
Altitudinal Variation of Metabolites, Mineral Elements and Antioxidant Activities of Rhodiola crenulata (Hook.f. & Thomson) H.Ohba

Abstract
:1. Introduction
2. Results and Discussion
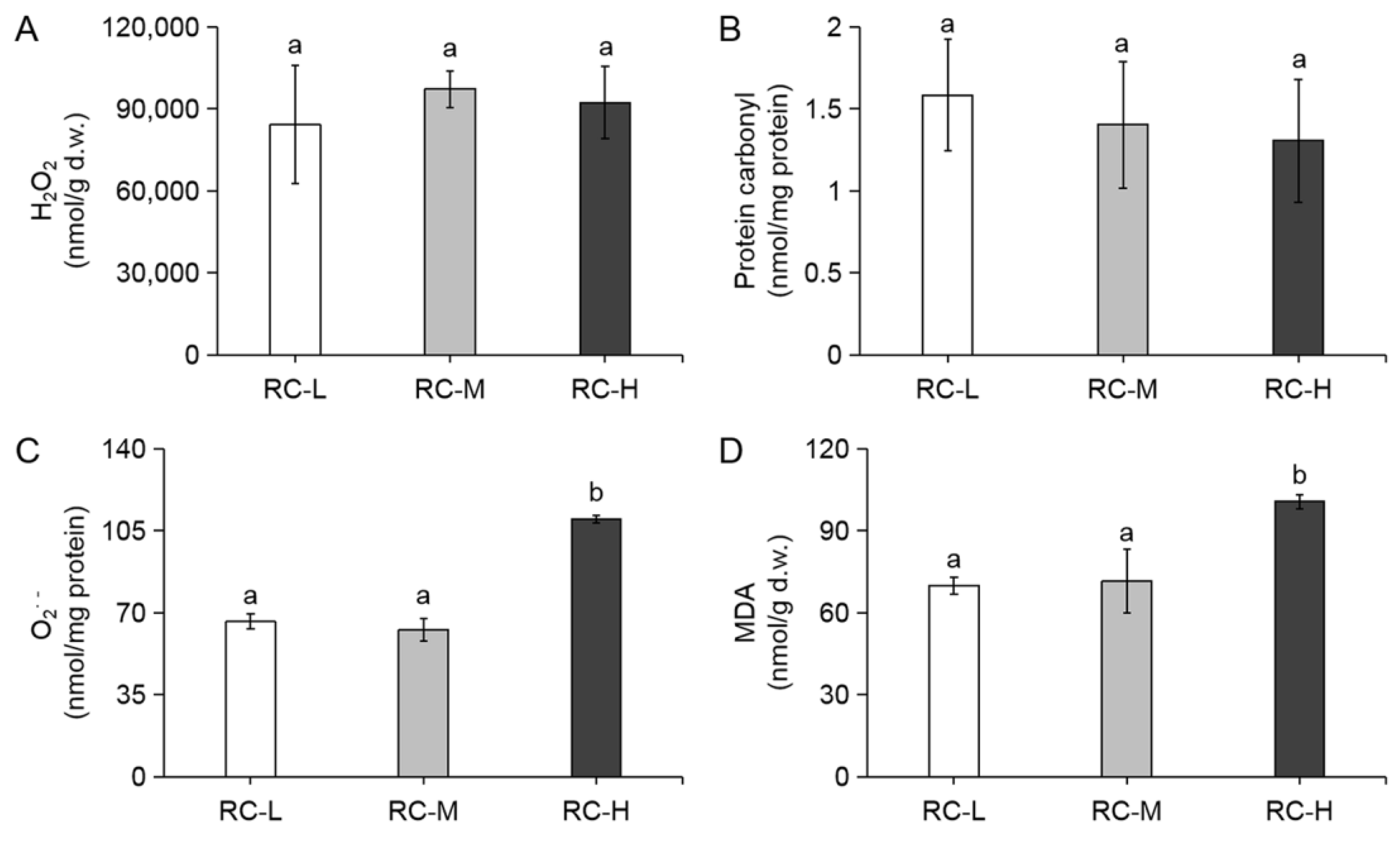
2.1. Effect of Altitude Gradient on Oxidative Stress Levels in Rhodiola crenulata
2.2. Effect of Altitude Gradient on the Accumulation of Seven Mineral Elements in Rhodiola crenulata
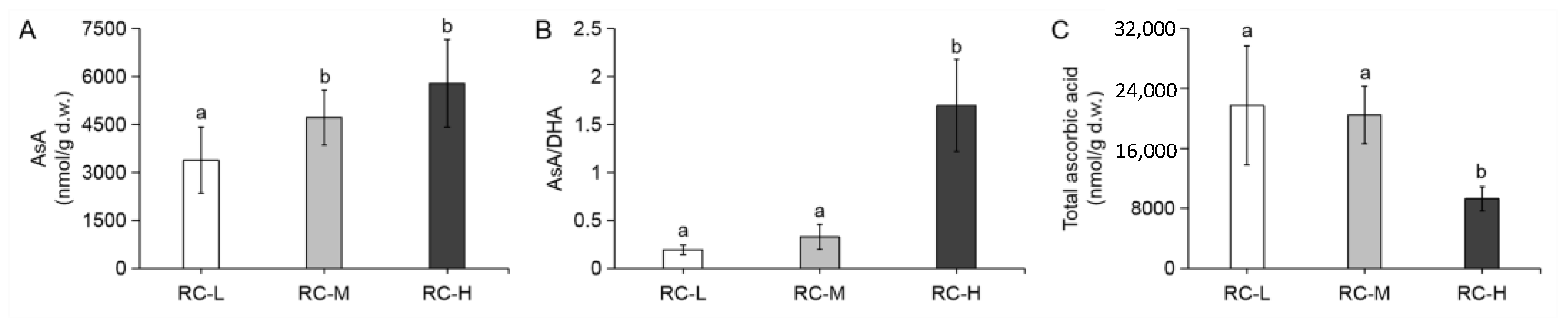
2.3. Effects of Altitude Gradient on Phenolic Components Content, Ascorbic Acid Content and Antioxidant Capacity in Rhodiola crenulata
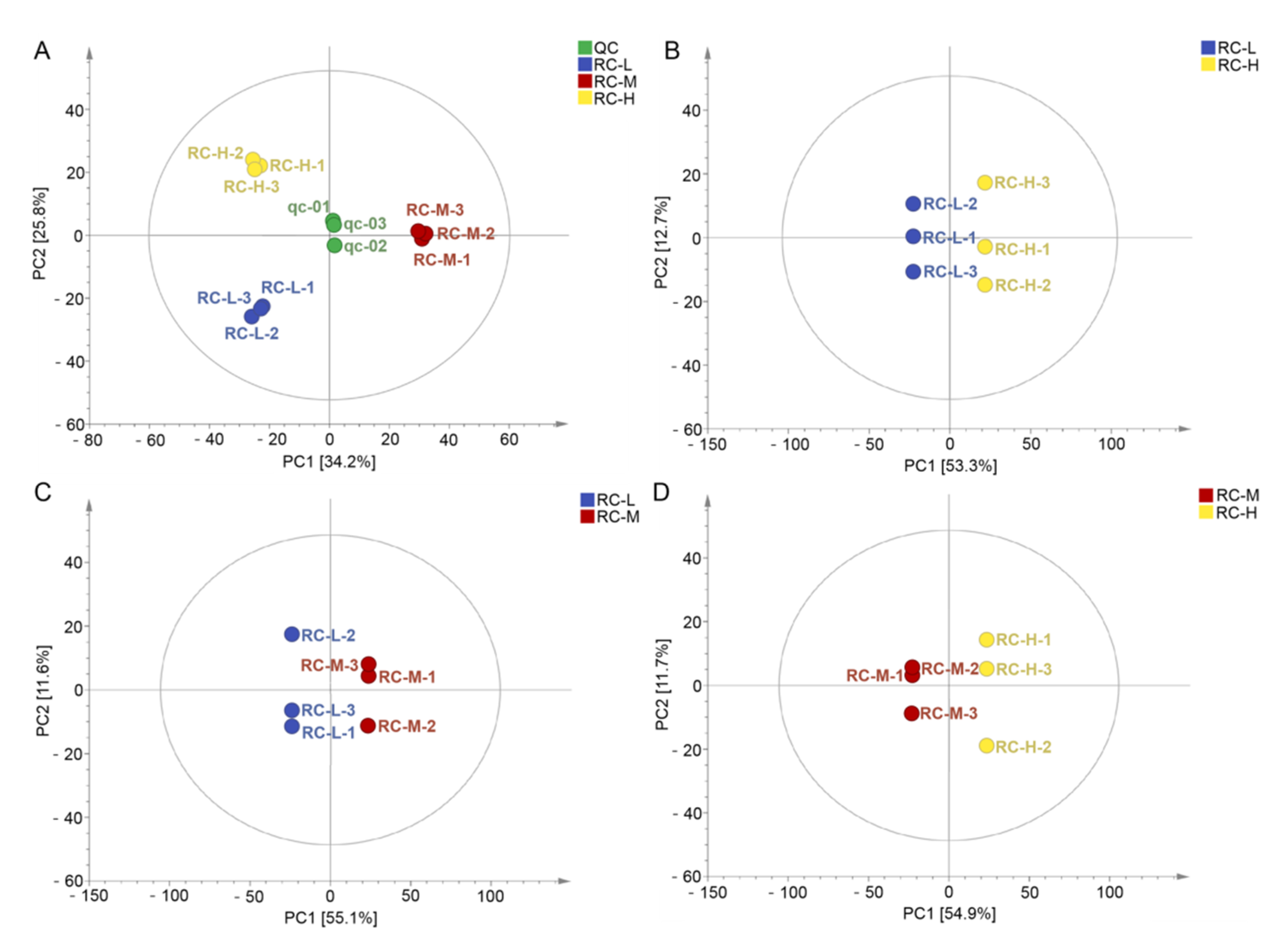
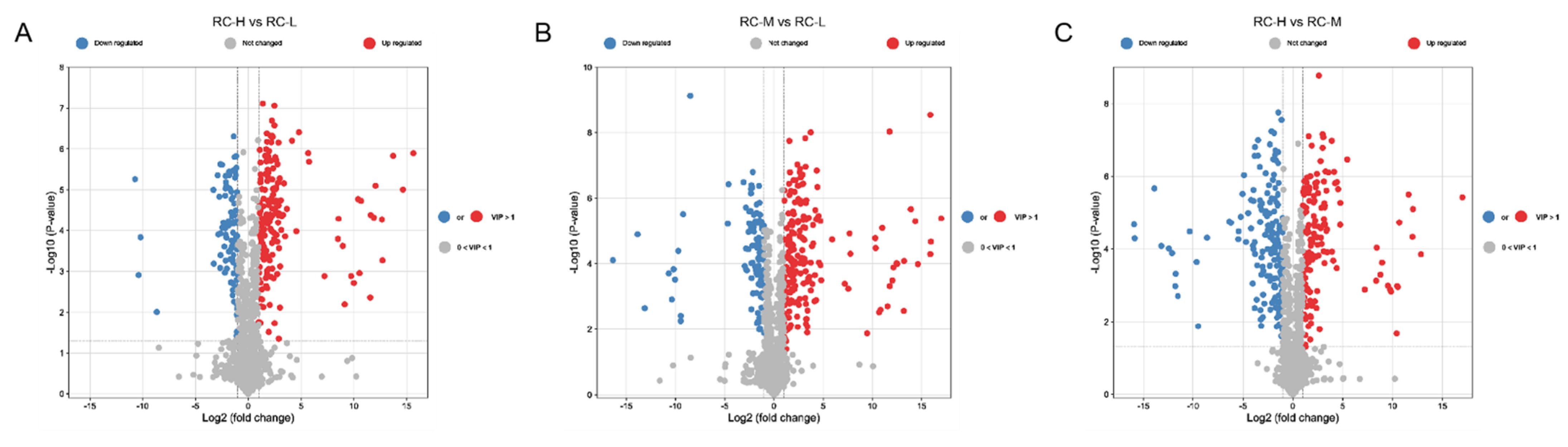
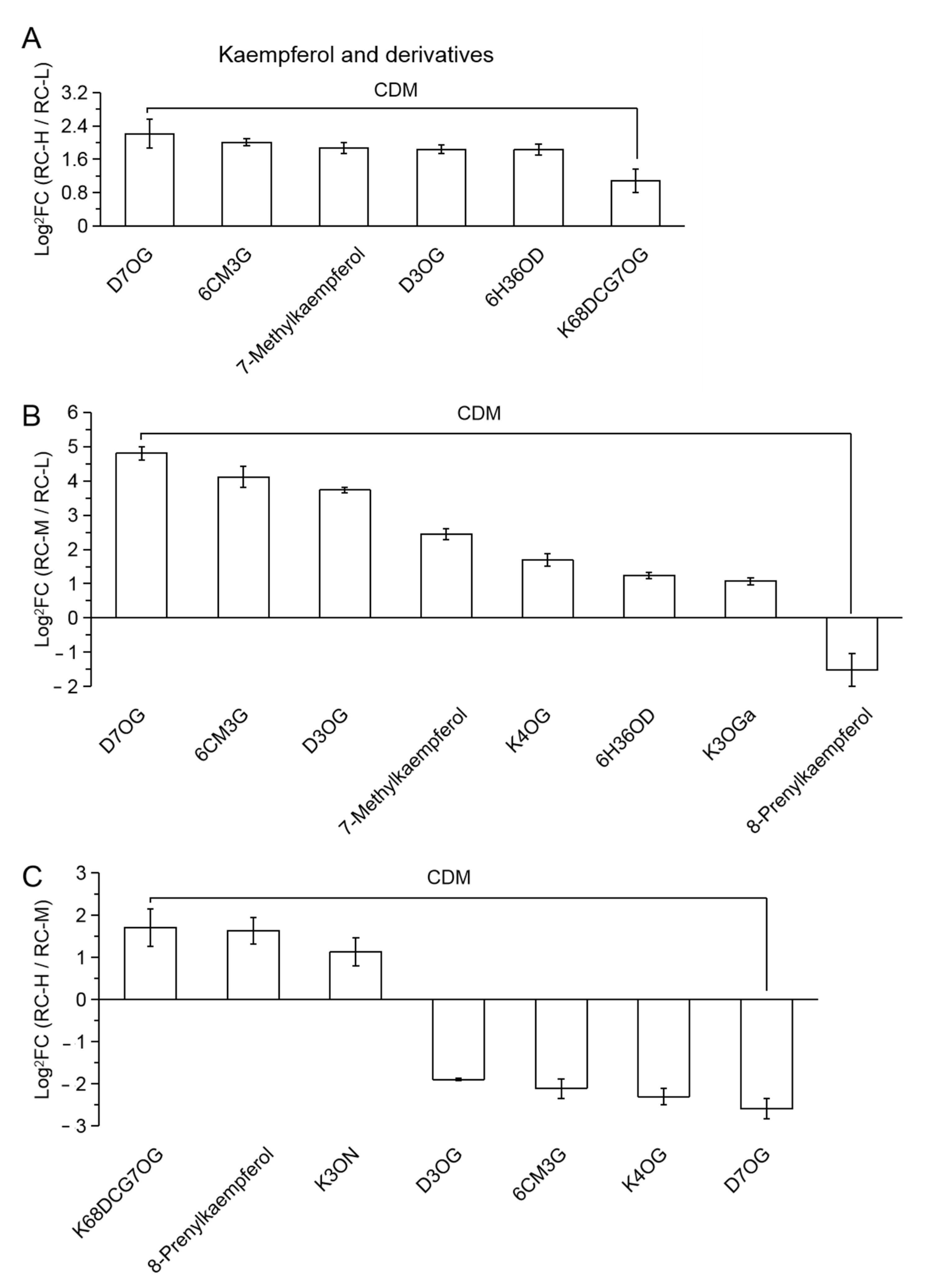
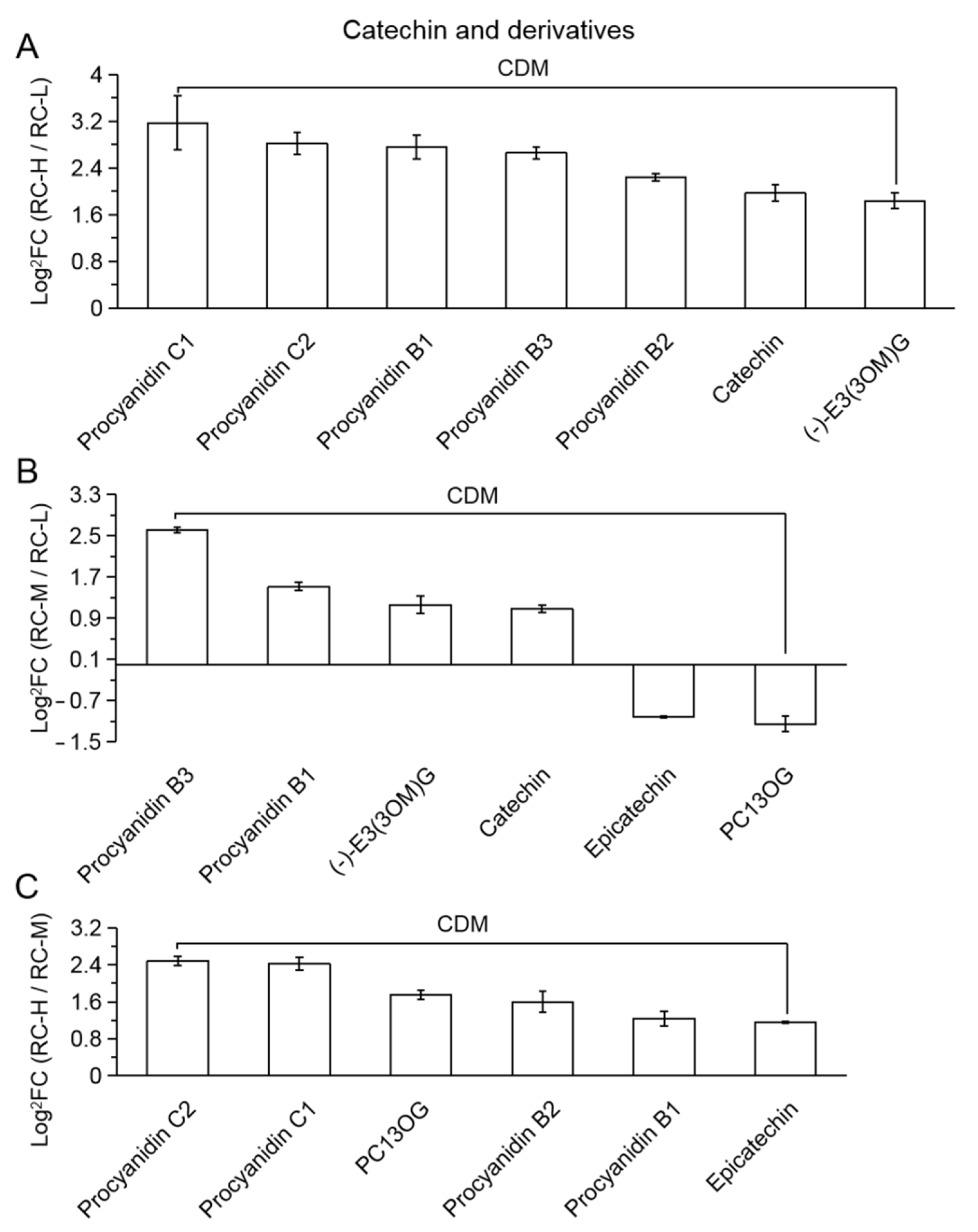
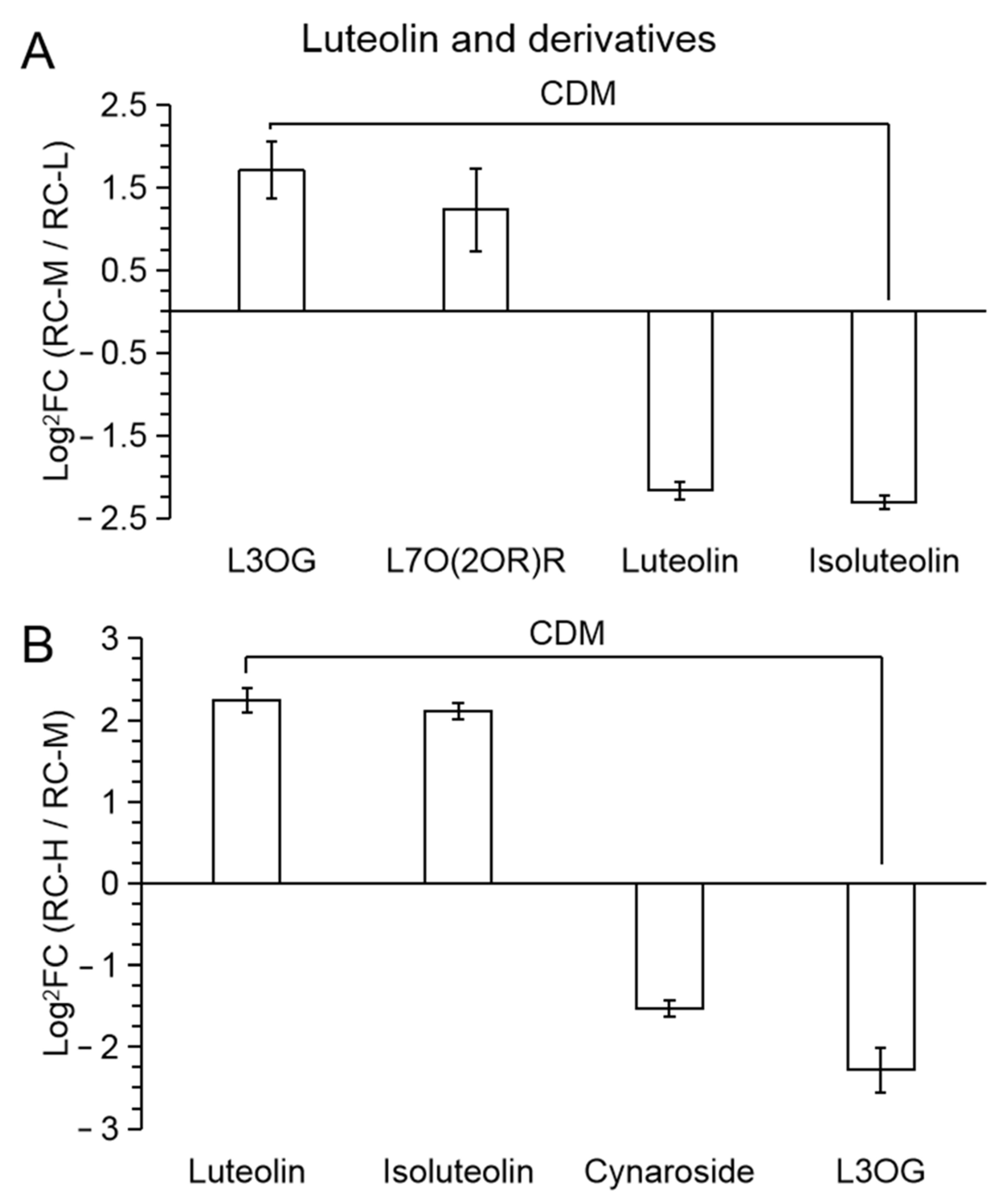
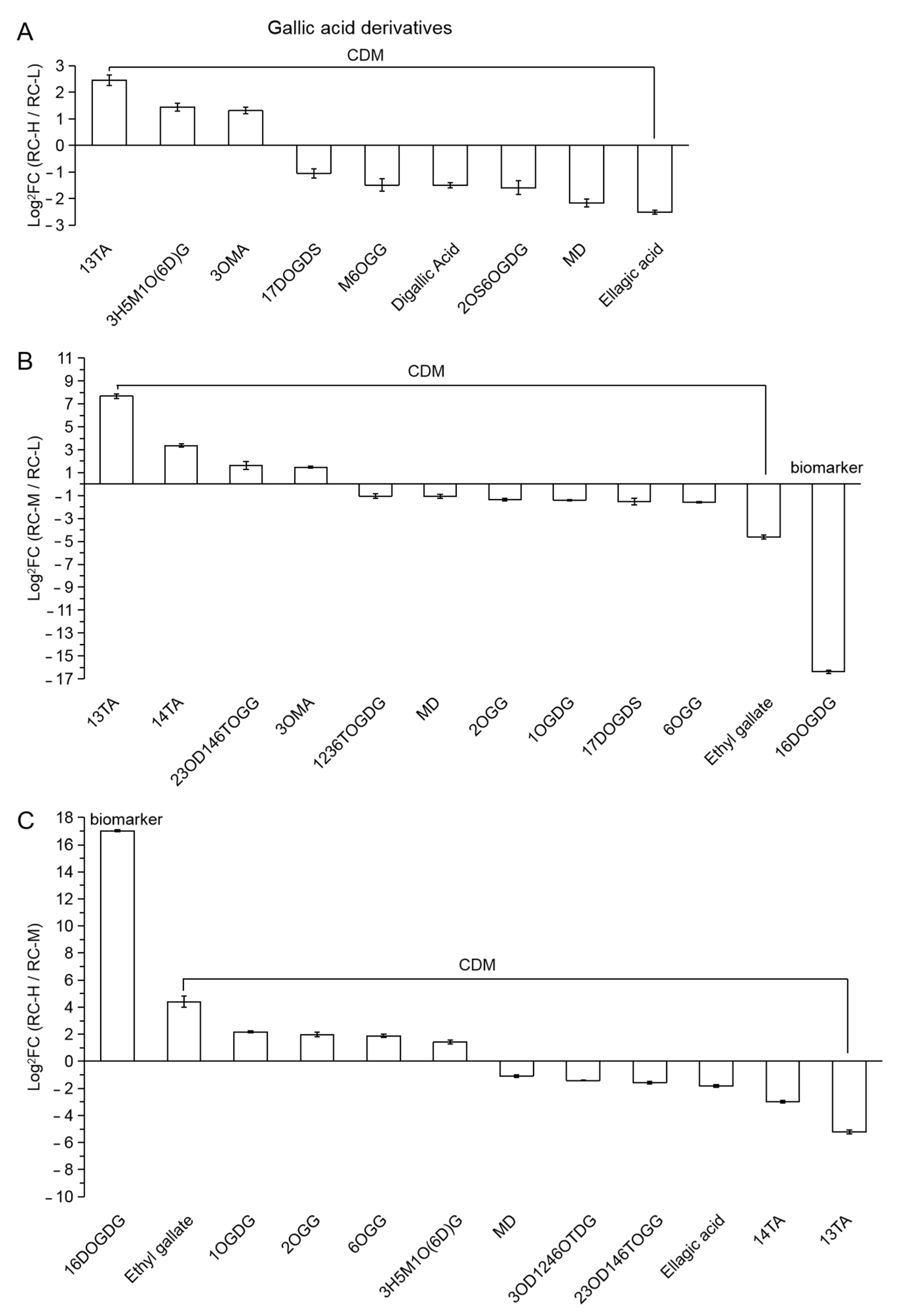
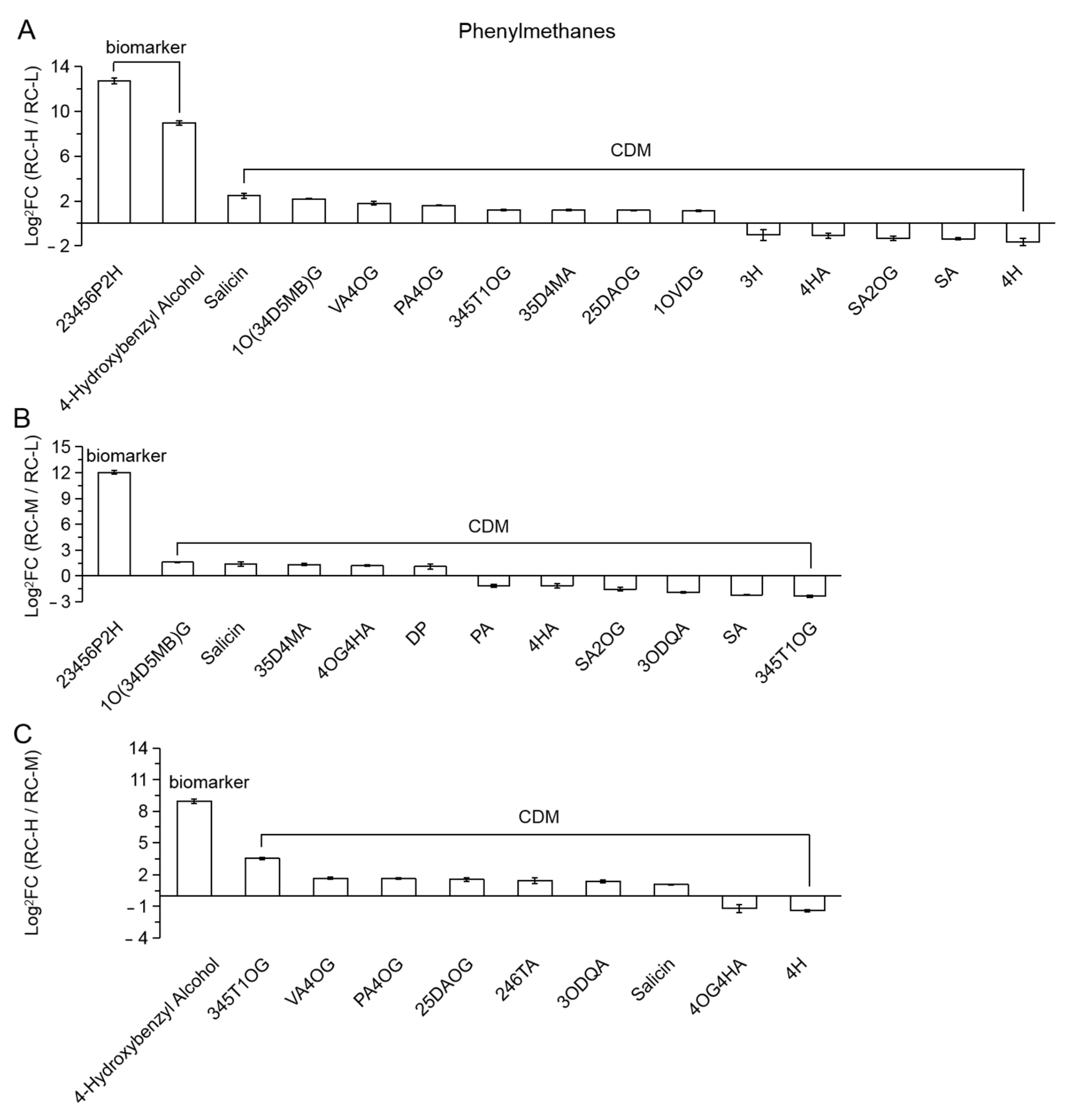
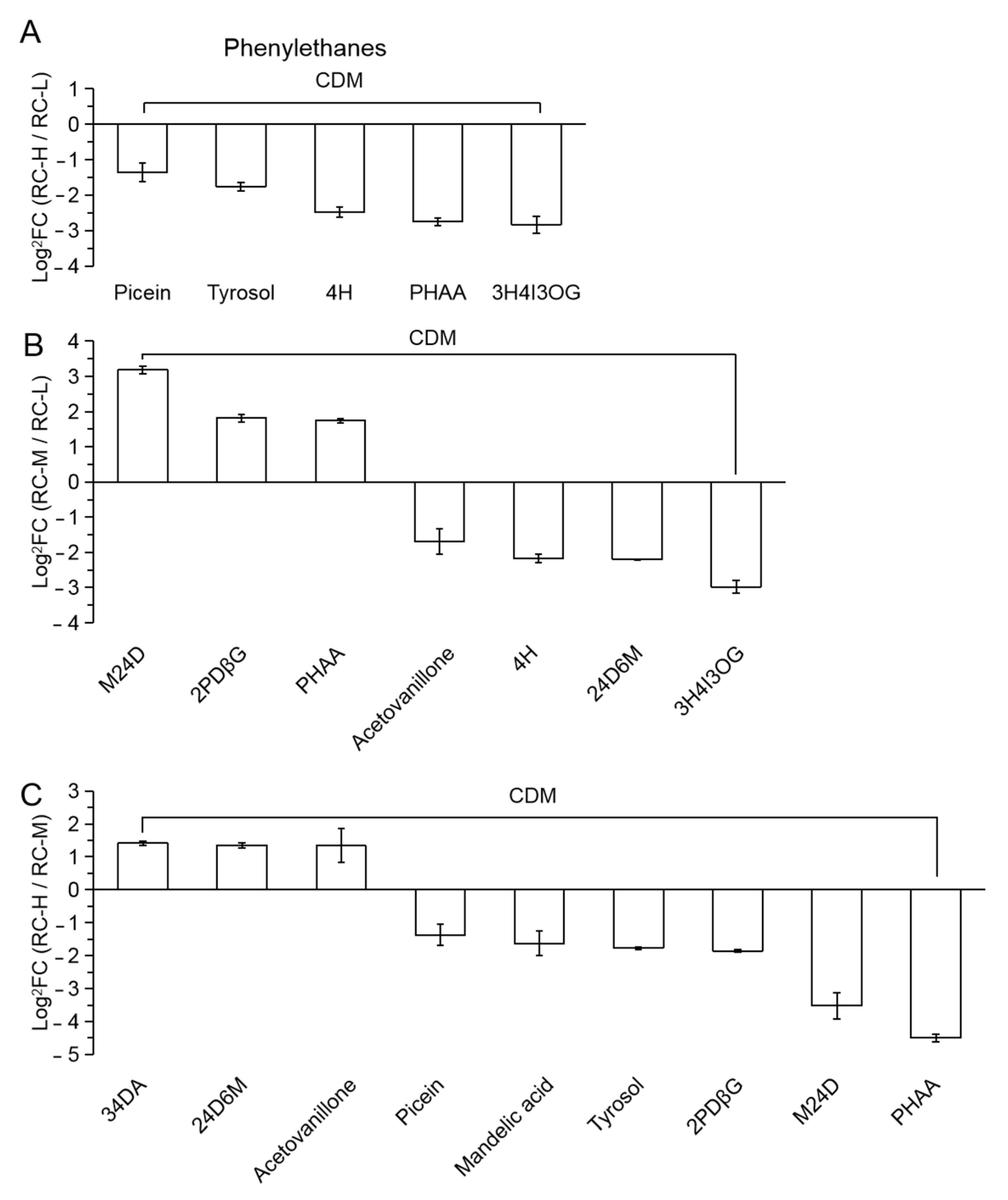
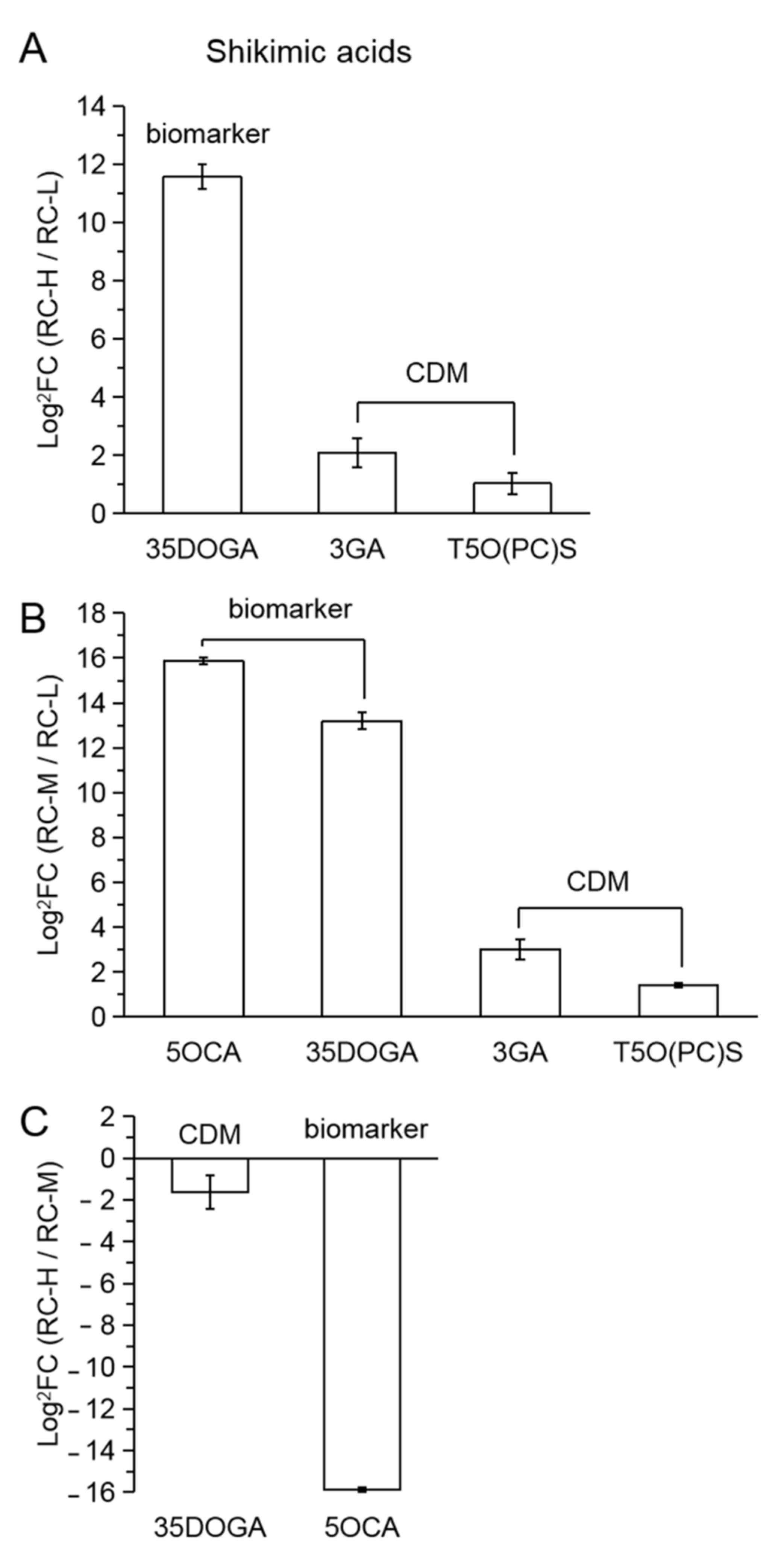
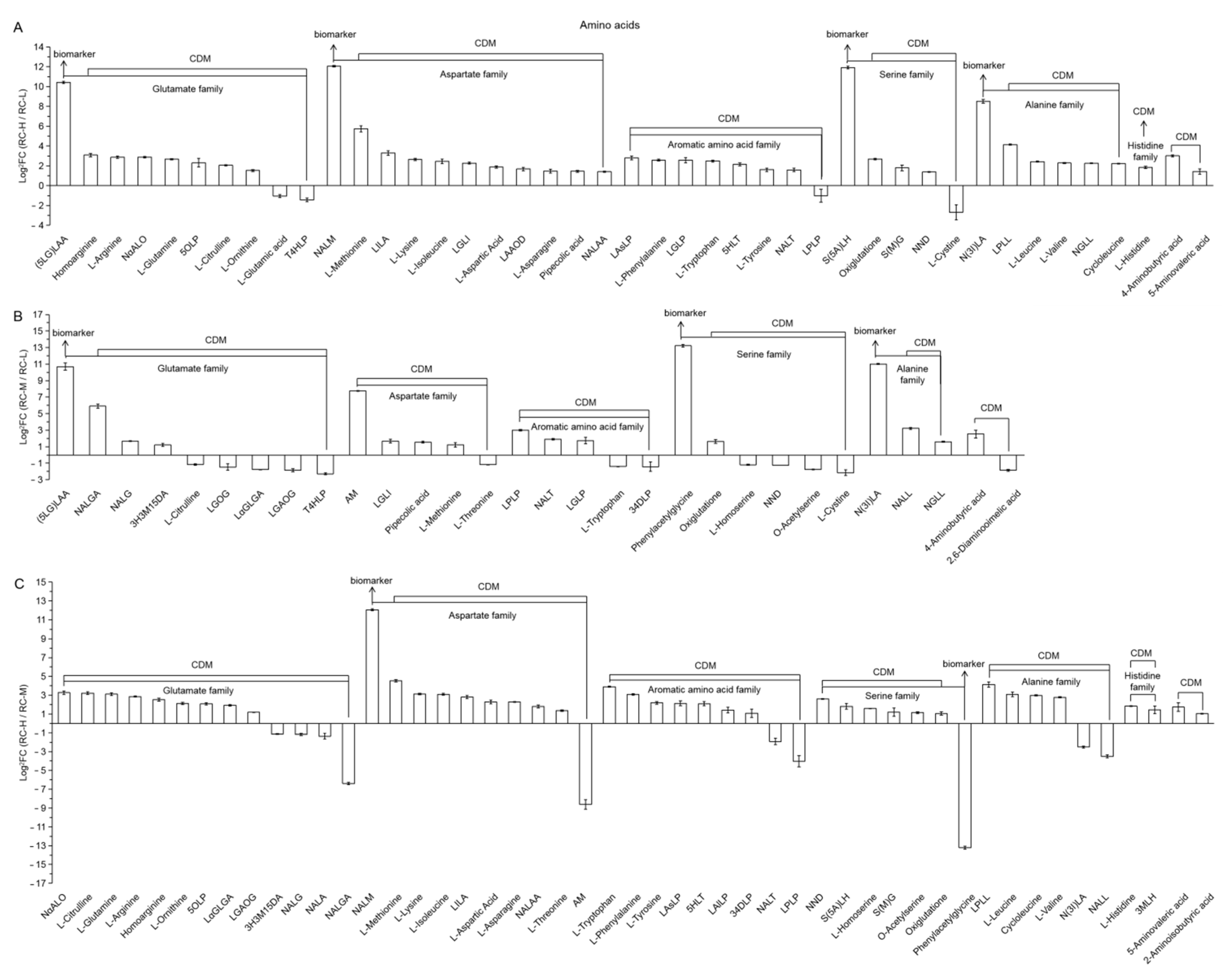
2.4. Characteristics of Chemical Constituents of Rhodiola crenulata in Response to Altitude Gradient
2.4.1. Biomarkers
2.4.2. Six Categories of Differential Metabolites
- Flavonoids
- 2.
- Gallic Acid and derivatives
- 3.
- Phenylpropanoids
- 4.
- Amino Acids
- 5.
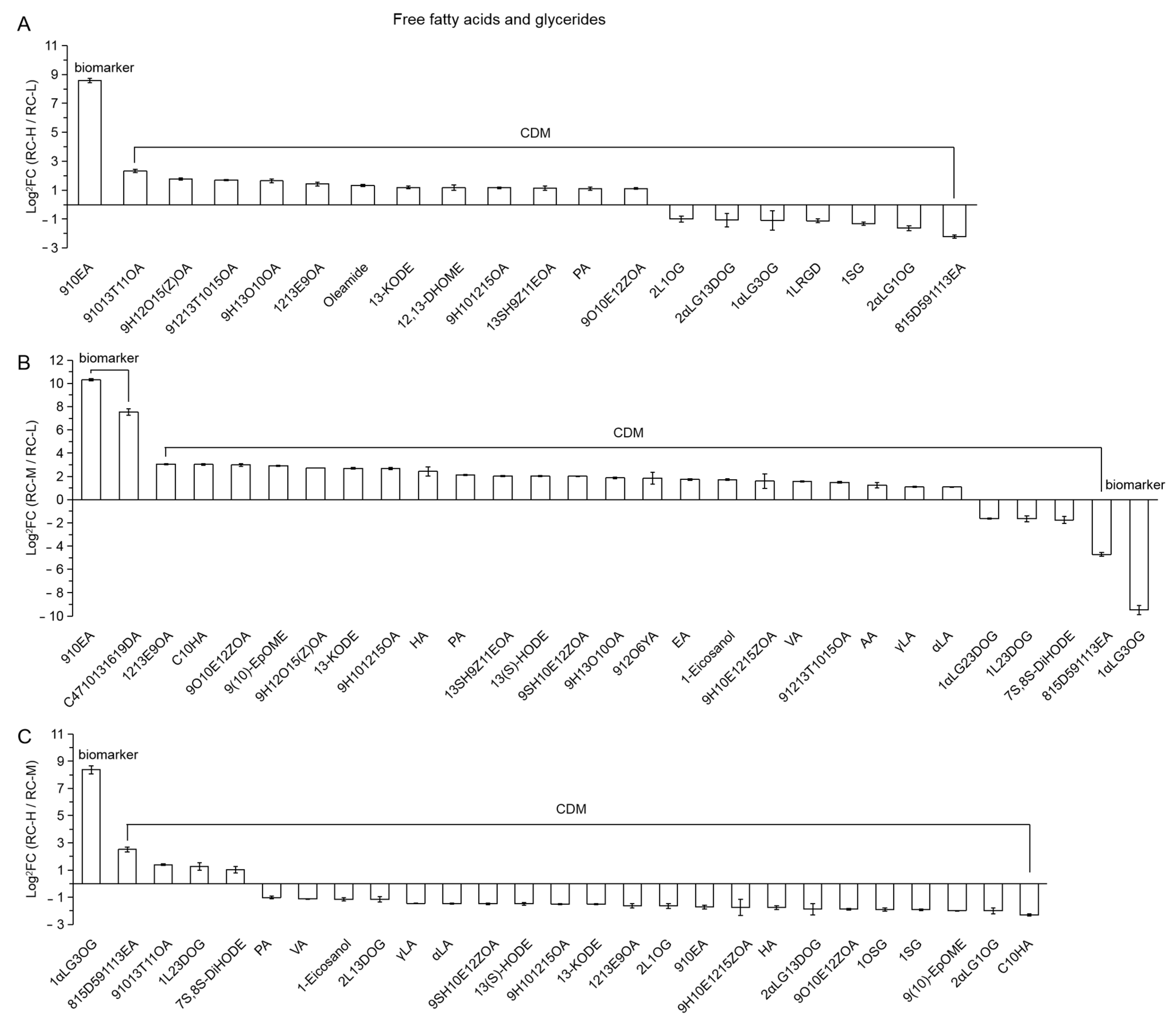
- Free fatty acids and glycerides
- 6.
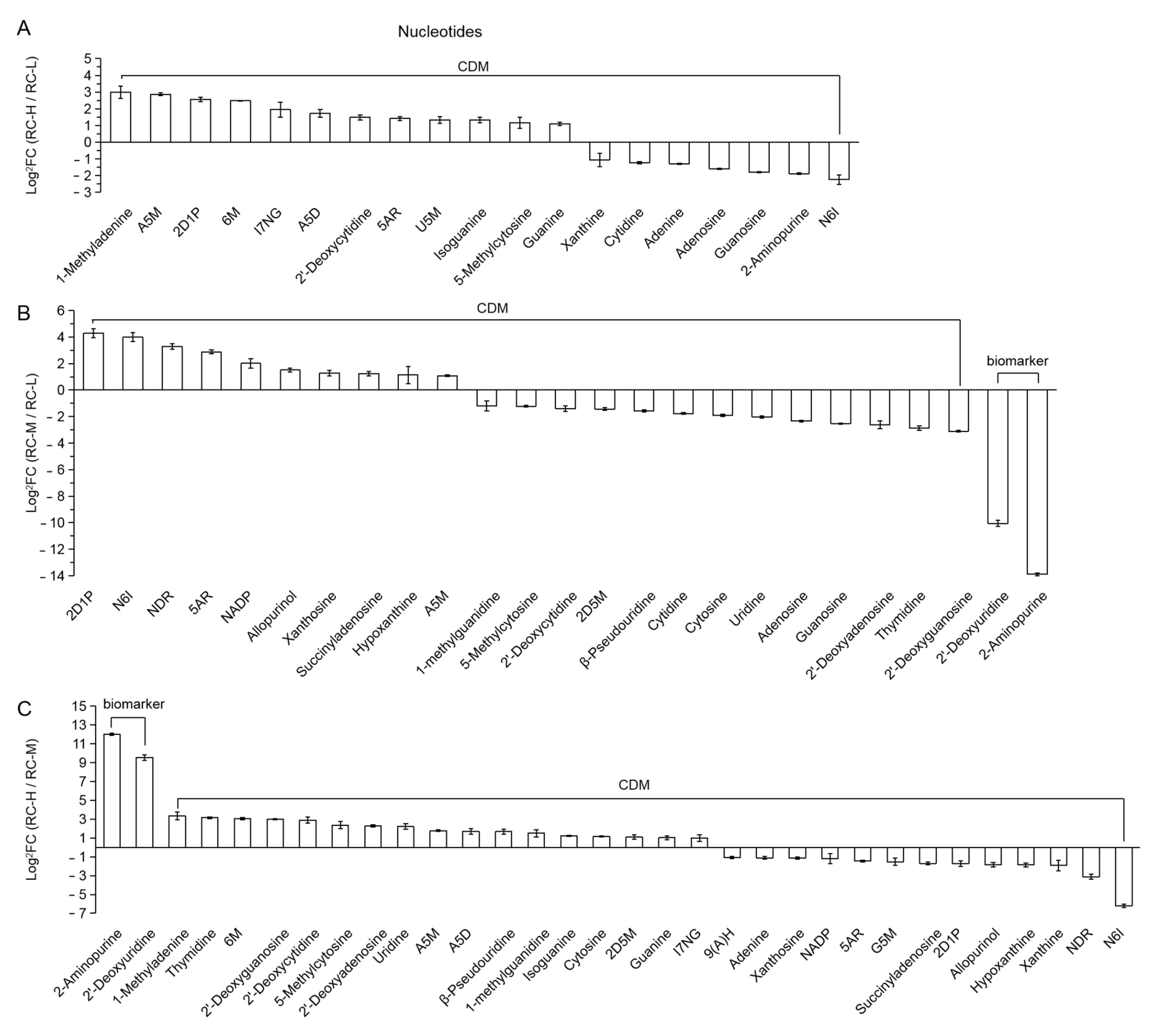
- Nucleotides
3. Materials and Methods
3.1. Plant Materials and Extracts Preparation
3.2. Determination of Oxidative States and Ascorbic Acids
3.3. Determination of Phenolic Components
3.3.1. Total Phenols
3.3.2. Total Tannins
3.3.3. Total Flavonoids
3.3.4. Condensed Tannins
3.4. Determination of Antioxidant Capacity
3.4.1. DPPH-Scavenging Activity
3.4.2. ABTS+-Scavenging Activity
3.5. Mineral Content Analysis
3.6. LC-ESI-MS/MS
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| 1 (Figure 7) | Q3ORu | quercetin-3-O-rutinoside (Rutin) |
| 2 | 7OMQ | 7-O-methxyl quercetin (Rhamnetin) |
| 3 | Q7OR | quercetin-7-O-rutinoside |
| 4 | Q3βDS | quercetin 3-β-D-sophoroside |
| 5 | Q3O(2OG)G | quercetin-3-O-(2′’-O-galactosyl)glucoside |
| 6 | Q3OGl | quercetin-3-O-glucoside (Isoquercitrin) |
| 7 | Q3OS | quercetin-3-O-sophoroside (Baimaside) |
| 8 | Q3OG(1→4)R7OR | quercetin-3-O-glucosyl(1 → 4)rhamnoside-7-O-rutinoside |
| 9 | Q3O(2OR)R | quercetin-3-O-(2′’-O-Rhamnosyl)rutinoside |
| 10 | Q3O(6OA)G | quercetin-3-O-(6′’-O-arabinosyl)glucoside |
| 11 | Q3O(2OG)glucuronide | quercetin-3-O-(2′’-O-glucosyl)glucuronide |
| 12 | 5OM | 5-O-methylquercetin (Azaleatin) |
| 13 | I3OR | isorhamnetin-3-O-rutinoside |
| 14 | R3OR | rhamnetin-3-O-rutinoside |
| 15 | Q7OβDG | quercetin 7-O-β-D-glucoside |
| 16 | Q3OGa | quercetin-3-O-galactoside (Hyperin) |
| 17 | Q3OR7OG | quercetin-3-O-rutinoside-7-O-glucoside |
| 18 | Q3OX | quercetin-3-O-xyloside (Reynoutrin) |
| 19 | Q3ORo | quercetin-3-O-robinobioside |
| 20 | Q3O(6G)G | quercetin-3-O-(6′’-galloyl)galactoside |
| 21 (Figure 8) | D7OG | dihydrokaempferol-7-O-glucoside |
| 22 | 6CM3G | 6-C-methylkaempferol-3-glucoside |
| 23 | D3OG | dihydrokaempferol-3-O-glucoside |
| 24 | 6H36OD | 6-hydroxykaempferol-3,6-O-diglucoside |
| 25 | K68DCG7OG | kaempferol-6,8-di-C-glucoside-7-O-glucoside |
| 26 | K4OG | kaempferol-4′-O-glucoside |
| 27 | K3OGa | kaempferol-3-O-galactoside (Trifolin) |
| 28 | K3ON | kaempferol-3-O-neohesperidoside |
| 29 (Figure 9) | (−)−E3(3OM)G | (−)-epicatechin-3-(3′’-O-methyl)gallate |
| 30 | PC13OG | procyanidin C1 3′-O-gallate |
| 31 (Figure 10) | L3OG | luteolin-3′-O-glucoside |
| 32 | L7O(2OR)R | luteolin-7-O-(2′’-O-rhamnosyl)rutinoside |
| 33 (Figure 11) | 13TA | 1,3-trigallic acid |
| 34 | 3H5M1O(6D)G | 3-hydroxy-5-methylphenol-1-O-(6′-digalloyl) glucoside |
| 35 | 3OMA | 3-O-methylgallic acid |
| 36 | 17DOGDS | 1,7-di-O-galloyl-D-sedoheptulose |
| 37 | M6OGG | methyl 6-O-galloyl-glucoside |
| 38 | 2OS6OGDG | 2-O-salicyl-6-O-galloyl-D-glucose |
| 39 | MD | monogalloyl-diglucose |
| 40 | 14TA | 1,4-trigallic acid |
| 41 | 23OD146TOGG | 2,3-O-digalloyl-1,4,6-tri-O-galloyl-glucose |
| 42 | 1236TOGDG | 1,2,3,6-tetra-O-galloyl-D-glucose |
| 43 | 2OGG | 2-O-galloyl-glucose |
| 44 | 1OGDG | 1-O-galloyl-D-glucose |
| 45 | 6OGG | 6-O-galloyl-glucose |
| 46 | 16DOGDG | 1,6-di-O-galloyl-D-glucose |
| 47 | 3OD1246OTDG | 3-O-digalloyl-1,2,4,6-O-tetragalloyl-D-glucose |
| 48 (Figure 12) | CA | caffeoylbenzoyltartaric acid |
| 49 | 12OD | 1,2-O-diferuloylglycerol |
| 50 | CAME | chlorogenic acid methyl ester |
| 51 | SM | sinapoyl malate |
| 52 | MC | methyl caffeate |
| 53 | 4HA4DG | 4-hydroxycinnamyl alcohol 4-D-glucoside |
| 54 | 7M5P | 7-methoxy-5-prenyloxycoumarin |
| 55 | SADG | syringoylcaffeoylquinic acid-D-glucose |
| 56 | 3OPCAOG | 3-O-p-Coumaroylquinic acid O-glucoside |
| 57 | SA | sinapic acid |
| 58 | 1OGS | 1-O-glucosyl sinapate |
| 59 | 1O[(E)PC]DG | 1-O-[(E)-p-coumaroyl]-D-glucose |
| 60 | 3M48D34D | 3-methyl-4,8-dihydroxy-3,4-dihydroisocoumarin |
| 61 | 6OFDG | 6-O-feruloyl-D-glucose |
| 62 | PCA | p-coumaryl alcohol |
| 63 | 7H5M | 7-hydroxy-5-methoxycoumarin |
| 64 | 2HA | 2-hydroxycinnamic acid |
| 65 | 3(4H)PA | 3-(4-hydroxyphenyl)-propionic acid |
| 66 | C(PH)TA | caffeoyl(p-hydroxybenzoyl)tartaric acid |
| 67 | 3(3H)PA | 3-(3-hydroxyphenyl)-propionate acid |
| 68 | (S)2H3(4H)PA | (S)-2-hydroxy-3-(4-hydroxyphenyl)propanoic acid |
| 69 | PA | phaseolic acid |
| 70 | FA | ferulic acid |
| 71 | BCA | brevifolin carboxylic acid |
| 72 | MCA | maleoyl-caffeoylquinic acid |
| 73 | 57D | 5,7-dimethoxycoumarin |
| 74 | 3OPCA | 3-O-p-coumaroylquinic acid |
| 75 | NA | neochlorogenic acid(5-O-caffeoylquinic acid) |
| 76 | 1O(34D)OCG | 1′-O-(3,4-dihydroxyphenethyl)-O-caffeoyl-glucoside |
| 77 | M4H | methyl 4-hydroxycinnamate |
| 78 (Figure 13) | 23456P2H | 2,3,4,5,6-pentahydroxyhexyl 2-hydroxybenzoate |
| 79 | 1O(34D5MB)G | 1-O-(3,4-dihydroxy-5-methoxy-benzoyl)-glucoside |
| 80 | VA4OG | vanillic acid-4-O-glucoside |
| 81 | PA4OG | protocatechuic acid-4-O-glucoside |
| 82 | 345T1OG | 3,4,5-trimethoxyphenyl-1-O-glucoside |
| 83 | 35D4MA | 3,5-dihydroxy-4-methoxybenzoic acid |
| 84 | 25DAOG | 2,5-dihydroxybenzoic acid O-glucoside |
| 85 | 1OVDG | 1-O-vanilloyl-D-glucose |
| 86 | 3H | 3-hydroxybenzaldehyde |
| 87 | 4HA | 4-hydroxybenzoic acid |
| 88 | SA2OG | salicylic acid-2-O-glucoside |
| 89 | SA | syringic acid |
| 90 | 4H | 4-hydroxybenzaldehyde |
| 91 | 4OG4HA | 4-O-glucosyl-4-hydroxybenzoic acid |
| 92 | DP | dimethyl phthalate |
| 93 | PA | protocatechuic aldehyde |
| 94 | 3ODQA | 3-O-digalloyl quinic acid |
| 95 | 246TA | 2,4,6-trihydroxybenzoic acid |
| 96 (Figure 14) | 4H | 4-hydroxyacetophenone |
| 97 | PHAA | p-hydroxyphenyl acetic acid |
| 98 | 3H4I3OG | 3-hydroxy-4-isopropylbenzylalcohol-3-O-glucoside |
| 99 | M24D | methyl 2,4-dihydroxyphenylacetate |
| 100 | 2PDβG | 2-phenylethyl-D-β-glucopyranoside |
| 101 | 24D6M | 2′,4′-dihydroxy-6′-methoxyacetophenone |
| 102 | 34DA | 3,4-dihydroxybenzeneacetic acid |
| 103 (Figure 15) | 35DOGA | 3,5-di-O-galloylshikimic acid |
| 104 | 3GA | 3-galloylshikimic acid |
| 105 | T5O(PC)S | trans-5-O-(p-coumaroyl) shikimate |
| 106 | 5OCA | 5-O-caffeoylshikimic acid |
| 107 (Figure 16) | (5LG)LAA | (5-L-glutamyl)-L-amino acid |
| 108 | NαALO | N-α-acetyl-L-ornithine |
| 109 | 5OLP | 5-oxo-L-proline |
| 110 | T4HLP | trans-4-hydroxy-L-proline |
| 111 | NALM | N-acetyl-L-methionine |
| 112 | LILA | L-isoleucyl-L-aspartate |
| 113 | LGLI | L-glycyl-L-isoleucine |
| 114 | LAAOD | L-aspartic acid-O-diglucoside |
| 115 | NALAA | N-acetyl-L-aspartic acid |
| 116 | LAsLP | L-aspartyl-L-phenylalanine |
| 117 | LGLP | L-glycyl-L-phenylalanine |
| 118 | 5HLT | 5-hydroxy-L-tryptophan |
| 119 | NALT | N-acetyl-L-tryptophan |
| 120 | LPLP | L-prolyl-L-phenylalanine |
| 121 | S(5A)LH | S-(5′-adenosy)-L-homocysteine |
| 122 | S(M)G | S-(methyl)glutathione |
| 123 | NND | N,N-dimethylglycine |
| 124 | N(3I)LA | N-(3-indolylacetyl)-L-alanine |
| 125 | LPLL | L-prolyl-L-leucine |
| 126 | NGLL | N-glycyl-L-leucine |
| 127 | NALGA | N-acetyl-L-glutamic acid |
| 128 | NALG | N-acetyl-L-glutamine |
| 129 | 3H3M15DA | 3-hydroxy-3-methylpentane-1,5-dioic acid |
| 130 | LGOG | L-glutamine-O-glycoside |
| 131 | LαGLGA | L-α-glutamyl-L-glutamic acid |
| 132 | LGAOG | L-glutamic acid-O-glycoside |
| 133 | AM | acetylleucine monoethanolamine |
| 134 | 34DLP | 3,4-dihydroxy-L-phenylalanine |
| 135 | NALL | N-acetyl-L-leucine |
| 136 | NALA | N-acetyl-L-arginine |
| 137 | LAlLP | L-alanyl-L-phenylalanine |
| 138 | 3MLH | 3-methyl-L-histidine |
| 139 (Figure 17) | 910EA | 9,10-epoxyoctadecanoic acid |
| 140 | 91013T11OA | 9,10,13-trihydroxy-11-octadecenoic acid |
| 141 | 9H12O15(Z)OA | 9-hydroxy-12-oxo-15(Z)-octadecenoic acid |
| 142 | 91213T1015OA | 9,12,13-trihydroxy-10,15-octadecadienoic acid |
| 143 | 9H13O10OA | 9-hydroxy-13-oxo-10-octadecenoic acid |
| 144 | 1213E9OA | 12,13-epoxy-9-octadecenoic acid |
| 145 | 13-KODE | (9Z,11E)-13-oxooctadeca-9,11-dienoic acid |
| 146 | 12,13-DHOME | (9Z)-12,13-dihydroxyoctadec-9-enoic acid |
| 147 | 9H101215OA | 9-hydroxy-10,12,15-octadecatrienoic acid |
| 148 | 13SH9Z11EOA | 13S-hydroperoxy-9Z,11E-octadecadienoic acid |
| 149 | PA | punicic acid |
| 150 | 9O10E12ZOA | 9-oxo-10E,12Z-octadecadienoic acid |
| 151 | 2L1OG | 2-linoleoylglycerol-1-O-glucoside |
| 152 | 2αLG13DOG | 2-α-linolenoyl-glycerol-1,3-di-O-glucoside |
| 153 | 1αLG3OG | 1-α-linolenoyl-glycerol-3-O-glucoside |
| 154 | 1LRGD | 1-linolenoyl-rac-glycerol-diglucoside |
| 155 | 1SG | 1-stearidonoyl-glycerol |
| 156 | 2αLG1OG | 2-α-linolenoyl-glycerol-1-O-glucoside |
| 157 | 815D591113EA | 8,15-dihydroxy-5,9,11,13-eicosatetraenoic acid |
| 158 | C4710131619DA | cis-4,7,10,13,16,19-docosahexaenoic acid |
| 159 | C10HA | cis-10-heptadecenoic acid |
| 160 | 9(10)-EpOME | (9R,10S)-(12Z)-9,10-epoxyoctadecenoic acid |
| 161 | HA | heptadecanoic acid |
| 162 | 13(S)-HODE | 13(S)-hydroxyoctadeca-9Z,11E-dienoic acid |
| 163 | 9SH10E12ZOA | 9S-hydroxy-10E,12Z-octadecadienoic acid |
| 164 | 912O6YA | 9,12-octadecadien-6-ynoic acid |
| 165 | EA | eicosadienoic acid |
| 166 | 9H10E1215ZOA | 9-hydroperoxy-10E,12,15Z-octadecatrienoic acid |
| 167 | VA | vaccenic acid |
| 168 | AA | arachidonic acid |
| 169 | γLA | γ-linolenic acid |
| 170 | αLA | α-linolenic acid |
| 171 | 1αLG23DOG | 1-α-linolenoyl-glycerol-2,3-di-O-glucoside |
| 172 | 1L23DOG | 1-linoleoylglycerol-2,3-di-O-glucoside |
| 173 | 7S,8S-DiHODE | (9Z,12Z)-(7S,8S)-dihydroxyoctadeca-9,12-dienoic acid |
| 174 | 2L13DOG | 2-linoleoylglycerol-1,3-di-O-glucoside |
| 175 | 1OSG | 1-oleoyl-sn-glycerol |
| 176 (Figure 18) | A5M | adenosine 5′-monophosphate |
| 177 | 2D1P | 2-deoxyribose-1-phosphate |
| 178 | 6M | 6-methylmercaptopurine |
| 179 | I7NG | isopentenyladenine-7-N-glucoside |
| 180 | A5D | adenosine 5′-diphosphate |
| 181 | 5AR | 5-aminoimidazole ribonucleotide |
| 182 | U5M | uridine 5′-monophosphate |
| 183 | N6I | N6-isopentenyladenine |
| 184 | NDR | nicotinate D-ribonucleoside |
| 185 | NADP | nicotinamide adenine dinucleotide phosphate |
| 186 | 2D5M | 2′-deoxyadenosine-5′-monophosphate |
| 187 | 9(A)H | 9-(arabinosyl)hypoxanthine |
| 188 | G5M | guanosine 5′-monophosphate |
References
- Panossian, A.; Wikman, G.; Sarris, J. Rosenroot (Rhodiola rosea): Traditional use, chemical composition, pharmacology and clinical efficacy. Phytomedicine 2010, 17, 481–493. [Google Scholar] [CrossRef]
- Panossian, A.G.; Efferth, T.; Shikov, A.N.; Pozharitskaya, O.N.; Kuchta, K.; Mukherjee, P.K.; Banerjee, S.; Heinrich, M.; Wu, W.; Guo, D.; et al. Evolution of the adaptogenic concept from traditional use to medical systems: Pharmacology of stress- and aging-related diseases. Med. Res. Rev. 2021, 41, 630–703. [Google Scholar] [CrossRef]
- Shikov, A.N.; Narkevich, I.A.; Flisyuk, E.V.; Luzhanin, V.G.; Pozharitskaya, O.N. Medicinal plants from the 14th edition of the Russian Pharmacopoeia, recent updates. J. Ethnopharmacol. 2021, 268, 113685. [Google Scholar] [CrossRef]
- Tao, H.; Wu, X.; Cao, J.; Peng, Y.; Wang, A.; Pei, J.; Xiao, J.; Wang, S.; Wang, Y. Rhodiola species: A comprehensive review of traditional use, phytochemistry, pharmacology, toxicity, and clinical study. Med. Res. Rev. 2019, 39, 1779–1850. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of People’s Republic of China; People’s Medical Publishing House: Beijing, China, 2005; pp. 211–212. ISBN 7-117-06982-1/R 6983. [Google Scholar]
- Booker, A.; Zhai, L.; Gkouva, C.; Li, S.; Heinrich, M. From traditional resource to global commodities—A comparison of Rhodiola species using NMR spectroscopy—Metabolomics and HPTLC. Front. Pharmacol. 2016, 7, 254. [Google Scholar] [CrossRef] [Green Version]
- Recio, M.-C.; Giner, R.; Máñez, S. Immunmodulatory and antiproliferative properties of Rhodiola species. Planta Med. 2016, 82, 952–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.-L.; Guo, T.-T.; Ren, Z.-X.; Zhang, N.-S.; Wang, M.-L. Diversity and antioxidant activity of culturable Endophytic fungi from Alpine plants of Rhodiola crenulata, R. angusta, and R. sachalinensis. PLoS ONE 2015, 10, e0118204. [Google Scholar] [CrossRef] [Green Version]
- Bassa, L.M.; Jacobs, C.; Gregory, K.; Henchey, E.; Ser-Dolansky, J.; Schneider, S.S. Rhodiola crenulata induces an early estrogenic response and reduces proliferation and tumorsphere formation over time in MCF7 breast cancer cells. Phytomedicine 2016, 23, 87–94. [Google Scholar] [CrossRef]
- Hsiao, Y.-W.; Tsai, Y.-N.; Huang, Y.-T.; Liu, S.-H.; Lin, Y.-J.; Lo, L.-W.; Hu, Y.-F.; Chung, F.-P.; Lin, S.-F.; Chang, S.-L.; et al. Rhodiola crenulata reduces ventricular arrhythmia through mitigating the activation of IL-17 and inhibiting the MAPK signaling pathway. Cardiovasc. Drugs Ther. 2020, 35, 1–12. [Google Scholar] [CrossRef]
- Chiu, T.-F.; Chen, L.L.-C.; Su, D.-H.; Lo, H.-Y.; Chen, C.-H.; Wang, S.-H.; Chen, W.-L. Rhodiola crenulata extract for prevention of acute mountain sickness: A randomized, double-blind, placebo-controlled, crossover trial. BMC Complement. Altern. Med. 2013, 13, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, Y.; Yang, W.; He, X.; Xu, S.; Liu, X.; He, Y.; Hu, Q.; Yuan, D.; Jin, T. Network pharmacology and molecular docking analysis on mechanisms of Tibetan Hongjingtian (Rhodiola crenulata) in the treatment of COVID-19. J. Med. Microbiol. 2021, 70, 001374. [Google Scholar] [CrossRef] [PubMed]
- Chuang, M.-L.; Wu, T.-C.; Wang, Y.-T.; Wang, Y.-C.; Tsao, T.C.-Y.; Wei, J.C.-C.; Chen, C.-Y.; Lin, I.-F. Adjunctive treatment with Rhodiola Crenulata in patients with chronic obstructive pulmonary disease—A randomized placebo controlled double blind clinical trial. PLoS ONE 2015, 10, e0128142. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Guo, Y.; Xiong, C.; Sun, L. Evaluation of two major Rhodiola species and the systemic changing characteristics of metabolites of Rhodiola crenulata in different altitudes by chemical methods combined with UPLC-QqQ-MS-based metabolomics. Molecules 2020, 25, 4062. [Google Scholar] [CrossRef] [PubMed]
- Rhodiola Crenulata. Available online: http://www.iplant.cn/info/Rhodiola%20crenulata?t=foc (accessed on 28 October 2021).
- Oscar, V.; Monica, B. Flavonoids: Antioxidant compounds for plant defence. and for a healthy human diet. Not. Bot. Horti Agrobot. Cluj Napoca 2017, 46, 14–21. [Google Scholar]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus × acerifolia plants during Mediterranean summers. New Phytol. 2015, 207, 613–626. [Google Scholar] [CrossRef]
- Hagerman, A.E.; Riedl, K.M.; Jones, G.A.; Sovik, K.N.; Ritchard, N.T.; Hartzfeld, A.P.W.; Riechel, T.L. High molecular weight plant Polyphenolics (Tannins) as biological antioxidants. J. Agric. Food Chem. 1998, 46, 1887–1892. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Qin, H.; Li, Z.; Liu, H.; Wang, J.; Zhang, H.; Quan, R.; Huang, R.; Zhang, Z. The synthesis of ascorbic acid in rice roots plays an important role in the salt tolerance of rice by scavenging ROS. Int. J. Mol. Sci. 2018, 19, 3347. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.; Viola, R. Biosynthesis and catabolism of L-Ascorbic acid in plants. Crit. Rev. Plant Sci. 2005, 24, 167–188. [Google Scholar] [CrossRef]
- Xu, C.; Wei, L.; Huang, S.; Yang, C.; Wang, Y.; Yuan, H.; Xu, Q.; Zhang, W.; Wang, M.; Zeng, X.; et al. Drought resistance in Qingke involves a reprogramming of the Phenylpropanoid Pathway and UDP-Glucosyltransferase regulation of abiotic stress tolerance targeting Flavonoid Biosynthesis. J. Agric. Food Chem. 2021, 69, 3992–4005. [Google Scholar] [CrossRef] [PubMed]
- Geng, D.; Shen, X.; Xie, Y.; Yang, Y.; Bian, R.; Gao, Y.; Li, P.; Sun, L.; Feng, H.; Ma, F.; et al. Regulation of phenylpropanoid biosynthesis by MdMYB88 and MdMYB124 contributes to pathogen and drought resistance in apple. Hortic. Res. 2020, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Moreno, M.J.; Omranian, N.; Sáez, P.L.; Figueroa, C.M.; Del-Saz, N.F.; Elso, M.; Poblete, L.; Orf, I.; Cuadros-Inostroza, A.; Cavieres, L.A.; et al. Low-temperature tolerance of the Antarctic species Deschampsia antarctica: A complex metabolic response associated with nutrient remobilization. Plant Cell Environ. 2020, 43, 1376–1393. [Google Scholar] [CrossRef]
- Zhong, Z.; Liu, S.; Zhu, W.; Ou, Y.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Tian, J.; Komatsu, S. Phosphoproteomics reveals the biosynthesis of secondary metabolites in Catharanthus roseus under Ultraviolet-B radiation. J. Proteome Res. 2019, 18, 3328–3341. [Google Scholar] [CrossRef]
- Zhu, L.-J.; Deng, X.-G.; Zhang, D.-W.; Lin, H.-H. Enhancement of stress tolerance in cucumber seedlings by proanthocyanidins. Biol. Plant 2017, 61, 323–332. [Google Scholar] [CrossRef]
- Wilson, K.E.; Wilson, M.I.; Greenberg, B.M. Identification of the Flavonoid Glycosides that accumulate in Brassica napus L. cv. Topas specifically in response to ultraviolet B radiation. Photochem. Photobiol. 1998, 67, 547–553. [Google Scholar] [CrossRef]
- Ortega, E.; Nair, V.; Welti-Chanes, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Wounding and UVB light Synergistically induce the Biosynthesis of Phenolic compounds and Ascorbic Acid in Red Prickly Pears (Opuntia ficus-indica cv. Rojo Vigor). Int. J. Mol. Sci. 2019, 20, 5327. [Google Scholar] [CrossRef] [Green Version]
- Xiang, N.; Hu, J.; Wen, T.; Brennan, M.A.; Brennan, C.S.; Guo, X. Effects of temperature stress on the accumulation of ascorbic acid and folates in sweet corn (Zea mays L.) seedlings. J. Sci. Food Agric. 2019, 100, 1694–1701. [Google Scholar] [CrossRef]
- Varela, M.C.; Arslan, I.; Reginato, M.A.; Cenzano, A.M.; Luna, M.V. Phenolic compounds as indicators of drought resistance in shrubs from Patagonian shrublands (Argentina). Plant Physiol. Biochem. 2016, 104, 81–91. [Google Scholar] [CrossRef]
- Amir, R.; Galili, G.; Cohen, H. The metabolic roles of free amino acids during seed development. Plant Sci. 2018, 275, 11–18. [Google Scholar] [CrossRef]
- Moe, L.A. Amino acids in the rhizosphere: From plants to microbes. Am. J. Bot. 2013, 100, 1692–1705. [Google Scholar] [CrossRef]
- Harding, H.; Zhang, Y.; Zeng, H.; Novoa, I.; Lu, P.D.; Calfon, M.; Sadri, N.; Yun, C.; Popko, B.; Paules, R.S.; et al. An integrated stress response regulates Amino Acid metabolism and resistance to oxidative stress. Mol. Cell 2003, 11, 619–633. [Google Scholar] [CrossRef]
- Nehela, Y.; Killiny, N. ‘Candidatus Liberibacter asiaticus’ and its vector, Diaphorina citri, Augment the Tricarboxylic acid cycle of their host via the γ-Aminobutyric acid shunt and polyamines pathway. Mol. Plant Microbe Interact. 2019, 32, 413–427. [Google Scholar] [CrossRef] [Green Version]
- Cheng, B.; Li, Z.; Liang, L.; Cao, Y.; Zeng, W.; Zhang, X.; Ma, X.; Huang, L.; Nie, G.; Liu, W.; et al. The γ-Aminobutyric acid (GABA) Alleviates salt stress damage during seeds germination of white clover associated with Na+/K+ transportation, Dehydrins accumulation, and stress-related genes expression in white clover. Int. J. Mol. Sci. 2018, 19, 2520. [Google Scholar] [CrossRef]
- Adams, Z.; Ehlting, J.; Edwards, R. The regulatory role of shikimate in plant phenylalanine metabolism. J. Theor. Biol. 2019, 462, 158–170. [Google Scholar] [CrossRef]
- Manela, N.; Oliva, M.; Ovadia, R.; Sikron-Persi, N.; Ayenew, B.; Fait, A.; Galili, G.; Perl, A.; Weiss, D.; Oren-Shamir, M. Phenylalanine and tyrosine levels are rate-limiting factors in production of health promoting metabolites in Vitis vinifera cv. Gamay Red cell suspension. Front. Plant Sci. 2015, 6, 538. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Chapter 13: Secondary metabolites and plant defense. In Plant Physiology, 4th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2006; pp. 290–292. ISBN 9780878938568. [Google Scholar]
- Acuña-Avila, P.E.; Vásquez-Murrieta, M.S.; Hernández, M.O.F.; López-Cortéz, M.D.S. Relationship between the elemental composition of grapeyards and bioactive compounds in the Cabernet Sauvignon grapes Vitis vinífera harvested in Mexico. Food Chem. 2016, 203, 79–85. [Google Scholar] [CrossRef]
- Martins, V.; Garcia, A.; Costa, C.; Sottomayor, M.; Gerós, H. Calcium- and hormone-driven regulation of secondary metabolism and cell wall enzymes in grape berry cells. J. Plant Physiol. 2018, 231, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langer, S.E.; Marina, M.; Burgos, J.L.; Martínez, G.A.; Civello, P.M.; Villarreal, N.M. Calcium chloride treatment modifies cell wall metabolism and activates defense responses in strawberry fruit (Fragaria × ananassa, Duch). J. Sci. Food Agric. 2019, 99, 4003–4010. [Google Scholar] [CrossRef]
- Kováčik, J.; Klejdus, B.; Bačkor, M. Phenolic metabolism of Matricaria chamomilla plants exposed to nickel. J. Plant Physiol. 2009, 166, 1460–1464. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Song, H.; Guan, C.; Zhang, Z. Boron mitigates cadmium toxicity to rapeseed (Brassica napus) shoots by relieving oxidative stress and enhancing cadmium chelation onto cell walls. Environ. Pollut. 2020, 263, 114546. [Google Scholar] [CrossRef]
- Hanaka, A.; Dresler, S.; Wójciak-Kosior, M.; Strzemski, M.; Kováčik, J.; Latalski, M.; Zawiślak, G.; Sowa, I. the impact of long-and short-term Strontium treatment on metabolites and minerals in Glycine max. Molecules 2019, 24, 3825. [Google Scholar] [CrossRef] [Green Version]
- Han, T.; Sun, M.; Zhao, J.; Dai, C.; Li, Y.; Zhang, P.; Lang, D.; Zhou, J.; Li, X.; Ge, S. The roles of cadmium on growth of seedlings by analysing the composition of metabolites in pumpkin tissues. Ecotoxicol. Environ. Saf. 2021, 226, 112817. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhou, X.; Liu, D.; Feng, W. Enhancing the activity and stability of Mn-superoxide dismutase by one-by-one ligation to catalase. Free. Radic. Biol. Med. 2018, 129, 138–145. [Google Scholar] [CrossRef]
- Robinett, N.G.; Peterson, R.L.; Culotta, V.C. Eukaryotic copper-only superoxide dismutases (SODs): A new class of SOD enzymes and SOD-like protein domains. J. Biol. Chem. 2018, 293, 4636–4643. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.R.; Dubey, K.; Goswami, S.; Hasija, S.; Pandey, R.; Singh, P.K.; Singh, B.; Sareen, S.; Rai, G.K.; Singh, G.P.; et al. Heterologous expression and characterization of novel manganese superoxide dismutase (Mn-SOD)—A potential biochemical marker for heat stress-tolerance in wheat (Triticum aestivum). Int. J. Biol. Macromol. 2020, 161, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.E.; Castro, P.R.; Azevedo, R.A. Hormesis in plants under Cd exposure: From toxic to beneficial element? J. Hazard. Mater. 2020, 384, 121434. [Google Scholar] [CrossRef]
- Singh, D.P.; Liu, L.H.; Øiseth, S.K.; Beloy, J.; Lundin, L.; Gidley, M.J.; Day, L. Influence of boron on carrot cell wall structure and its resistance to fracture. J. Agric. Food Chem. 2010, 58, 9181–9189. [Google Scholar] [CrossRef] [PubMed]
- Song, W.-Y.; Zhang, Z.-B.; Shao, H.-B.; Guo, X.-L.; Cao, H.-X.; Zhao, H.-B.; Fu, Z.-Y.; Hu, X.-J. Relationship between calcium decoding elements and plant abiotic-stress resistance. Int. J. Biol. Sci. 2008, 4, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Martens, S.N.; Boyd, R.S. The ecological significance of nickel hyperaccumulation: A plant chemical defense. Oecologia 1994, 98, 379–384. [Google Scholar] [CrossRef]
- Peterson, R.L.; Galaleldeen, A.; Villarreal, J.; Taylor, A.B.; Cabelli, D.E.; Hart, P.J.; Culotta, V.C. The phylogeny and active site design of Eukaryotic Copper-only superoxide dismutases. J. Biol. Chem. 2016, 291, 20911–20923. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Ding, N.-Z. Plant unsaturated fatty acids: Multiple roles in stress response. Front. Plant Sci. 2020, 11, 562785. [Google Scholar] [CrossRef]
- Cao, Y.; Tanaka, K.; Nguyen, C.; Stacey, G. Extracellular ATP is a central signaling molecule in plant stress responses. Curr. Opin. Plant Biol. 2014, 20, 82–87. [Google Scholar] [CrossRef]
- Gao, L.; Chen, W.; Xu, X.; Zhang, J.; Singh, T.; Liu, S.; Zhang, D.; Tian, L.; White, A.; Shrestha, P.; et al. Engineering Trienoic fatty acids into Cottonseed oil improves low-temperature seed germination, plant photosynthesis and cotton fiber quality. Plant Cell Physiol. 2020, 61, 1335–1347. [Google Scholar] [CrossRef]
- Kalinger, R.S.; Pulsifer, I.P.; Hepworth, S.R.; Rowland, O. Fatty acyl synthetases and Thioesterases in plant lipid metabolism: Diverse functions and biotechnological applications. Lipids 2020, 55, 435–455. [Google Scholar] [CrossRef]
- De Col, V.; Fuchs, P.; Nietzel, T.; Elsässer, M.; Voon, C.P.; Candeo, A.; Seeliger, I.; Fricker, M.D.; Grefen, C.; Møller, I.M.; et al. ATP sensing in living plant cells reveals tissue gradients and stress dynamics of energy physiology. eLife 2017, 6. [Google Scholar] [CrossRef]
- Chen, Z.; He, C.; Hu, H. Temperature responses of growth, photosynthesis, fatty acid and nitrate reductase in Antarctic and temperate Stichococcus. Extremophiles 2011, 16, 127–133. [Google Scholar] [CrossRef]
- Baâtour, O.; Kaddour, R.; Mahmoudi, H.; Tarchoun, I.; Bettaieb, I.; Nasri, N.; Mrah, S.; Hamdaoui, G.; Lachaâl, M.; Marzouk, B. Salt effects on Origanum majorana fatty acid and essential oil composition. J. Sci. Food Agric. 2011, 91, 2613–2620. [Google Scholar] [CrossRef]
- Yu, P.; Jiang, N.; Fu, W.; Zheng, G.; Li, G.; Feng, B.; Chen, T.; Ma, J.; Li, H.; Tao, L.; et al. ATP Hydrolysis determines cold tolerance by regulating available energy for Glutathione synthesis in rice seedling plants. Rice 2020, 13, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Bajerski, F.; Stock, J.; Hanf, B.; Darienko, T.; Heine-Dobbernack, E.; Lorenz, M.; Naujox, L.; Keller, E.R.J.; Schumacher, H.M.; Friedl, T.; et al. ATP content and cell viability as indicators for Cryostress across the diversity of life. Front. Physiol. 2018, 9, 921. [Google Scholar] [CrossRef]
- Zhu, W.; Miao, Q.; Sun, D.; Yang, G.; Wu, C.; Huang, J.; Zheng, C. The mitochondrial phosphate transporters modulate plant responses to salt stress via affecting ATP and Gibberellin metabolism in Arabidopsis thaliana. PLoS ONE 2012, 7, e43530. [Google Scholar] [CrossRef] [Green Version]
- Reyes, T.H.; Esparza, E.; Crestani, G.; Limonchi, F.; Cruz, R.; Salinas, N.; Scartazza, A.; Guglielminetti, L.; Cosio, E. Physiological responses of maca (Lepidium meyenii Walp.) plants to UV radiation in its high-altitude mountain ecosystem. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cui, G.; Wei, X.; Degen, A.A.; Wei, X.; Zhou, J.; Ding, L.; Shang, Z.; Liu, S.; Long, R. Trolox-equivalent antioxidant capacity and composition of five alpine plant species growing at different elevations on the Qinghai–Tibetan Plateau. Plant Ecol. Divers. 2016, 9, 387–396. [Google Scholar] [CrossRef]
- Neuner, G. Frost resistance in alpine woody plants. Front. Plant Sci. 2014, 5, 654. [Google Scholar] [CrossRef] [Green Version]
- Mehla, N.; Sindhi, V.; Josula, D.; Bisht, P.; Wani, S.H. Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Khan, M.I.R., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 1–23. ISBN 978-981-10-5253-8. [Google Scholar]
- Alché, J.D.D. A concise appraisal of lipid oxidation and lipoxidation in higher plants. Redox Biol. 2019, 23, 101136. [Google Scholar] [CrossRef]
- Sytykiewicz, H.; Łukasik, I.; Golawska, S.; Chrzanowski, G. Aphid-triggered changes in oxidative damage markers of nucleic acids, proteins, and lipids in Maize (Zea mays L.) seedlings. Int. J. Mol. Sci. 2019, 20, 3742. [Google Scholar] [CrossRef] [Green Version]
- Ciacka, K.; Tymiński, M.; Gniazdowska, A.; Krasuska, U. Carbonylation of proteins—An element of plant ageing. Planta 2020, 252, 1–13. [Google Scholar] [CrossRef]
- Singh, P.; Negi, J.S.; Rawat, M.S.M.; Pant, G.J.N. Quantification of mineral elements of Rheum emodi Wallr. (Polygonaceae). Biol. Trace Elem. Res. 2010, 138, 293–299. [Google Scholar] [CrossRef]
- Negi, J.S.; Singh, P.; Pant, G.J.N.; Rawat, M.S.M. Study on the variations of mineral elements in Swertia speciosa (G. Don). Biol. Trace Elem. Res. 2010, 138, 300–306. [Google Scholar] [CrossRef]
- Zeng, X.; Yuan, H.; Dong, X.; Peng, M.; Jing, X.; Xu, Q.; Tang, T.; Wang, Y.; Zha, S.; Gao, M.; et al. Genome-wide dissection of Co-selected UV-B responsive pathways in the UV-B adaptation of Qingke. Mol. Plant 2020, 13, 112–127. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Ganzera, M.; Guggenberger, M.; Stuppner, H.; Zidorn, C. Altitudinal variation of secondary metabolite profiles in flowering heads of Matricaria chamomilla cv. BONA. Planta Med. 2008, 74, 453–457. [Google Scholar] [CrossRef]
- Kumari, M.; Joshi, R.; Kumar, R. Metabolic signatures provide novel insights to Picrorhiza kurroa adaptation along the altitude in Himalayan region. Metabolomics 2020, 16, 1–17. [Google Scholar] [CrossRef]
- De Oliveira, J.B.; Egipto, R.; Laureano, O.; de Castro, R.; Pereira, G.E.; Ricardo-Da-Silva, J.M. Climate effects on physicochemical composition of Syrah grapes at low and high altitude sites from tropical grown regions of Brazil. Food Res. Int. 2019, 121, 870–879. [Google Scholar] [CrossRef]
- Ni, Q.; Wang, Z.; Xu, G.; Gao, Q.; Yang, D.; Morimatsu, F.; Zhang, Y. Altitudinal variation of antioxidant components and capability in Indocalamus latifolius (Keng) McClure Leaf. J. Nutr. Sci. Vitaminol. 2013, 59, 336–342. [Google Scholar] [CrossRef]
- Barnes, P.W.; Flint, S.D.; Tobler, M.A.; Ryel, R.J. Diurnal adjustment in ultraviolet sunscreen protection is widespread among higher plants. Oecologia 2016, 181, 55–63. [Google Scholar] [CrossRef]
- Rizi, M.R.; Azizi, A.; Sayyari, M.; Mirzaie-Asl, A.; Conti, L. Increased phenylpropanoids production in UV-B irradiated Salvia verticillata as a consequence of altered genes expression in young leaves. Plant Physiol. Biochem. 2021, 167, 174–184. [Google Scholar] [CrossRef]
- Rozema, J.; Björn, L.; Bornman, J.; Gaberščik, A.; Häder, D.-P.; Trošt, T.; Germ, M.; Klisch, M.; Gröniger, A.; Sinha, R.; et al. The role of UV-B radiation in aquatic and terrestrial ecosystems—an experimental and functional analysis of the evolution of UV-absorbing compounds. J. Photochem. Photobiol. B Biol. 2002, 66, 2–12. [Google Scholar] [CrossRef]
- Solovchenko, A. Significance of skin flavonoids for UV-B-protection in apple fruits. J. Exp. Bot. 2003, 54, 1977–1984. [Google Scholar] [CrossRef]
- Shafi, M.; Mansoor, S.; Jan, S.; Singh, D.B.; Kazi, M.; Raish, M.; Al Wadei, M.; Mir, J.I.; Ahmad, P. Variability in Catechin and Rutin contents and their antioxidant potential in diverse apple Genotypes. Molecules 2019, 24, 943. [Google Scholar] [CrossRef] [Green Version]
- Rajan, M.; Feba, K.S.; Chandran, V.; Shahena, S.; Mathew, L. Enhancement of rhamnetin production in Vernonia anthelmintica (L.) Willd. cell suspension cultures by eliciting with methyl jasmonate and salicylic acid. Physiol. Mol. Biol. Plants 2020, 26, 1531–1539. [Google Scholar] [CrossRef]
- An, J.-P.; Li, R.; Qu, F.-J.; You, C.-X.; Wang, X.-F.; Hao, Y.-J. R2R3-MYB transcription factor MdMYB23 is involved in the cold tolerance and proanthocyanidin accumulation in apple. Plant J. 2018, 96, 562–577. [Google Scholar] [CrossRef] [Green Version]
- Neugart, S.; Bumke-Vogt, C. Flavonoid Glycosides in Brassica species respond to UV-B depending on exposure time and adaptation time. Molecules 2021, 26, 494. [Google Scholar] [CrossRef]
- Spitaler, R.; Winkler, A.; Lins, I.; Yanar, S.; Stuppner, H.; Zidorn, C. Altitudinal variation of phenolic contents in flowering heads of Arnica montana cv. ARBO: A 3-year comparison. J. Chem. Ecol. 2008, 34, 369–375. [Google Scholar] [CrossRef]
- Ohno, A.; Oka, K.; Sakuma, C.; Okuda, H.; Fukuhara, K. Characterization of tea cultivated at four different altitudes using 1H NMR analysis coupled with multivariate statistics. J. Agric. Food Chem. 2011, 59, 5181–5187. [Google Scholar] [CrossRef]
- Berardi, A.E.; Fields, P.D.; Abbate, J.L.; Taylor, D.R. Elevational divergence and clinal variation in floral color and leaf chemistry in Silene vulgaris. Am. J. Bot. 2016, 103, 1508–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rather, R.A.; Bhagat, M. Quercetin as an innovative therapeutic tool for cancer chemoprevention: Molecular mechanisms and implications in human health. Cancer Med. 2020, 9, 9181–9192. [Google Scholar] [CrossRef]
- Chen, A.Y.; Chen, Y.C. A review of the dietary flavonoid, kaempferol on human health and cancer chemoprevention. Food Chem. 2013, 138, 2099–2107. [Google Scholar] [CrossRef] [Green Version]
- Mishra, C.B.; Pandey, P.; Sharma, R.D.; Malik, Z.; Mongre, R.K.; Lynn, A.M.; Prasad, R.; Jeon, R.; Prakash, A. Identifying the natural polyphenol catechin as a multi-targeted agent against SARS-CoV-2 for the plausible therapy of COVID-19: An integrated computational approach. Brief. Bioinform. 2021, 22, 1346–1360. [Google Scholar] [CrossRef]
- Koirala, N.; Pandey, R.P.; Parajuli, P.; Jung, H.J.; Sohng, J.K. Methylation and subsequent glycosylation of 7,8-dihydroxyflavone. J. Biotechnol. 2014, 184, 128–137. [Google Scholar] [CrossRef]
- Sordon, S.; Popłoński, J.; Huszcza, E. Microbial Glycosylation of Flavonoids. Pol. J. Microbiol. 2016, 65, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Stürzenbaum, S.R. Proanthocyanidins of natural origin: Molecular mechanisms and implications for lipid disorder and aging-associated diseases. Adv. Nutr. 2019, 10, 464–478. [Google Scholar] [CrossRef]
- Sandhu, A.K.; Miller, M.G.; Thangthaeng, N.; Scott, T.M.; Shukitt-Hale, B.; Edirisinghe, I.; Burton-Freeman, B. Metabolic fate of strawberry polyphenols after chronic intake in healthy older adults. Food Funct. 2018, 9, 96–106. [Google Scholar] [CrossRef]
- Kim, Y.-J. Antimelanogenic and antioxidant properties of gallic acid. Biol. Pharm. Bull. 2007, 30, 1052–1055. [Google Scholar] [CrossRef] [Green Version]
- Punia, A.; Chauhan, N.S.; Singh, D.; Kesavan, A.K.; Kaur, S.; Sohal, S.K. Effect of gallic acid on the larvae of Spodoptera litura and its parasitoid Bracon hebetor. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Rubin-Aguirre, A.; Saenz-Romero, C.; Lindig-Cisneros, R.; Del-Rio-Mora, A.; Tena-Morelos, C.; Campos-Bolaños, R.; Del-Val, E. Bark beetle pests in an altitudinal gradient of a Mexican managed forest. For. Ecol. Manag. 2015, 343, 73–79. [Google Scholar] [CrossRef]
- Rudrappa, T.; Choi, Y.S.; Levia, D.F.; Legates, D.R.; Lee, K.H.; Bais, H.P. Phragmites australisroot secreted phytotoxin undergoes photo-degradation to execute severe phytotoxicity. Plant Signal. Behav. 2009, 4, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Neelam; Khatkar, A.; Sharma, K.K. Phenylpropanoids and its derivatives: Biological activities and its role in food, pharmaceutical and cosmetic industries. Crit. Rev. Food Sci. Nutr. 2020, 60, 2655–2675. [Google Scholar] [CrossRef]
- Zhang, S.; Huang, Y.; Li, Y.; Wang, Y.; He, X. Anti-neuroinflammatory and antioxidant phenylpropanoids from Chinese olive. Food Chem. 2019, 286, 421–427. [Google Scholar] [CrossRef]
- Kolaj, I.; Liyanage, S.I.; Weaver, D.F. Phenylpropanoids and Alzheimer’s disease: A potential therapeutic platform. Neurochem. Int. 2018, 120, 99–111. [Google Scholar] [CrossRef]
- Kishorekumar, R.; Bulle, M.; Wany, A.; Gupta, K.J. An overview of important enzymes involved in nitrogen assimilation of plants. Springer Protoc. Handb. 2020, 2057, 1–13. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Chapter 12: Assimilation of mineral nutrients. In Plant Physiology, 4th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2006; pp. 265–266. ISBN 9780878938568. [Google Scholar]
- Rehman, H.U.; Alharby, H.F.; Bamagoos, A.A.; Abdelhamid, M.T.; Rady, M.M. Sequenced application of glutathione as an antioxidant with an organic biostimulant improves physiological and metabolic adaptation to salinity in wheat. Plant Physiol. Biochem. 2021, 158, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Schenck, C.; Maeda, H.A. Tyrosine biosynthesis, metabolism, and catabolism in plants. Phytochemistry 2018, 149, 82–102. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological implications of arginine metabolism in plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef] [Green Version]
- Preczenhak, A.P.; Orsi, B.; Lima, G.P.P.; Tezotto-Uliana, J.V.; Minatel, I.O.; Kluge, R. Cysteine enhances the content of betalains and polyphenols in fresh-cut red beet. Food Chem. 2019, 286, 600–607. [Google Scholar] [CrossRef]
- Wang, Y.; Xiong, F.; Nong, S.; Liao, J.; Xing, A.; Shen, Q.; Ma, Y.; Fang, W.; Zhu, X. Effects of nitric oxide on the GABA, polyamines, and proline in tea (Camellia sinensis) roots under cold stress. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Anwar, A.; She, M.; Wang, K.; Riaz, B.; Ye, X. Biological roles of Ornithine Aminotransferase (OAT) in plant stress tolerance: Present progress and future perspectives. Int. J. Mol. Sci. 2018, 19, 3681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiaochuang, C.; Chu, Z.; Lianfeng, Z.; Junhua, Z.; Hussain, S.; Lianghuan, W.; Qianyu, J. Glycine increases cold tolerance in rice via the regulation of N uptake, physiological characteristics, and photosynthesis. Plant Physiol. Biochem. 2017, 112, 251–260. [Google Scholar] [CrossRef]
- Han, M.; Zhang, C.; Suglo, P.; Sun, S.; Wang, M.; Su, T. l-Aspartate: An essential metabolite for plant growth and stress acclimation. Molecules 2021, 26, 1887. [Google Scholar] [CrossRef]
- Arruda, P.; Barreto, P. Lysine Catabolism through the Saccharopine pathway: Enzymes and intermediates involved in plant responses to abiotic and biotic stress. Front. Plant Sci. 2020, 11, 587. [Google Scholar] [CrossRef]
- Joshi, V.; Jander, G. Arabidopsis Methionine γ-Lyase is regulated according to Isoleucine Biosynthesis needs but plays a subordinate role to Threonine Deaminase. Plant Physiol. 2009, 151, 367–378. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zayed, O.; Yu, Z.; Jiang, W.; Zhu, P.; Hsu, C.-C.; Zhang, L.; Tao, W.A.; Lozano-Durán, R.; Zhu, J.-K. Leucine-rich repeat extensin proteins regulate plant salt tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, 13123–13128. [Google Scholar] [CrossRef] [Green Version]
- Gipson, A.; Morton, K.J.; Rhee, R.J.; Simo, S.; Clayton, J.A.; Perrett, M.E.; Binkley, C.G.; Jensen, E.L.; Oakes, D.L.; Rouhier, M.F.; et al. Disruptions in valine degradation affect seed development and germination in Arabidopsis. Plant J. 2017, 90, 1029–1039. [Google Scholar] [CrossRef]
- Sazuka, T.; Kamiya, N.; Nishimura, T.; Ohmae, K.; Sato, Y.; Imamura, K.; Nagato, Y.; Koshiba, T.; Nagamura, Y.; Ashikari, M.; et al. A rice tryptophan deficient dwarf mutant, tdd1, contains a reduced level of indole acetic acid and develops abnormal flowers and organless embryos. Plant J. 2009, 60, 227–241. [Google Scholar] [CrossRef]
- Thevenet, D.; Pastor, V.; Baccelli, I.; Balmer, A.; Vallat, A.; Neier, R.; Glauser, G.; Mauch-Mani, B. The priming molecule β-aminobutyric acid is naturally present in plants and is induced by stress. New Phytol. 2017, 213, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Balmer, A.; Glauser, G.; Mauch-Mani, B.; Baccelli, I. Accumulation patterns of endogenous β-aminobutyric acid during plant development and defence in Arabidopsis thaliana. Plant Biol. 2018, 21, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Kuczyńska, A.; Cardenia, V.; Ogrodowicz, P.; Kempa, M.; Rodriguez-Estrada, M.T.; Mikołajczak, K. Effects of multiple abiotic stresses on lipids and sterols profile in barley leaves (Hordeum vulgare L.). Plant Physiol. Biochem. 2019, 141, 215–224. [Google Scholar] [CrossRef]
- Xu, C.; Shanklin, J. Triacylglycerol metabolism, function, and accumulation in plant vegetative tissues. Annu. Rev. Plant Biol. 2016, 67, 179–206. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, X.; Cao, X.; Hu, T.; Xia, D.; Zhu, J.; Zhan, X. Identification, classification, and expression analysis of the Triacylglycerol Lipase (TGL) gene family related to abiotic stresses in tomato. Int. J. Mol. Sci. 2021, 22, 1387. [Google Scholar] [CrossRef] [PubMed]
- Lafta, A.M.; Fugate, K.K. Metabolic profile of wound-induced changes in primary carbon metabolism in sugarbeet root. Phytochemistry 2011, 72, 476–489. [Google Scholar] [CrossRef]
- Zhang, Y.-G.; Kan, H.; Chen, S.-X.; Thakur, K.; Wang, S.; Zhang, J.-G.; Shang, Y.-F.; Wei, Z.-J. Comparison of phenolic compounds extracted from Diaphragma juglandis fructus, walnut pellicle, and flowers of Juglans regia using methanol, ultrasonic wave, and enzyme assisted-extraction. Food Chem. 2020, 321, 126672. [Google Scholar] [CrossRef]
- Sun, L.; Isaak, C.K.; Zhou, Y.; Petkau, J.C.; Karmin, O.; Liu, Y.; Siow, Y. Salidroside and tyrosol from Rhodiola protect H9c2 cells from ischemia/reperfusion-induced apoptosis. Life Sci. 2012, 91, 151–158. [Google Scholar] [CrossRef]
- Fan, H.; Sun, L.; Yang, L.; Zhou, J.; Yin, P.; Li, K.; Xue, Q.; Li, X.; Liu, Y. Assessment of the bioactive phenolic composition of Acer truncatum seed coat as a byproduct of seed oil. Ind. Crop. Prod. 2018, 118, 11–19. [Google Scholar] [CrossRef]
- Ministry of Environmental Protection of China. Solid waste-determination of metals-inductively coupled plasma mass spectrometry (ICP-MS). In National Environmental Protection Standards of the People’s Republic of China (HJ766–2015); China Environmental Press: Beijing, China, 2015; ISBN 135111068. [Google Scholar]
- Fan, W.; Li, B.; Tian, H.; Li, X.; Ren, H.; Zhou, Q. Metabolome and transcriptome analysis predicts metabolism of violet-red color change in Lilium bulbs. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | IC50 (μg/mL) DPPH | IC50 (μg/mL) ABTS |
|---|---|---|
| RC-L | 29.05 | 34.37 |
| RC-M | 20.06 | 24.16 |
| RC-H | 16.39 | 16.61 |
| Metabolite | Category | Fold Change (RC-H vs. RC-L) | Type | Fold Change (RC-M vs. RC-L) | Type | Fold Change (RC-H vs. RC-M) | Type |
|---|---|---|---|---|---|---|---|
| Quercetin-3-O-rutinoside (Rutin) | Flavonoids | 50,814.82 | Up | 60,233.70 | Up | N/A | N/A |
| 1-Methylpiperidine-2-carboxylic acid | Others | N/A | N/A | 63,187.78 | Up | 1.58258E-05 | Down |
| 5-O-Caffeoylshikimic acid | Phenylpropanoids | N/A | N/A | 60,226.67 | Up | 1.66039E-05 | Down |
| Rhododendrol | Phenylpropanoids | 25,852.22 | Up | 25,319.26 | Up | N/A | N/A |
| 7-O-Methxyl Quercetin (Rhamnetin) | Flavonoids | 13,547.41 | Up | 130,288.89 | Up | N/A | N/A |
| LysoPC 19:2 | Others | N/A | N/A | 15,415.56 | Up | 6.48695E-05 | Down |
| Phenylacetylglycine | Amino acids | N/A | N/A | 9512.74 | Up | 0.00011 | Down |
| 2,3,4,5,6-pentahydroxyhexyl 2-hydroxybenzoate | Phenylpropanoids | 6782.52 | Up | 4217.04 | Up | N/A | N/A |
| Catalposide | Others | N/A | N/A | 3510.22 | Up | 0.00029 | Down |
| N-Acetyl-D-glucosamine-1-phosphate | Others | 6525.93 | Up | 5938.93 | Up | N/A | N/A |
| LysoPC 20:1 | Others | N/A | N/A | 5650.63 | Up | 0.00018 | Down |
| 3-(3-Hydroxyphenyl)-propionate acid | Phenylpropanoids | N/A | N/A | 4544.74 | Up | 0.00022 | Down |
| N-Acetyl-L-methionine | Amino acids | 4257.63 | Up | N/A | N/A | 4257.63 | Up |
| S-(5′-Adenosy)-L-homocysteine | Amino acids | 3839.48 | Up | N/A | N/A | N/A | N/A |
| Tryptamine | Others | 3096.93 | Up | 20,954.07 | Up | N/A | N/A |
| 3,5-Di-O-galloylshikimic acid | Phenylpropanoids | 3020.26 | Up | 9381.78 | Up | N/A | N/A |
| Caffeoylbenzoyltartaric acid | Phenylpropanoids | 1654.26 | Up | N/A | N/A | 1654.26 | Up |
| 1,2-O-Diferuloylglycerol | Phenylpropanoids | 1494.74 | Up | N/A | N/A | 1494.74 | Up |
| 3-Aminosalicylic acid | Others | N/A | N/A | N/A | N/A | 1415.11 | Up |
| (5-L-Glutamyl)-L-amino acid | Amino acids | 1368.89 | Up | 1649.89 | Up | N/A | N/A |
| LysoPC 18:4 | Others | N/A | N/A | 1302.52 | Up | 0.0008 | Down |
| Phthalic acid | Others | 1038.84 | Up | 1834.74 | Up | N/A | N/A |
| Chlorogenic acid methyl ester | Phenylpropanoids | 868.57 | Up | N/A | N/A | 868.57 | Up |
| Sinapoyl malate | Phenylpropanoids | 565.92 | Up | 3459.89 | Up | N/A | N/A |
| Scopoletin-7-O-glucoside (Scopolin) | Phenylpropanoids | N/A | N/A | 2996.11 | Up | 0.0003 | Down |
| 4-Hydroxybenzyl Alcohol | Phenylpropanoids | 497.65 | Up | N/A | N/A | 497.65 | Up |
| 9,10-Epoxyoctadecanoic Acid | Free fatty acids and glycerides | 381.88 | Up | 1260.63 | Up | N/A | N/A |
| Indole 3-acetic acid (IAA) | Others | N/A | N/A | 716.29 | Up | 0.0014 | Down |
| Cis-4,7,10,13,16,19-Docosahexaenoic Acid | Free fatty acids and glycerides | N/A | N/A | 185.51 | Up | N/A | N/A |
| 5,7-Dimethoxycoumarin | Phenylpropanoids | N/A | N/A | 0.0028 | Down | 445.73 | Up |
| Isoeugenol | Phenylpropanoids | N/A | N/A | 0.0017 | Down | 345.22 | Up |
| 1-α-Linolenoyl-glycerol-3-O-glucoside | Free fatty acids and glycerides | N/A | N/A | 0.0014 | Down | 330.92 | Up |
| 3-Indolepropionic acid | Others | N/A | N/A | 0.0014 | Down | 3134.78 | Up |
| 3-O-p-Coumaroylquinic acid | Phenylpropanoids | N/A | N/A | 0.0012 | Down | 1371.50 | Up |
| 2′-Deoxyuridine | Nucleotides | N/A | N/A | 0.0009 | Down | 748.40 | Up |
| N-(3-Indolylacetyl)-L-alanine | Amino acids | 365.44 | Up | 2071.70 | Up | N/A | N/A |
| Methyl caffeate | Phenylpropanoids | 147.26 | Up | N/A | N/A | 147.26 | Up |
| Clove chromone | Others | 0.00085 | Down | 0.0009 | Down | N/A | N/A |
| LysoPE 15:0(2n isomer) | Others | 0.00075 | Down | 0.0008 | Down | N/A | N/A |
| 4-Pyridoxic acid-O-glucoside | Others | N/A | N/A | 0.0006 | Down | 931.35 | Up |
| Quercetin-3-O-(6′’-galloyl)galactoside | Flavonoids | N/A | N/A | 0.0001 | Down | 7396.52 | Up |
| 2-Aminopurine | Nucleotides | N/A | N/A | 6.617E-05 | Down | 4106.22 | Up |
| 1,6-Di-O-Galloyl-D-Glucose | Gallic acid derivatives | N/A | N/A | 1.17381E-05 | Down | 131,281.48 | Up |
| LysoPC 15:0 | Others | 0.00059 | Down | N/A | N/A | 0.00028 | Down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, T.; Sha, Y.; Liu, H.; Sun, L. Altitudinal Variation of Metabolites, Mineral Elements and Antioxidant Activities of Rhodiola crenulata (Hook.f. & Thomson) H.Ohba. Molecules 2021, 26, 7383. https://doi.org/10.3390/molecules26237383
Dong T, Sha Y, Liu H, Sun L. Altitudinal Variation of Metabolites, Mineral Elements and Antioxidant Activities of Rhodiola crenulata (Hook.f. & Thomson) H.Ohba. Molecules. 2021; 26(23):7383. https://doi.org/10.3390/molecules26237383
Chicago/Turabian StyleDong, Tingting, Yueqi Sha, Hairong Liu, and Liwei Sun. 2021. "Altitudinal Variation of Metabolites, Mineral Elements and Antioxidant Activities of Rhodiola crenulata (Hook.f. & Thomson) H.Ohba" Molecules 26, no. 23: 7383. https://doi.org/10.3390/molecules26237383
APA StyleDong, T., Sha, Y., Liu, H., & Sun, L. (2021). Altitudinal Variation of Metabolites, Mineral Elements and Antioxidant Activities of Rhodiola crenulata (Hook.f. & Thomson) H.Ohba. Molecules, 26(23), 7383. https://doi.org/10.3390/molecules26237383