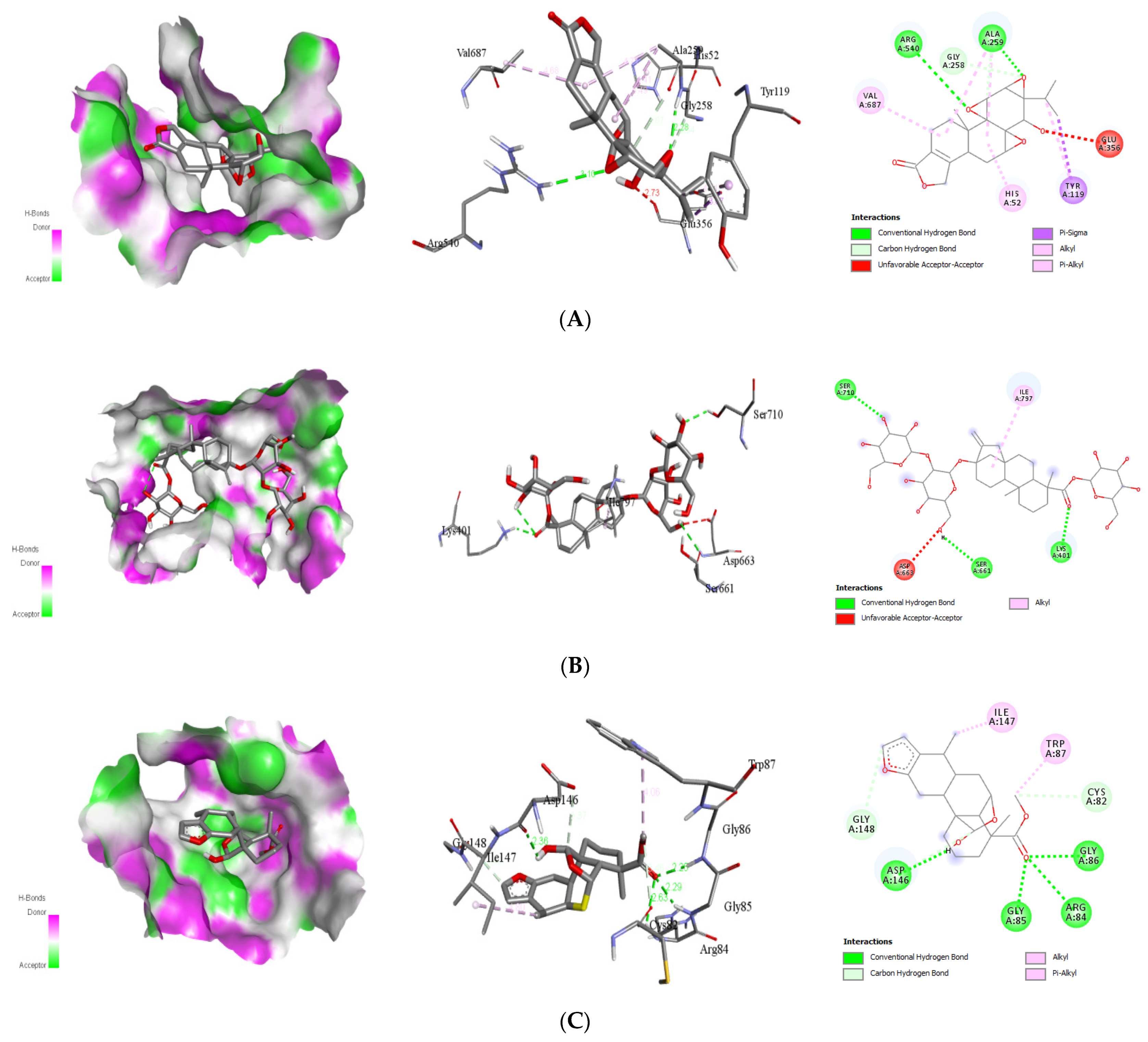
Interaction with Viral NS3
Interaction with Viral NS5
Interaction with Viral NS1
2.3.2. Docking Approach of Chemical Analog (Pyrimethamine) against DENV Proteins
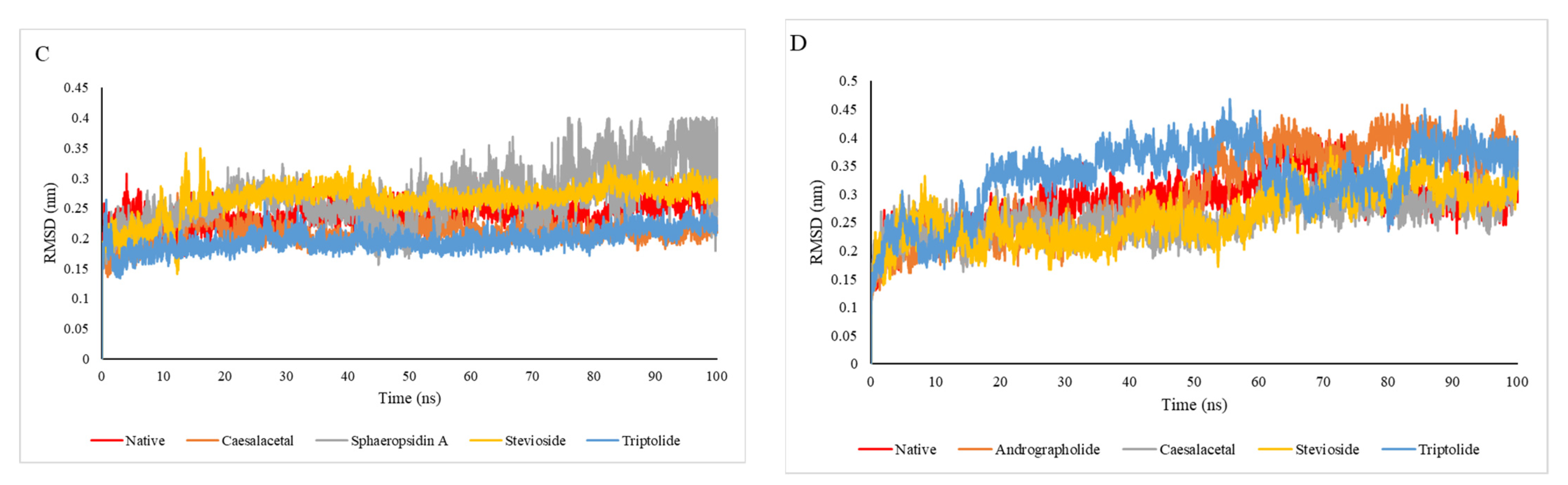
2.4. Molecular Dynamic Simulation Analysis
2.5. MM-PBSA Analysis
2.6. Network Pharmacology of Diterpenoids
2.6.1. Gene Set Enrichment Analysis
2.6.2. Construction of “Drug-Target-Pathway” Network
3. Materials and Methods
3.1. Protein/Macromolecule Structure Preparation
3.2. Active Site Prediction
3.3. Selection and Preparation of Ligands
3.4. In Silico Pharmacokinetic Study
3.5. Molecular Docking Protocol
3.6. Molecular Dynamics (MD) Simulation Study
3.7. Molecular Mechanics Poisson–Boltzmann Surface Area (MMPBSA) Analysis
3.8. Network Pharmacology of Diterpenoid
4. Conclusions and Final Considerations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Vaddadi, S.; Vaddadi, R.S. Dengue fever: Are view article. J. Evol. Med. Dent. Sci. 2015, 4, 5048–5058. [Google Scholar]
- Sood, S.K.; Sood, V.; Mahajan, I.; Sahil. An intelligent healthcare system for predicting and preventing dengue virus infection. Comput. Comput. 2021, 1–39. [Google Scholar] [CrossRef]
- Henchal, E.A.; Putnak, J.R. The dengue viruses. Clin. Microbiol. Rev. 1990, 3, 376–396. [Google Scholar] [CrossRef]
- Chambers, T. Flaavi virus genome organization, expression, and replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Ross, T.M.; Neville, S.M.; Innes, D.S.; Turner, D.R.; Moubaraki, B.; Murray, K.S. Spin crossover in iron (III) Schiff-base 1-D chain complexes. Dalton Trans. 2010, 39, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Tassaneetrithep, B.; Burgess, T.H.; Granelli-Piperno, A.; Trumpfheller, C.; Finke, J.; Sun, W.; Eller, M.A.; Pattanapanyasat, K.; Sarasombath, S.; Birx, D.L.; et al. DC-SIGN (CD 209) mediates dengue virus infection of human dendritic cells. J. Exp. Med. 2003, 197, 823–829. [Google Scholar] [CrossRef] [Green Version]
- Heinz, F.X.; Auer, G.; Stiasny, K.; Holzmann, H.; Mandl, C.; Guirakhoo, F.; Kunz, C. The interactions of the flavivirus envelope proteins: Implications for virus entry and release. Arch. Virol. Suppl. 1994, 9, 339–348. [Google Scholar]
- Heinz, F.X.; Stiasny, K.; Püschner-Auer, G.; Holzmann, H.; Allison, S.L.; Mandl, C.W.; Kunz, C. Structural changes and functional control of the tick-borne encephalitis virus glycoprotein E by the heterodimeric association with protein prM. Virology 1994, 198, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.M.; Jones, M.K.; Westaway, E.G. Markers for trans-Golgi membranes and the intermediate compartment localize to induced membranes with distinct replication functions in flavivirus-infected cells. J. Virol. 1999, 73, 9555–9567. [Google Scholar] [CrossRef] [Green Version]
- Halstead, S.B. Pathogenesis of dengue: Challenges to molecular biology. Science 1988, 239, 476–481. [Google Scholar] [CrossRef]
- Monath, T.P. Yellow fever and dengue—The interactions of virus, vector and host in there-emergence of epidemic disease. Semin. Virol. 1994, 5, 133–145. [Google Scholar] [CrossRef]
- Rodhain, F. The situation of dengue in the world. Bull. Soc. Pathol. Exot. 1996, 89, 87–90. [Google Scholar] [PubMed]
- Rigau-Pérez, J.G.; Clark, G.G.; Gubler, D.J.; Reiter, P.; Sanders, E.J.; Vorndam, A.V. Dengue and dengue haemorrhagic fever. Lancet 1998, 352, 971–977. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Abdalgader, T.; Pedersen, M.; Ren, D.; Zhang, L. Trade-off between climatic and human population impacts on Aedes aegypti life-history shapes its geographic distribution. J. Theor. Biol. 2021, 10, 201–206. [Google Scholar]
- Silva, H.H.G.D.; Geris, R.; Rodrigues Filho, E.; Rocha, C.; Silva, I.G.D. Larvicidal activity of oil-resin fractions from the Brazilian medicinal plant Copaifera reticulata Ducke (Leguminosae-Caesalpinioideae) against Aedes aegypti (Diptera, Culicidae). Rev. Soc. Bras. Med. Trop. 2007, 40, 264–267. [Google Scholar]
- Valotto, C.F.B.; da Silva, H.H.G.; Cavasin, G.; Geris, R.; Rodrigues Filho, E.; da Silva, I.G. Ultrastructural alterations in larvae of Aedes aegypti subject to labdane diterpene isolated from Copaifera reticulata (Leguminosae) and a fraction enriched with tannins of Magonia pubescens (Sapindaceae). Rev. Soc. Bras. Med. Trop. 2011, 44, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Blythe, E.K.; Tabanca, N.; Demirci, B.; Tsikolia, M.; Bloomquist, J.R.; Bernier, U.R. L antanamont evidensis Essential Oil: Chemical Composition and Mosquito Repellent Activity against Aedes aegypti. Nat. Prod. Commun. 2016, 11, 1713–1716. [Google Scholar]
- Niu, H.-M.; Zeng, D.-Q.; Long, C.-L.; Peng, Y.-H.; Wang, Y.-H.; Luo, J.-F.; Wang, H.-S.; Shi, Y.-N.; Tang, G.-H.; Zhao, F.-W. Clerodane diterpenoids and prenylated flavonoids from Dodonaea viscosa: Original article. J. Asian Nat. Prod. Res. 2010, 12, 7–14. [Google Scholar] [CrossRef]
- Akihara, Y.; Kamikawa, S.; Harauchi, Y.; Ohta, E.; Nehira, T.; Ômura, H.; Ohta, S. Hydroxylated furanoditerpenoids from pupal cases produced by the bruchid beetle Sulcobruchus sauteri inside the seeds of Caesalpinia decapetala. Phytochemistry 2018, 156, 151–158. [Google Scholar] [CrossRef]
- Luna, E.C.; Luna, I.S.; Scotti, L.; Monteiro, A.F.M.; Scotti, M.T.; de Moura, R.O.; de Araújo, R.S.A.; Monteiro, K.L.C.; de Aquino, T.M.; Ribeiro, F.F.; et al. Active essential oils and their components in use against neglected diseases and arboviruses. Oxid. Med. Cell. Longev. 2019, 2019, 6587150. [Google Scholar] [CrossRef] [Green Version]
- Noble, C.G.; Chen, Y.-L.; Dong, H.; Gu, F.; Lim, S.P.; Schul, W.; Wang, Q.-Y.; Shi, P.-Y. Strategies for development of Dengue virus inhibitors. Antivir. Res. 2010, 85, 450–462. [Google Scholar] [CrossRef]
- Islam, M.T.; Mubarak, M.S. Diterpenes and their derivatives as promising agents against dengue virus and dengue vectors: A literature-based review. Phytother. Res. 2020, 34, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Dundas, J.; Ouyang, Z.; Tseng, J.; Binkowski, A.; Turpaz, Y.; Liang, J. CASTp: Computed atlas of surface topography of proteins with structural and topographical mapping of functionally annotated residues. Nucleic Acids Res. 2006, 34, W116–W118. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-J.; Jiang, Z.-Z.; Zhang, L.-Y. Triptolide: Progress on research in pharmacodynamics and toxicology. J. Ethnopharmacol. 2014, 155, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Brinker, A.M.; Raskin, I. Determination of triptolide in root extracts of Tripterygium wilfordii by solid-phase extraction and reverse-phase high-performance liquid chromatography. J. Chromatogr. A 2005, 1070, 65–70. [Google Scholar] [CrossRef]
- Liou, J.T.; Chen, Z.Y.; Ho, L.J.; Yang, S.P.; Chang, D.M.; Liang, C.C.; Lai, J.H. Differential effects of triptolide and tetrandrine on activation of COX-2, NF-κB, and AP-1 and virus production in dengue virus-infected human lung cells. Eur. J. Pharmacol. 2008, 589, 288–298. [Google Scholar] [CrossRef]
- Wan, Z.; Chen, X. Triptolide inhibits human immunodeficiency virus type 1 replication by promoting proteasomal degradation of Tat protein. Retrovirology 2014, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Guo, W.; Zhou, H.; Wang, J.; Wang, H.; Sun, X. Triptolide decreases expression of latency-associated nuclear antigen 1 and reduces viral titers in Kaposi’s sarcoma-associated and herpesvirus-related primary effusion lymphoma cells. Int. J. Oncol. 2016, 48, 1519–1530. [Google Scholar] [CrossRef] [Green Version]
- Sehar, I.; Kaul, A.; Bani, S.; Pal, H.C.; Saxena, A.K. Immune up regulatory response of a non-caloric natural sweetener, stevioside. Chem. Biol. Interact. 2008, 173, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Loaiza-Cano, V.; Monsalve-Escudero, L.M.; Filho, C.M.; Martinez-Gutierre, M.; De Sousa, D.P. Antiviral Role of Phenolic Compounds against Dengue Virus: A Review. Biomolecules 2020, 11, 11. [Google Scholar] [CrossRef]
- Nguyen, T.T.H.; Si, J.; Kang, C.; Chung, B.; Chung, D.; Kim, D. Facile preparation of water-soluble curcuminoids extracted from turmeric (Curcuma longa L.) powder by using steviolglucosides. Food Chem. 2017, 214, 366–373. [Google Scholar] [CrossRef] [Green Version]
- Gregersen, S.; Jeppesen, P.B.; Holst, J.J.; Hermansen, K. Anti-hyperglycemic effects of stevioside in type 2 diabetic subjects. Metabolism 2004, 53, 73–76. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Avolio, F.; Ali, A.; Tabanca, N.; Khan, I.A.; Evidente, A. Cyclopaldic Acid, Seiridin, and Sphaeropsidin A as Fungal Phytotoxins, and Larvicidal and Biting Deterrents against Aedes aegypti (Diptera: Culicidae): Structure-Activity Relationships. Chem. Biodivers. 2013, 10, 1239–1251. [Google Scholar] [CrossRef]
- Roscetto, E.; Masi, M.; Esposito, M.; Di Lecce, R.; Delicato, A.; Maddau, L.; Calabrò, V.; Evidente, A.; Catania, M.R. Anti-biofilm activity of the fungal phytotoxin sphaeropsidin A against clinical isolates of antibiotic-resistant bacteria. Toxins 2020, 12, 444. [Google Scholar] [CrossRef]
- Mathieu, V.; Chantôme, A.; Lefranc, F.; Cimmino, A.; Miklos, W.; Paulitschke, V.; Mohr, T.; Maddau, L.; Kornienko, A.; Berger, W.; et al. Sphaeropsidin A shows promising activity against drug-resistant cancer cells by targeting regulatory volume increase. Cell. Mol. Life Sci. 2015, 72, 3731–3746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afolayan, M.; Srivedavyasasri, R.; Asekun, O.T.; Familoni, O.B.; Orishadipe, A.; Zulfiqar, F.; Ibrahim, M.A.; Ross, S.A. Phytochemical study of Piliostigmathonningii, a medicinal plant grown in Nigeria. Med. Chem. Res. 2018, 27, 2325–2350. [Google Scholar] [CrossRef]
- Venkatesh, S.; Reddy, Y.S.R.; Ramesh, M.; Swamy, M.M.; Mahadevan, N.; Suresh, B. Pharmacognostical studies on Dodonaea viscosa leaves. Afr. J. Pharm. Pharmacol. 2008, 2, 083–088. [Google Scholar]
- Mussard, E.; Cesaro, A.; Lespessailles, E.; Legrain, B.; Berteina-Raboin, S.; Toumi, H. Andrographolide, a natural antioxidant: An update. Antioxidants 2019, 8, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Ghefreh, A.A.; Canatan, H.; Ezeamuzie, C.I. In vitro and in vivo anti-inflammatory effects of andrographolide. Int. Immunopharmacol. 2009, 9, 313–318. [Google Scholar] [CrossRef]
- Chan, S.J.; Wong, W.S.F.; Wong, P.T.H.; Bian, J.-S. Neuroprotective effects of andrographolide in a rat model of permanent cerebral ischemia: Effect of andrographolide on cerebral ischaemic damage. Br. J. Pharmacol. 2010, 161, 668–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa, S.S.; Sharma, A. Hepatoprotective activity of andrographolide from Andrographis paniculata against carbon tetrachloride. Indian J. Med. Res. 1990, 92, 276–283. [Google Scholar]
- Wintachai, P.; Kaur, P.; Lee, R.C.H.; Ramphan, S.; Kuadkitkan, A.; Wikan, N.; Ubol, S.; Roytrakul, S.; Chu, J.J.H.; Smith, D.R. Activity of andrographolide against chikungunya virus infection. Sci. Rep. 2015, 5, 14179. [Google Scholar] [CrossRef] [Green Version]
- Manjula, S.; Kalaiarasi, C.; Pavan, M.S.; Hathwar, V.R.; Kumaradhas, P. Charge density and electrostatic potential of hepatitis C anti-viral agent andrographolide: An experimental and theoretical study. Acta Crystallogr. B Struct. Sci. Cryst. Eng. Mater. 2018, 74, 693–704. [Google Scholar] [CrossRef]
- Paemanee, A.; Hitakarun, A.; Wintachai, P.; Roytrakul, S.; Smith, D.R. A proteomic analysis of the anti-dengue virus activity of andrographolide. Biomed. Pharmacother. 2019, 109, 322–332. [Google Scholar] [CrossRef]
- Mussbacher, M.; Salzmann, M.; Brostjan, C.; Hoesel, B.; Schoergenhofer, C.; Datler, H.; Hohensinner, P.; Basílio, J.; Petzelbauer, P.; Assinger, A.; et al. Cell type-specific roles of NF-κB linking inflammation and thrombosis. Front. Immunol. 2019, 10, 85. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.-C.; Chou, Y.-J.; Lin, C.-C.; Liu, S.-H.; Oswita, A.; Huang, Y.-L.; Wang, Y.-L.; Syu, J.-L.; Sun, C.-M.; Leu, C.-M.; et al. Andrographolide and its potent derivative exhibit anticancer effects against imatinib-resistant chronic myeloid leukemia cells by downregulating the Bcr-Abl oncoprotein. Biochem. Pharmacol. 2019, 163, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q. Triptolide and its expanding multiple pharmacological functions. Int. Immunopharmacol. 2011, 11, 377–383. [Google Scholar] [CrossRef]
- Noel, P.; Von Hoff, D.D.; Saluja, A.K.; Velagapudi, M.; Borazanci, E.; Han, H. Triptolide and its derivatives as cancer therapies. Trends Pharmacol. Sci. 2019, 40, 327–341. [Google Scholar] [CrossRef]
- Kamikawa, S.; Oshimo, S.; Ohta, E.; Nehira, T.; Ômura, H.; Ohta, S. Cassane diterpenoids from the roots of Caesalpiniadecapetala var. japonica and structure revision of caesaljapin. Phytochemistry 2016, 121, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, S.M.; Malmstrom, R.D.; Russo, A.; Mueller, N.; Pang, Y.-P.; Watowich, S.J. Structure-based discovery of dengue virus protease inhibitors. Antivir. Res. 2009, 82, 110–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constant, D.A.; Mateo, R.; Nagamine, C.M.; Kirkegaard, K. Targeting intramolecular proteinase NS2B/3 cleavages for trans-dominant inhibition of dengue virus. Proc. Natl. Acad. Sci. USA 2018, 115, 10136–10141. [Google Scholar] [CrossRef] [Green Version]
- Rogée, S.; Larrous, F.; Jochmans, D.; Ben-Khalifa, Y.; Neyts, J.; Bourhy, H. Pyrimethamine inhibits rabies virus replication in vitro. Antivir. Res. 2019, 161, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Moshiri, J.; Constant, D.A.; Liu, B.; Mateo, R.; Kearnes, S.; Novick, P.; Prasad, R.; Nagamine, C.; Pande, V.; Kirkegaard, K. A targeted computational screen of the SWEETLEAD database reveals FDA-approved compounds with anti-dengue viral activity. mBio 2020, 11. [Google Scholar] [CrossRef]
- Qiu, J. Traditional medicine: A culture in the balance: Traditional medicine. Nature 2007, 448, 126–128. [Google Scholar] [CrossRef] [PubMed]
- Al-Khafaji, K.; TaskinTok, T. Molecular dynamics simulation, free energy landscape and binding free energy computations in exploration the anti-invasive activity of amygdale in against metastasis. Comput. Methods Progr. Biomed. 2020, 195, 105660. [Google Scholar] [CrossRef]
- Modis, Y.; Ogata, S.; Clements, D.; Harrison, S.C. A ligand-binding pocket in the dengue virus envelope glycoprotein. Proc. Natl. Acad. Sci. USA 2003, 100, 6986–6991. [Google Scholar] [CrossRef] [Green Version]
- Luo, D.; Xu, T.; Hunke, C.; Gruber, G.; Vasudevan, S.G.; Lescar, J. Crystal structure of the NS3 protease-helicase from dengue virus. Acta Crystallogr. A 2008, 82, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Soh, T.S.; Zheng, J.; Chan, K.W.K.; Phoo, W.W.; Lee, C.C.; Tay, M.Y.F.; Swaminathan, K.; Cornvik, T.C.; Lim, S.P.; et al. A crystal structure of the Dengue virus NS5 protein reveals a novel inter-domain interface essential for protein flexibility and virus replication. PLoS Pathog. 2015, 11, e1004682. [Google Scholar] [CrossRef]
- Scaturro, P.; Cortese, M.; Chatel-Chaix, L.; Fischl, W.; Bartenschlager, R. Dengue virus non-structural protein 1 modulates infectious particle production via interaction with the structural proteins. PLoS Pathog. 2015, 11, e1005277. [Google Scholar] [CrossRef]
- Mooers, B.H.M. A PyMOL snippet library for Jupyter to boost researcher productivity. Comput. Sci. Eng. 2021, 23, 47–53. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Thornton, J.M. PROCHECK: Validation of protein-structure coordinates. In International Tables for Crystallography; International Union of Crystallography: Chester, UK, 2012; ISBN 9780470685754. [Google Scholar]
- Yuan, S.; Chan, H.C.S.; Hu, Z. Using PyMOL as a platform for computational drug design. Comput. Mol. Sci. 2017, 7, e1298. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [PubMed]
- Binkowski, T.A.; Naghibzadeh, S.; Liang, J. CASTp: Computed Atlas of Surface Topography of proteins. Nucleic Acids Res. 2003, 31, 3352–3355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, J.R. Diterpenoids. Nat. Prod. Rep. 2009, 26, 1156–1171. [Google Scholar] [CrossRef]
- Wang, G.-C.; Li, J.-G.; Li, G.-Q.; Xu, J.-J.; Wu, X.; Ye, W.-C.; Li, Y.-L. Clerodane diterpenoids from Crotoncras sifolius. J. Nat. Prod. 2012, 75, 2188–2192. [Google Scholar] [CrossRef] [PubMed]
- Olivon, F.; Palenzuela, H.; Girard-Valenciennes, E.; Neyts, J.; Pannecouque, C.; Roussi, F.; Grondin, I.; Leyssen, P.; Litaudon, M. Antiviral activity of flexibilane and tigliane diterpenoids from Stillingialineata. J. Nat. Prod. 2015, 78, 1119–1128. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Ali, M.T.; Shawan, M.M.A.K.; Sarwar, M.G.; Khan, M.A.K.; Halim, M.A. Halogen-directed drug design for Alzheimer’s disease: A combined density functional and molecular docking study. Springerplus 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Spessard, G.O. ACD Labs/LogP dB 3.5 and ChemSketch 3.5. J. Chem. Inf. Comput. Sci. 1998, 38, 1250–1253. [Google Scholar] [CrossRef]
- Lipinski, C.A. Lead-and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef]
- Ghose, A.K.; Herbertz, T.; Salvino, J.M.; Mallamo, J.P. Knowledge-based chemoinformatic approaches to drug discovery. Drug Discov. Today 2006, 11, 1107–1114. [Google Scholar] [CrossRef]
- Sneader, W.A.L.T.E.R. Chronology of drug introductions. Compr. Med. Chem. 1990, 1, 7–80. [Google Scholar]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef]
- Averbukh, I.; Ben-Zvi, D.; Mishra, S.; Barkai, N. Scaling morphogen gradients during tissue growth by a cell division rule. Development 2014, 141, 2150–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, D.E. In silico prediction of blood-brain barrier permeation. Drug Discov. Today 2003, 8, 927–933. [Google Scholar] [CrossRef]
- Bickerton, G.R.; Paolini, G.V.; Besnard, J.; Muresan, S.; Hopkins, A.L. Quantifying the chemical beauty of drugs. Nat. Chem. 2012, 4, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Leeson, P.D.; Davis, A.M.; Steele, J. Drug-like properties: Guiding principles for design–or chemical prejudice? Drug Discov. Today Technol. 2004, 1, 189–195. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Pires, D.E.V.; Blundell, T.L.; Ascher, D.B. PkCSM: Predicting small-molecule pharmacokinetic and toxicity properties using graph-based signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Feng, N.; Huang, J.; Wang, M.; Zhang, L.; Yu, J.; Dai, X.; Cao, J.; Huang, G. Incorporation of amino moiety to alepterolic acid improve activity against cancer cell lines: Synthesis and biological evaluation. Bioorg. Chem. 2020, 98, 103756. [Google Scholar] [CrossRef] [PubMed]
- Kar, S.; Roy, K. How far can virtual screening take us in drug discovery? Expert Opin. Drug Discov. 2013, 8, 245–261. [Google Scholar] [CrossRef] [PubMed]
- Bekker, G.-J.; Kamiya, N. Dynamic docking using multicanonical molecular dynamics: Simulating complex formation at the atomistic level. Methods Mol. Biol. 2021, 2266, 187–202. [Google Scholar] [PubMed]
- Berendsen, H.J.C.; van der Spoel, D.; van Drunen, R. GROMACS: A message-passing parallel molecular dynamics implementation. Comput. Phys. Commun. 1995, 91, 43–56. [Google Scholar] [CrossRef]
- Sapay, N.; Tieleman, D.P. Combination of the CHARMM27 force field with united-atom lipid force fields. J. Comput. Chem. 2011, 32, 1400–1410. [Google Scholar] [CrossRef]
- Al-Khafaji, K.; Tok, T.T. Understanding the mechanism of amygdalin’s multifunctional anti-cancer action using computational approach. J. Biomol. Struct. Dyn. 2021, 39, 1600–1610. [Google Scholar] [CrossRef]
- Kumari, R.; Kumar, R.; Open Source Drug Discovery Consortium; Lynn, A. g_mmpbsa—AGROMACS tool for high–throughput MM-PBSA calculations. J. Chem. Inf. Model. 2014, 54, 1951–1962. [Google Scholar] [CrossRef]
- Lagunin, A.; Ivanov, S.; Rudik, A.; Filimonov, D.; Poroikov, V. DIGEP-Pred: Web service for in silico prediction of drug-induced gene expression profiles based on structural formula. Bioinformatics 2013, 29, 2062–2063. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Triptolide | Stevioside | AlepterolicAcid | Sphaeropsidin A | Methyl DodovisateA | Andrographolide | Caesalacetal | Pyrimethamine |
|---|---|---|---|---|---|---|---|---|
| Formula | C20H24O6 | C38H60O18 | C20H32O3 | C20H26O5 | C22H28O2 | C20H30O5 | C21H28O5 | C12H13ClN4 |
| Molecular weight (g/mol) | 360.40 | 804.87 | 320.47 | 346.42 | 324.46 | 350.45 | 360.44 | 248.71g/mol |
| H-Bond Acceptors | 6 | 18 | 3 | 5 | 2 | 5 | 5 | 2 |
| H-Bond Donors | 1 | 11 | 2 | 2 | 0 | 3 | 1 | 2 |
| Num. Rotatable Bonds | 1 | 10 | 4 | 1 | 5 | 3 | 2 | 2 |
| TPSA (Å2) | 84.12 | 294.98 | 57.53 | 83.83 | 26.30 | 86.99 | 68.90 | 77.82 |
| Fraction Csp3 | 0.85 | 0.92 | 0.75 | 0.70 | 0.50 | 0.75 | 0.76 | 0.17 |
| Molar Refractivity | 88.54 | 188.26 | 95.49 | 91.95 | 100.18 | 95.21 | 95.30 | 71.06 |
| LogPo/w (XLOGP3) | 0.22 | −1.20 | 4.74 | 2.64 | 5.31 | 2.16 | 3.08 | 2.69 |
| LogS (ESOL) | −2.15 | −3.41 | −4.55 | −3.58 | −4.87 | −3.18 | −4.03 | −3.47 |
| Max. tolerated dose (human) (logmg/kg/day) | −0.321 | −1.524 | −0.297 | −0.074 | −0.159 | 0.128 | −0.14 | 0.113 |
| Oral Rat Acute Toxicity (LD50; mol/kg) | 3.107 | 2.597 | 2.28 | 1.92 | 1.779 | 2.162 | 2.581 | 2.912 |
| Hepatotoxicity | No | No | No | No | No | No | Yes | No |
| Minnow toxicity (logmM) | 1.983 | 9.202 | 0.459 | 1.606 | −0.525 | 1.37 | 0.418 | 0.919 |
| Blood brain barrier (logBB) | −0.362 | −2.029 | −0.018 | 0.016 | 0.629 | −0.598 | −0.163 | 0.278 |
| HIA (%) | 83.195 | 0 | 94.672 | 95.753 | 97.808 | 95.357 | 97.564 | 92.738 |
| CaCo2 Permeability | 0.401 | −1.087 | 1.432 | 1.135 | 1.64 | 1.07 | 1.145 | 0.927 |
| Total Clearance (logml/min/kg) | 0.484 | 0.691 | 1.122 | 0.541 | 1.381 | 1.183 | 0.538 | −0.033 |
| Compounds | Target | Interact Residues | No. of H-Bond | H-Bond Residues | H-Bond Length | Binding Energy (kcal/mol) |
|---|---|---|---|---|---|---|
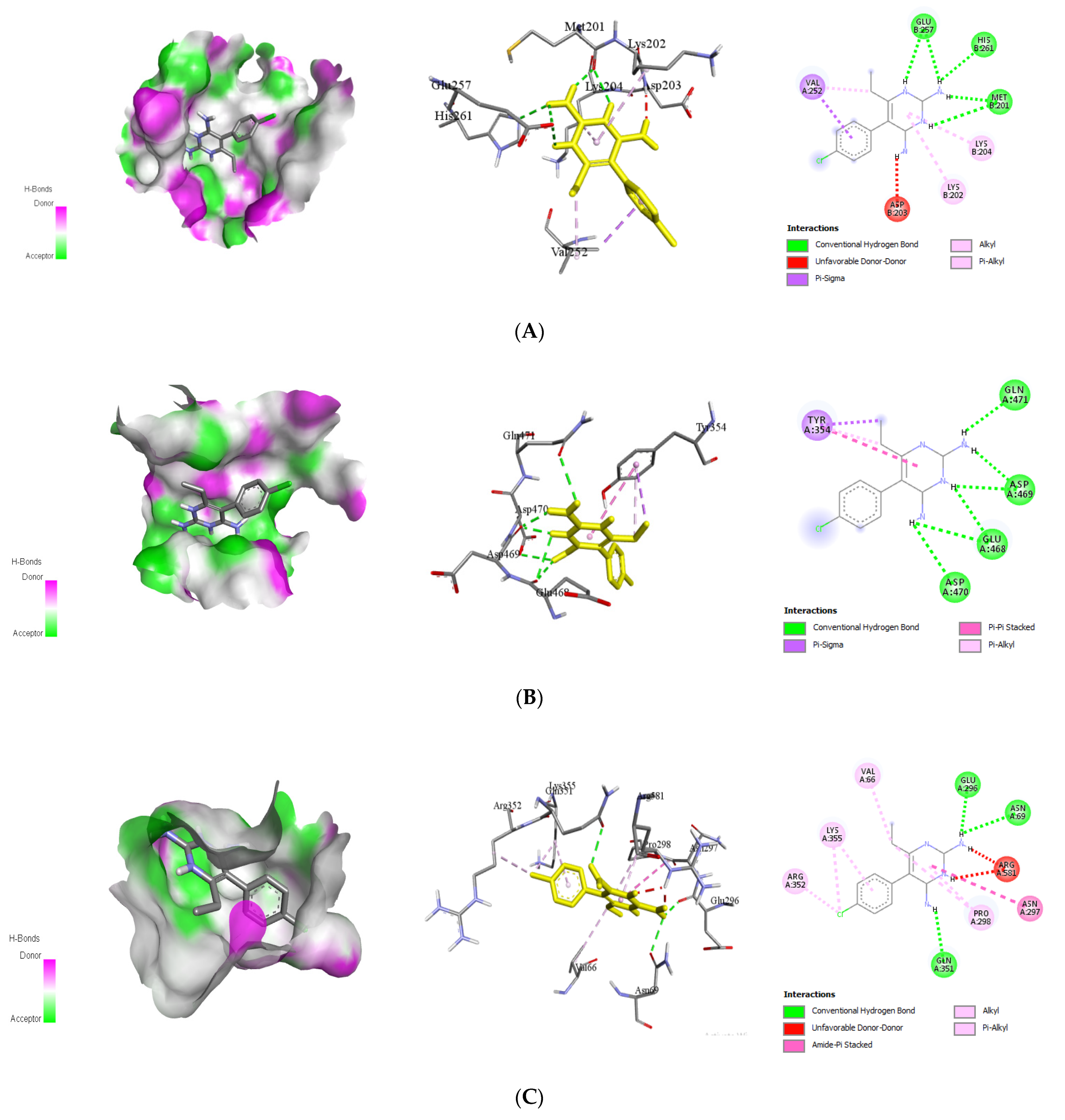
| Triptolide | 1OKE | Leu253 Thr236 Thr262 | 1 | Thr265 | 1.76 | −8.1 |
| Stevioside | Ala259 Ala263 Trp212 | 2 | Gln256 Hios209 | 2.09 2.16 | −8.4 | |
| Alepterolic acid | Leu253 Pro217 | 2 | Gln256 Thr265 | 2.31 1.87 | −8.3 | |
| Sphaeropsidin A | His261 Thr265 Trp206 | 0 | - | - | −8.7 |
| Compounds | Target | Interact Residues | No. of H-Bond | H-Bond Residue | H-Bond Length | Binding Energy (kcal/mol) |
|---|---|---|---|---|---|---|
| Stevioside | Asp192 Ile203 Met191 His194 | 7 | Arg202 Asn175 Glu173 Glu177 Gly198 His194 Pro174 | 2.33 2.29 1.96 2.59 2.17 2.39 2.60 | −8.0 | |
| Sphaeropsidin A | 2VBC | Ala197 Ile203 Leu193 | 1 | Asp175 | 2.57 | −8.3 |
| Methyldodovisate A | Asp258 Arg215 Arg217 His251 Ile256 | 3 | Arg254 Gly253 Thr252 | 2.17 2.17 2.94 | −9.2 | |
| Caesalacetal | Ala197 His194 Leu193 | 1 | Asp175 | 2.59 | −8.0 |
| Compounds | Target | Interact Residues | No. of H-Bond | H-Bond Residues | H-Bond Length | Binding Energy (kcal/mol) |
|---|---|---|---|---|---|---|
| Triptolide | Glu356 Gly258 His52 Tyr119 Val687 | 2 | Ala259 Arg540 | 2.28 3.10 | −8.8 | |
| Stevioside | 4V0Q | Asp663 Ile797 | 3 | Lys401 Ser661 Ser710 | 2.23 2.83 2.53 | −9.4 |
| Andrographolide | Ile797 Val603 | 4 | Asp663 Gly604 Thr605 Tyr606 | 2.25 2.39 2.56 1.82 | −8.4 | |
| Caesalacetal | Cys82 Gly148 Ile147 Trp87 | 4 | Arg84 Asp146 Gly85 Gly86 | 2.63 2.36 2.29 2.25 | −8.4 |
| Compounds | Target | Interacting Residues | No. of H-Bond | H-Bond Residue | Bond Length (A) | Binding Energy (Kcal/mol) |
|---|---|---|---|---|---|---|
| Triptolide | Lys174 Phe178 Pro226 | 3 | Lys227 Ser181 Ser227 | 2.28 2.52 2.69 | −8.3 | |
| Stevioside | Lys174 Lys227 Phe178 Pro226 | 5 | Asn234 Asp176 Glu154 Ser181 Trp232 | 2.72 2.41 1.99 2.14 2.06 | −9.3 | |
| Sphaeropsidin A | 4O6B | Lys172 Lys227 Phe178 Pro226 Trp232 | 2 | Asp176 Ser181 | 2.21 2.16 | −8.5 |
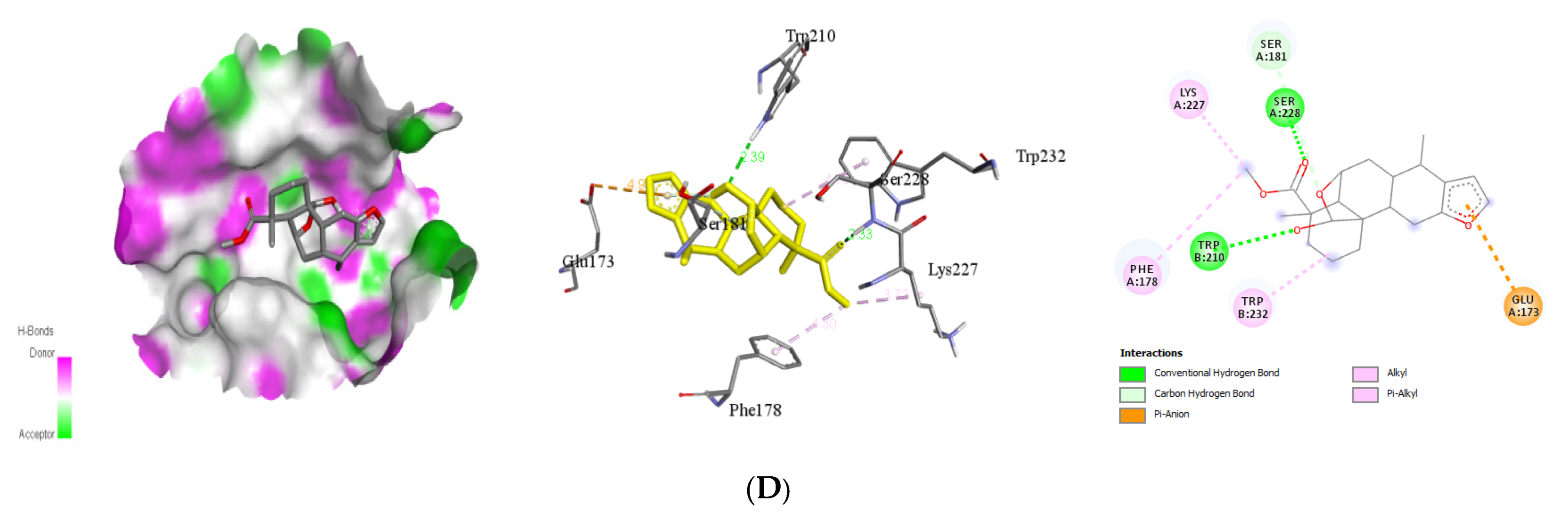
| Caesalacetal | Glu173 Lys227 Phe178 Ser181 Trp232 | 2 | Ser228 Trp210 | 2.33 2.39 | −8.5 |
| Compounds | Target | Interacting Residues | No. of H-Bond | H-Bond Residue | Bond Length (Å) | Binding Energy (kcal/mol) |
|---|---|---|---|---|---|---|
| Pyrimethamine | E protein (1OKE) | Asp203 Lys202 Lys204 Val252 | 5 | Glu257 His261 Met201 | 2.55 2.60 2.44 | −7.5 |
| NS3(2VBC) | Tyr354 | 6 | Asp469 Asp470 Gln471 Glu468 | 2.15 2.83 2.56 2.63 | −6.3 | |
| NS5(4V0Q) | Arg352, Arg581, Asn297, Lys355, Pro298, Val66 | 3 | Glu296 Asn69 Glu351 | 2.04 2.60 2.55 | −7.8 | |
| NS1(4O6B) | Phe178 Ser181 | 3 | Asp176 Asp180 Cys179 | 2.32 2.42 2.43 | −6.6 |
| Target Name | Compound Name | Vander Waals Energy (kJ/mol) | Electrostatic Energy (kJ/mol) | Polar Solvation Energy (kJ/mol) | SASA Energy (kJ/mol) | Binding Energy (kJ/mol) |
|---|---|---|---|---|---|---|
| 1OKE | Native | −96.764 | 314.382 | −253.723 | 32.344 | −3.761 |
| Alepterolicacid | −88.371 | −14.159 | 68.7155 | −12.475 | −46.2895 | |
| SphaeropsidinA | −122.068 | −5.756 | 71.871 | −14.922 | −70.875 | |
| Stevioside | −83.430 | −12.410 | 73.405 | −11.170 | −33.605 | |
| Triptolide | −130.551 | −13.742 | 66.847 | −13.472 | −90.918 | |
| 2VBC | Native | −149.888 | −15.821 | 87.180 | −16.847 | −94.556 |
| Caesalacetal | −148.616 | −24.435 | 104.966 | −16.718 | −84.309 | |
| MethyldodovisateA | −138.175 | −14.924 | 118.165 | −14.918 | −49.852 | |
| SphaeropsidinA | −115.183 | 3.982 | 43.248 | −13.188 | −81.141 | |
| Stevioside | −195.236 | −22.923 | 144.028 | −22.388 | −96.519 | |
| 4O6B | Native | −60.911 | −32.869 | 89.094 | −9.556 | −14.602 |
| Caesalacetal | −139.283 | −33.117 | 81.796 | −13.614 | −104.281 | |
| SphaeropsidinA | −90.141 | −8.125 | 43.680 | −9.868 | −64.454 | |
| Stevioside | −162.844 | −110.614 | 151.263 | −22.976 | −145.171 | |
| Triptolide | −116.724 | −75.279 | 100.616 | −13.566 | −104.953 | |
| 4V0Q | Native | −135.681 | −41.363 | 131.823 | −14.601 | −59.822 |
| Andrographolide | −0.036 | 4.881 | 58.619 | −2.893 | 60.571 | |
| Caesalacetal | −108.493 | 3.610 | 35.992 | −12.449 | −81.34 | |
| Stevioside | −270.746 | −68.928 | 236.892 | −28.205 | −130.987 | |
| Triptolide | −178.301 | −24.621 | 210.824 | −19.300 | −11.398 |
| Compounds | Source | PubChem ID | Chemical Structure |
|---|---|---|---|
| Phorbol ester | Jatropha curcas | 22833501 |  |
| Triptolide | Tripterygium wilfordii | 107985 |  |
| Steviol | Stevia rebaudiana | 452967 |  |
| Ferruginol | Prumnopitys andina | 442027 |  |
| 18-oxoferruginol | Torreya nucifera | 52946772 |  |
| Andrographolide | Andrographis paniculata | 5318517 |  |
| Stevioside | Stevia rebaudiana | 442089 |  |
| Rebaudioside A | Stevia rebaudiana Bertoni | 6918840 | 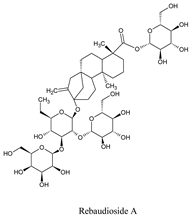 |
| Forskolin | Coleus forskohli | 47936 |  |
| Ent-kaur-16-en-19-oic acid | Elaeoselinum foetidum | 73062 |  |
| 9(11),16-kauradien-19-oic acid | Melantheria albinervia | 14635430 |  |
| Alepterolic acid | Copaifera reticulata | 13858188 |  |
| Zanzibaric acid | Hymenaea courbaril | 101289556 |  |
| Isoozic acid | Hymenaea courbaril | 100983062 |  |
| Sphaeropsidin A | Diplodia cupressi | 51361447 |  |
| Phytol | Hierochloëo dorata | 5280435 |  |
| 7-oxo-8,11,13-cleistanthatrien-3-ol | V. gigantea | 132609177 |  |
| Methyl dodovisate A | Hierochlo ëodorata | 146156767 | 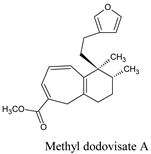 |
| Caesalacetal | Caesalpinia decapetala var. | 132918611 |  |
| Caesaljapin | Caesalpinia decapetala var. | 6712179 | 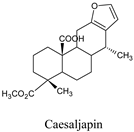 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, R.A.; Hossain, R.; Siyadatpanah, A.; Al-Khafaji, K.; Khalipha, A.B.R.; Dey, D.; Asha, U.H.; Biswas, P.; Saikat, A.S.M.; Chenari, H.A.; et al. Diterpenes/Diterpenoids and Their Derivatives as Potential Bioactive Leads against Dengue Virus: A Computational and Network Pharmacology Study. Molecules 2021, 26, 6821. https://doi.org/10.3390/molecules26226821
Khan RA, Hossain R, Siyadatpanah A, Al-Khafaji K, Khalipha ABR, Dey D, Asha UH, Biswas P, Saikat ASM, Chenari HA, et al. Diterpenes/Diterpenoids and Their Derivatives as Potential Bioactive Leads against Dengue Virus: A Computational and Network Pharmacology Study. Molecules. 2021; 26(22):6821. https://doi.org/10.3390/molecules26226821
Chicago/Turabian StyleKhan, Rasel Ahmed, Rajib Hossain, Abolghasem Siyadatpanah, Khattab Al-Khafaji, Abul Bashar Ripon Khalipha, Dipta Dey, Umma Hafsa Asha, Partha Biswas, Abu Saim Mohammad Saikat, Hadi Ahmadi Chenari, and et al. 2021. "Diterpenes/Diterpenoids and Their Derivatives as Potential Bioactive Leads against Dengue Virus: A Computational and Network Pharmacology Study" Molecules 26, no. 22: 6821. https://doi.org/10.3390/molecules26226821