Pineapple Leaf Phenols Attenuate DSS-Induced Colitis in Mice and Inhibit Inflammatory Damage by Targeting the NF-κB Pathway


 ,
,
Abstract
:1. Introduction
2. Materials and methods
2.1. Preparation of Extracts and Active Components
2.2. DSS-Induced Colitis Mouse Models
2.3. Disease Activity Index (DAI) Assay
2.4. Histological Analysis
2.5. MPO Activity Assay
2.6. Cell Culture
2.7. In Vitro Scratch Assay
2.8. Inflammation Induction
2.9. Western Blotting Analysis
2.10. Reverse Transcription and Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) Assay
2.11. Statistical Analysis
3. Results
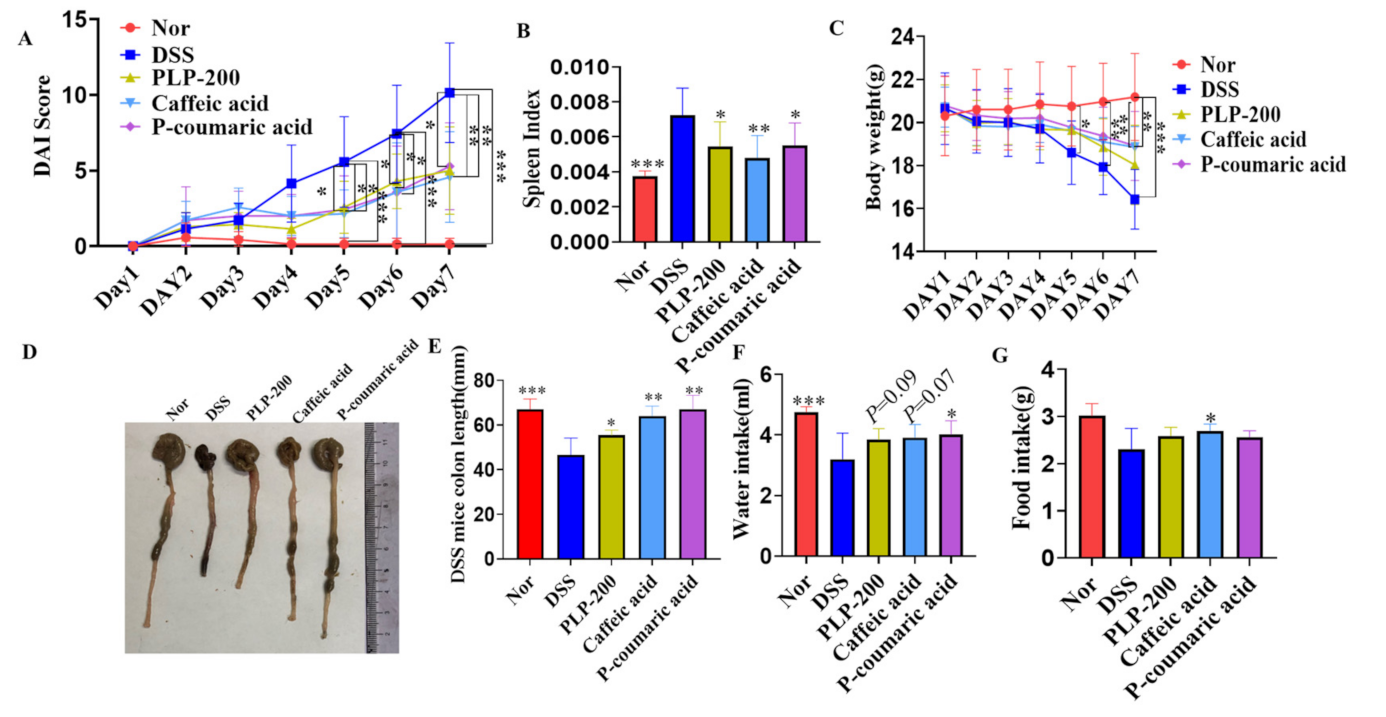
3.1. Treatment with PLPs and Their Monomers Caffeic Acid and P-Coumaric Acid Ameliorates the Symptoms of DSS-Induced Experimental Colitis in Mice
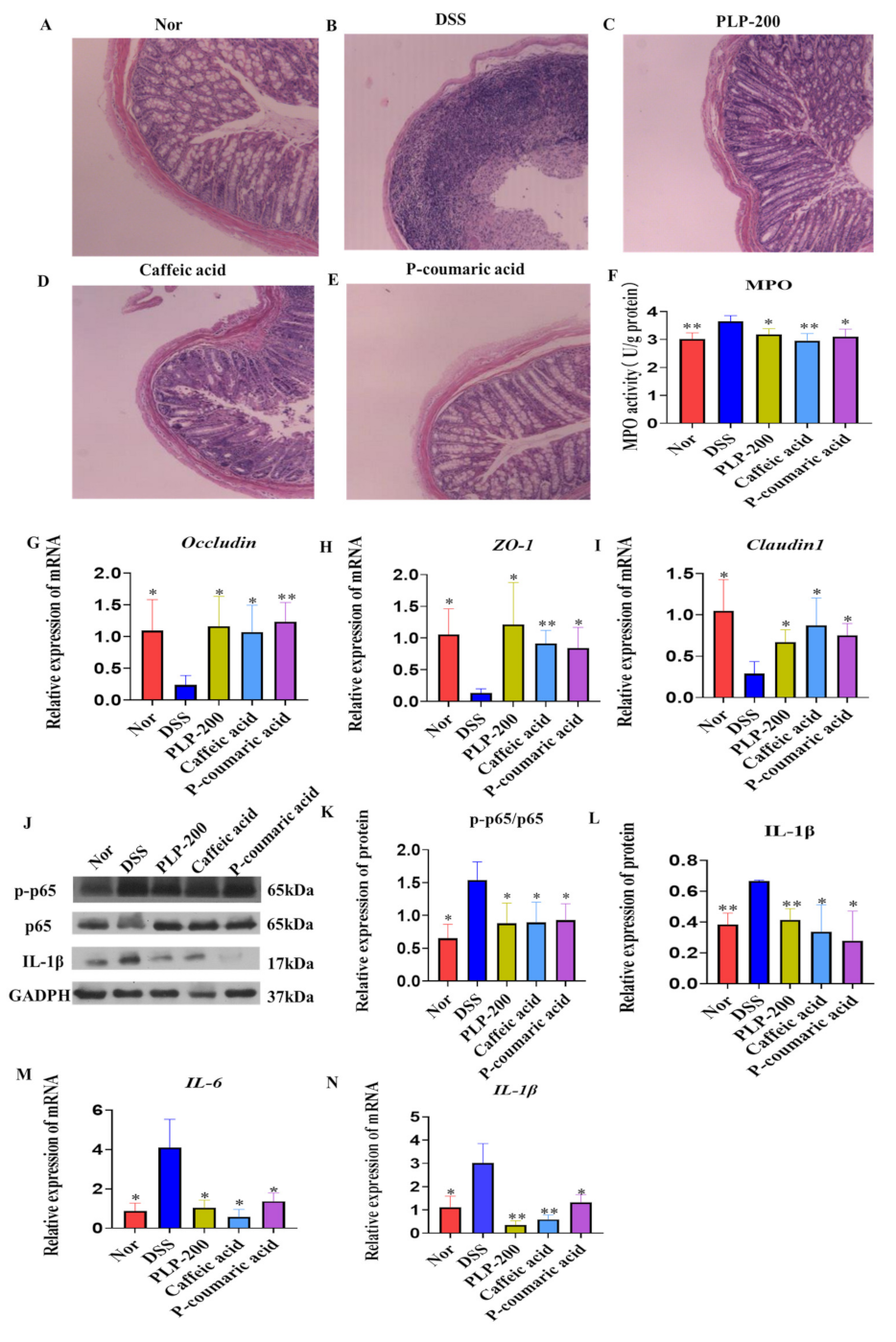
3.2. PLPs, Caffeic Acid, and P-Coumaric Acid Administration Reduces Histopathological Damage and Inflammatory Response in DSS-Induced Colitis
3.3. PLPs, Caffeic Acid, and P-Coumaric Acid Inhibit the Secretion of Pro-Inflammatory Factors and the Activation of NF-κB Cell Signaling in LPS-Stimulated RAW264.7 Cells
3.4. PLPs, Caffeic Acid, and P-Coumaric Acid Accelerate Wound Closure and Increase the Gene Expression of Occludin, ZO-1, and Claudin 1 in Caco-2 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Baumgart, D.C.; Sandborn, W.J. Inflammatory bowel disease: Clinical aspects and established and evolving therapies. Lancet 2007, 369, 1641–1657. [Google Scholar] [CrossRef]
- Loftus, E.V., Jr. Clinical epidemiology of inflammatory bowel disease: Incidence, prevalence, and environmental influences. Gastroenterology 2004, 126, 1504–1517. [Google Scholar] [CrossRef]
- Kaser, A.; Zeissig, S.; Blumberg, R.S. Inflammatory bowel disease. Annu. Rev. Immunol. 2010, 28, 573–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza, H.S.P.; Fiocchi, C. Immunopathogenesis of IBD: Current state of the art. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 13–27. [Google Scholar] [CrossRef]
- Dignass, A.; Lindsay, J.O.; Sturm, A.; Windsor, A.; Colombel, J.-F.; Allez, M.; D’Haens, G.; D’Hoore, A.; Mantzaris, G.; Novacek, G.; et al. Second European evidence-based consensus on the diagnosis and management of ulcerative colitis Part 2: Current management. J. Crohn’s Colitis 2012, 6, 991–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, T.; Kim, D.H.; Lim, B.O. Natural Products as a Source of Anti-Inflammatory Agents Associated with Inflammatory Bowel Disease. Molecules 2013, 18, 7253–7270. [Google Scholar] [CrossRef]
- Sun, Y.; Zhao, Y.; Yao, J.; Zhao, L.; Wu, Z.; Wang, Y.; Pan, D.; Miao, H.; Guo, Q.; Lu, N. Wogonoside protects against dextran sulfate sodium-induced experimental colitis in mice by inhibiting NF-kappaB and NLRP3 inflammasome activation. Biochem. Pharm. 2015, 94, 142–154. [Google Scholar] [CrossRef]
- Upadhyay, A.; Upadhyaya, I.; Johny, A.K.; Venkitanarayanan, K. Combating Pathogenic Microorganisms Using Plant-Derived Antimicrobials: A Minireview of the Mechanistic Basis. BioMed Res. Int. 2014, 2014, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Wang, W.; Su, H.; Xing, D.; Cai, G.; Du, L. Hypolipidemic mechanisms of Ananas comosus L. leaves in mice: Different from fibrates but similar to statins. J. Pharmacol. Sci. 2007, 103, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Wang, W.; Su, H.; Xing, D.; Pan, Y.; Du, L. Effect of ethanolic extracts of Ananas comosus L. leaves on insulin sensitivity in rats and HepG2. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 143, 429–435. [Google Scholar] [CrossRef]
- Xie, W.; Zhang, S.; Lei, F.; Ouyang, X.; Du, L. Ananas comosusL. Leaf Phenols and p-Coumaric Acid Regulate Liver Fat Metabolism by Upregulating CPT-1 Expression. Evidence-Based Complement. Altern. Med. 2014, 2014, 1–12. [Google Scholar] [CrossRef]
- Xie, W.; Xing, D.; Sun, H.; Wang, W.; Ding, Y.; Du, L. The effects of Ananas comosus L. leaves on diabetic-dyslipidemic rats induced by alloxan and a high-fat/high-cholesterol diet. Am. J. Chin. Med. 2005, 33, 95–105. [Google Scholar] [CrossRef]
- Dai, G.; Jiang, Z.; Sun, B.; Liu, C.; Meng, Q.; Ding, K.; Jing, W.; Ju, W. Caffeic Acid Phenethyl Ester Prevents Colitis-Associated Cancer by Inhibiting NLRP3 Inflammasome. Front. Oncol. 2020, 10, 721. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wu, X.; Cao, S.; Wang, L.; Wang, D.; Yang, H.; Feng, Y.; Wang, S.; Li, L. Caffeic acid ameliorates colitis in association with increased Akkermansia population in the gut microbiota of mice. Oncotarget 2016, 7, 31790. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; You, Y.; Xi, Y.; Ni, B.; Chu, X.; Zhang, R.; You, H. p-Coumaric Acid Attenuates IL-1beta-Induced Inflammatory Responses and Cellular Senescence in Rat Chondrocytes. Inflammation 2020, 43, 619–628. [Google Scholar] [CrossRef]
- Pei, K.; Ou, J.; Huang, J.; Ou, S. p-Coumaric acid and its conjugates: Dietary sources, pharmacokinetic properties and biological activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef]
- Randhawa, P.K.; Singh, K.; Singh, N.; Jaggi, A.S. A Review on Chemical-Induced Inflammatory Bowel Disease Models in Rodents. Korean J. Physiol. Pharmacol. 2014, 18, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Blanco, J.A.; Estévez, J.; Shea-Donohue, T.; Martínez, V.; Vergara, P. Changes in Epithelial Barrier Function in Response to Parasitic Infection: Implications for IBD Pathogenesis. J. Crohn’s Colitis 2015, 9, 463–476. [Google Scholar] [CrossRef] [Green Version]
- Holmberg, F.E.; Seidelin, J.; Yin, X.; E Mead, B.; Tong, Z.; Li, Y.; Karp, J.M.; Nielsen, O.H. Culturing human intestinal stem cells for regenerative applications in the treatment of inflammatory bowel disease. EMBO Mol. Med. 2017, 9, 558–570. [Google Scholar] [CrossRef]
- Li, P.; Zheng, Y.; Chen, X. Drugs for Autoimmune Inflammatory Diseases: From Small Molecule Compounds to Anti-TNF Biologics. Front. Pharmacol. 2017, 8, 460. [Google Scholar] [CrossRef] [PubMed]
- Bazzoni, G.; Dejana, E. Endothelial cell-to-cell junctions: Molecular organization and role in vascular homeostasis. Physiol. Rev. 2004, 84, 869–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siliciano, J.D.; Goodenough, D.A. Localization of the tight junction protein, ZO-1, is modulated by extracellular calcium and cell-cell contact in Madin-Darby canine kidney epithelial cells. J. Cell Biol. 1988, 107, 2389–2399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Guo, Y. Supplemental zinc reduced intestinal permeability by enhancing occludin and zonula occludens protein-1 (ZO-1) expression in weaning piglets. Br. J. Nutr. 2009, 102, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Heller, F.; Florian, P.; Bojarski, C.; Richter, J.; Christ, M.; Hillenbrand, B.; Mankertz, J.; Gitter, A.H.; Burgel, N.; Fromm, M.; et al. Interleukin-13 is the key effector Th2 cytokine in ulcerative colitis that affects epithelial tight junctions, apoptosis, and cell restitution. Gastroenterology 2005, 129, 550–564. [Google Scholar] [CrossRef]
- Zeissig, S.; Bürgel, N.; Günzel, D.; Richter, J.; Mankertz, J.; Wahnschaffe, U.; Kroesen, A.J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut 2007, 56, 61–72. [Google Scholar] [CrossRef]
- Nunes, C.; Almeida, L.; Barbosa, R.M.; Laranjinha, J. Luteolin suppresses the JAK/STAT pathway in a cellular model of intestinal inflammation. Food Funct. 2017, 8, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.; Freitas, V.; Almeida, L.; Laranjinha, J. Red wine extract preserves tight junctions in intestinal epithelial cells under inflammatory conditions: Implications for intestinal inflammation. Food Funct. 2019, 10, 1364–1374. [Google Scholar] [CrossRef]
- Christian, F.; Smith, E.L.; Carmody, R.J. The Regulation of NF-κB Subunits by Phosphorylation. Cells 2016, 5, 12. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | NCBI | Size |
|---|---|---|---|---|
| β-Actin (Mouse) | GTGACGTTGACATCCGTAAAGA | GCCGGACTCATCGTACTCC | NM_007393 | 245 |
| IL-1β (Mouse) | GCAACTGTTCCTGAACTCAACT | ATCTTTTGGGGTCCGTCAACT | NM_008361 | 89 |
| IL-6 (Mouse) | CTGCAAGAGACTTCCATCCAG | GAGTGGTATAGACAGGTCTGTTGG | NM_031168 | 131 |
| TNF-α (Mouse) | GGGCTTCCAGAACTCCA | GCTACAGGCTTGTCACTCG | NM_013693.2 | 213 |
| ZO-1 (Mouse) | GCCGCTAAGAGCACAGCAA | TCCCCACTCTGAAAATGAGGA | NM_001163574 | 134 |
| Claudin 1 (Mouse) | GGGGACAACATCGTGACCG | AGGAGTCGAAGACTTTGCACT | NM_016674 | 100 |
| Occluding (Mouse) | TTGAAAGTCCACCTCCTTACAGA | CCGGATAAAAAGAGTACGCTGG | NM_008756 | 129 |
| β-Actin (Human) | CATGTACGTTGCTATCCAGGC | CTCCTTAATGTCACGCACGAT | NM_001101 | 250 |
| ZO-1 (Human) | CAACATACAGTGACGCTTCACA | CACTATTGACGTTTCCCCACTC | NM_003257 | 105 |
| Claudin 1 (Human) | CCTCCTGGGAGTGATAGCAAT | GGCAACTAAAATAGCCAGACCT | NM_021101 | 145 |
| Occluding (Human) | ACAAGCGGTTTTATCCAGAGTC | GTCATCCACAGGCGAAGTTAAT | NM_001205254 | 89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Niu, Y.; Hao, W.; Zhang, W.; Lu, J.; Zhou, J.; Du, L.; Xie, W. Pineapple Leaf Phenols Attenuate DSS-Induced Colitis in Mice and Inhibit Inflammatory Damage by Targeting the NF-κB Pathway. Molecules 2021, 26, 7656. https://doi.org/10.3390/molecules26247656
Chen Y, Niu Y, Hao W, Zhang W, Lu J, Zhou J, Du L, Xie W. Pineapple Leaf Phenols Attenuate DSS-Induced Colitis in Mice and Inhibit Inflammatory Damage by Targeting the NF-κB Pathway. Molecules. 2021; 26(24):7656. https://doi.org/10.3390/molecules26247656
Chicago/Turabian StyleChen, Yang, Yaoyun Niu, Wenhui Hao, Wanqiu Zhang, Jinghua Lu, Jin Zhou, Lijun Du, and Weidong Xie. 2021. "Pineapple Leaf Phenols Attenuate DSS-Induced Colitis in Mice and Inhibit Inflammatory Damage by Targeting the NF-κB Pathway" Molecules 26, no. 24: 7656. https://doi.org/10.3390/molecules26247656
APA StyleChen, Y., Niu, Y., Hao, W., Zhang, W., Lu, J., Zhou, J., Du, L., & Xie, W. (2021). Pineapple Leaf Phenols Attenuate DSS-Induced Colitis in Mice and Inhibit Inflammatory Damage by Targeting the NF-κB Pathway. Molecules, 26(24), 7656. https://doi.org/10.3390/molecules26247656