
Kahweol, a Diterpenoid Molecule, Inhibits CTGF-Dependent Synthetic Phenotype Switching and Migration in Vascular Smooth Muscle Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
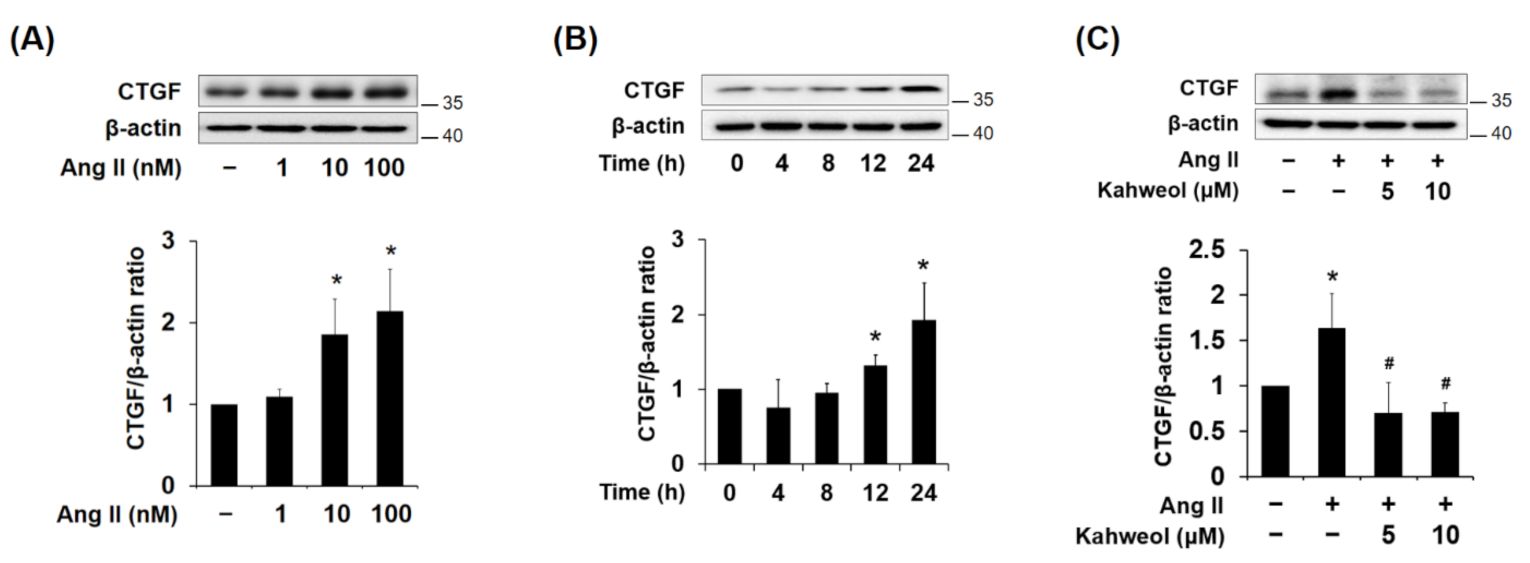
2.1. Kahweol Reduces Ang II-Induced CTGF Protein Expression in VSMCs
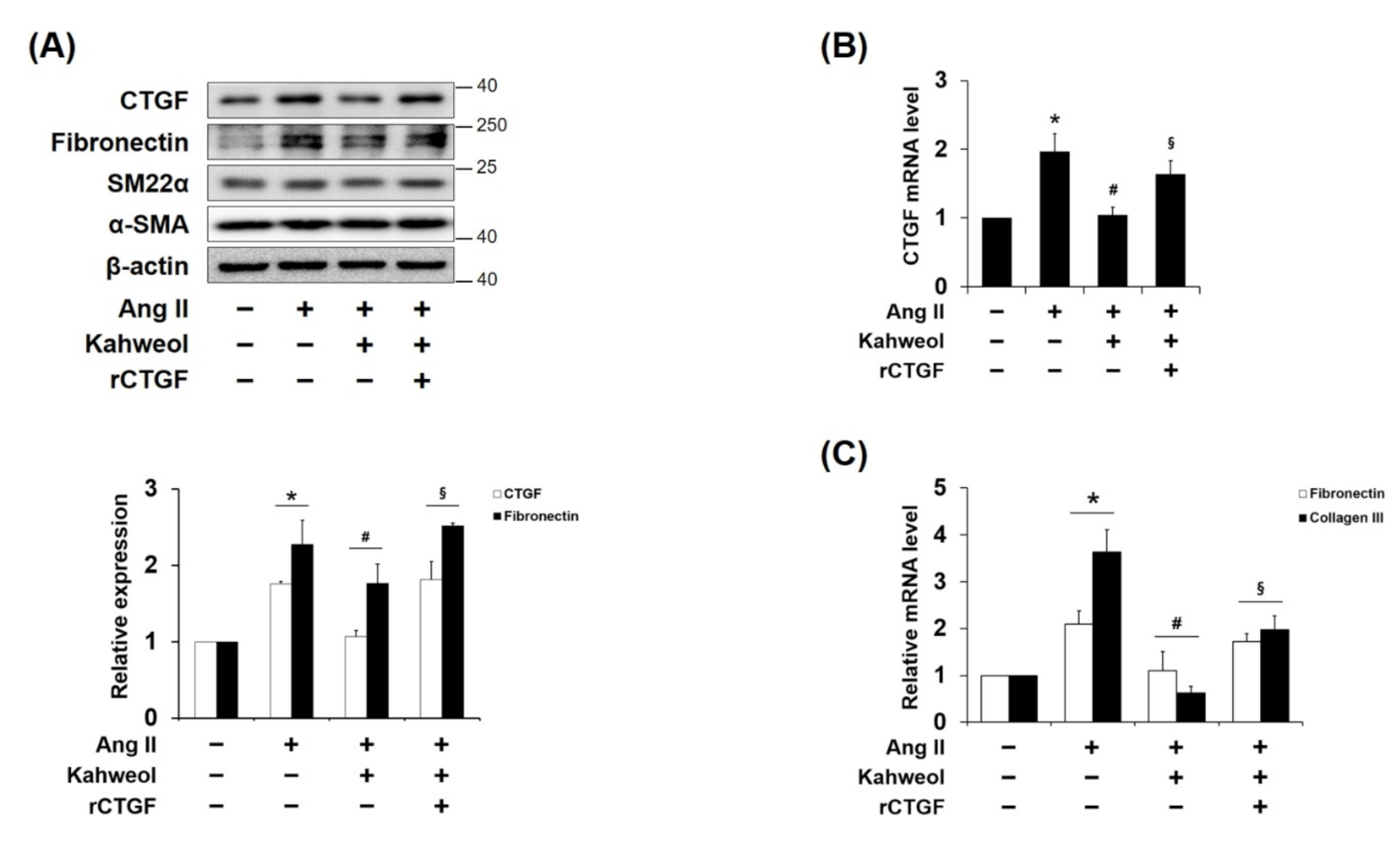
2.2. Kahweol Inhibits Ang II-Induced CTGF and Synthetic Phenotype Marker Expressions, and rCTGF Reverses the Kahweol Effect in VSMCs
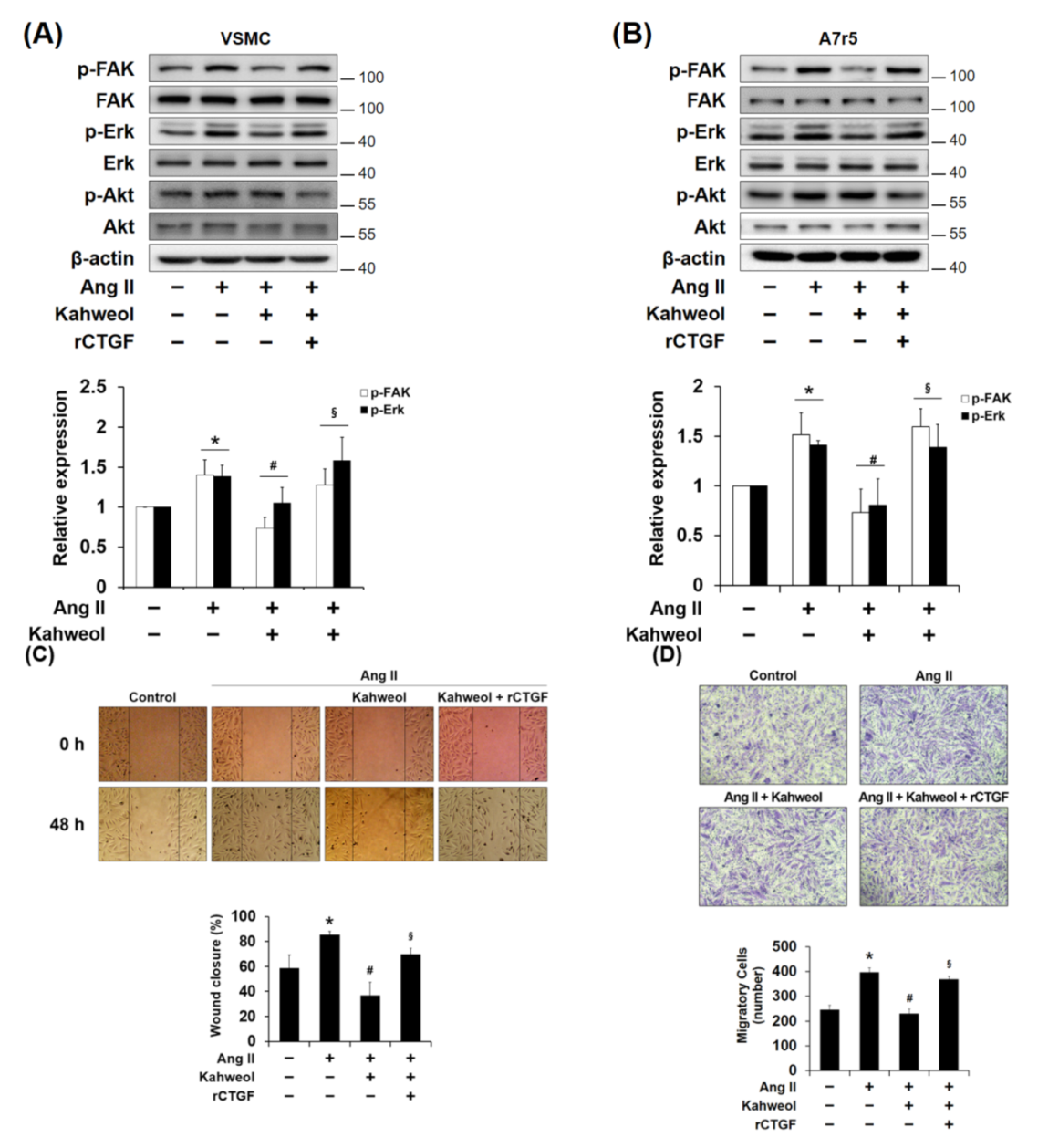
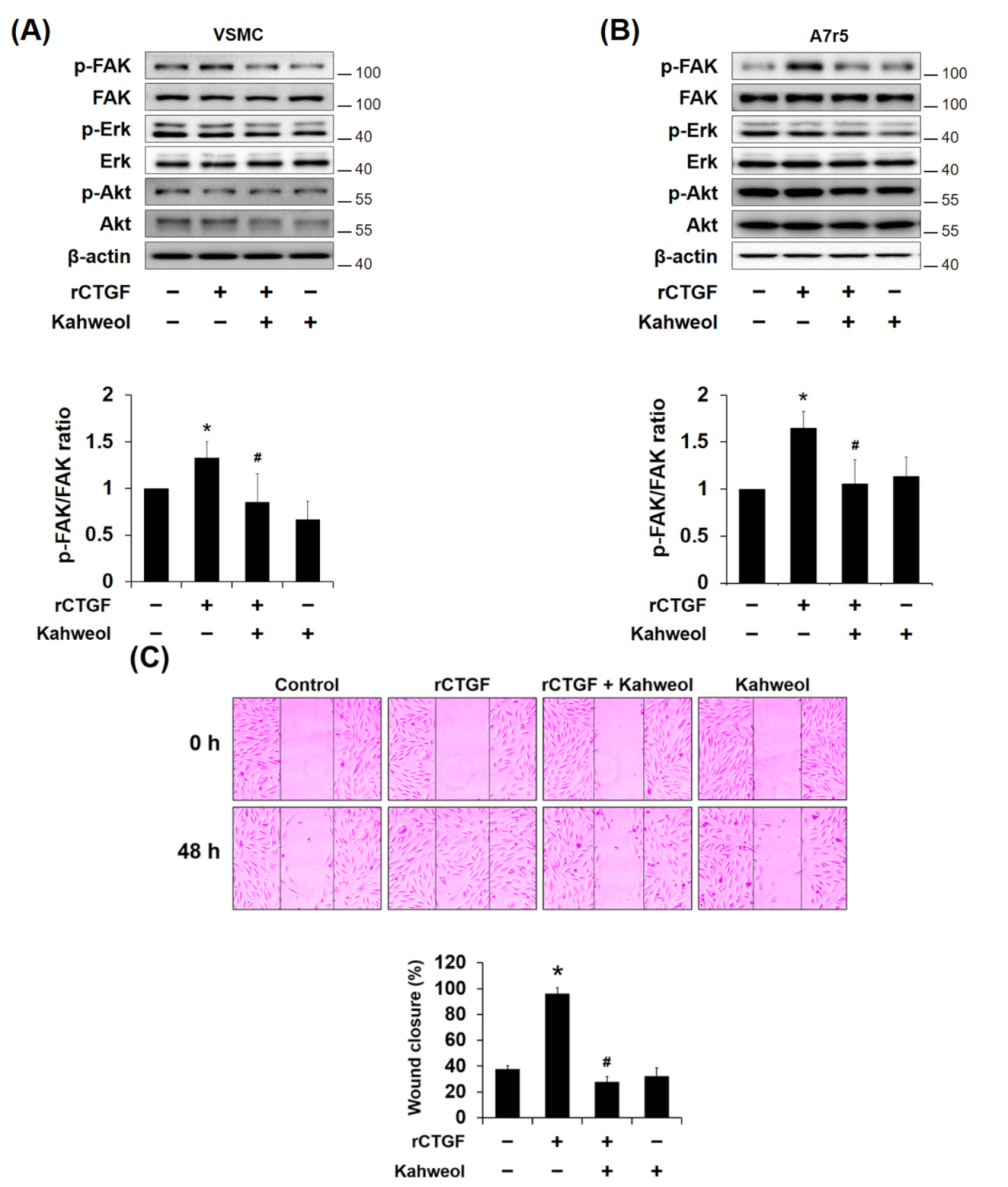
2.3. Kahweol Inhibits Ang II-Induced FAK and Erk Phosphorylation and Migration, and rCTGF Reverses the Kahweol Effects in VSMCs
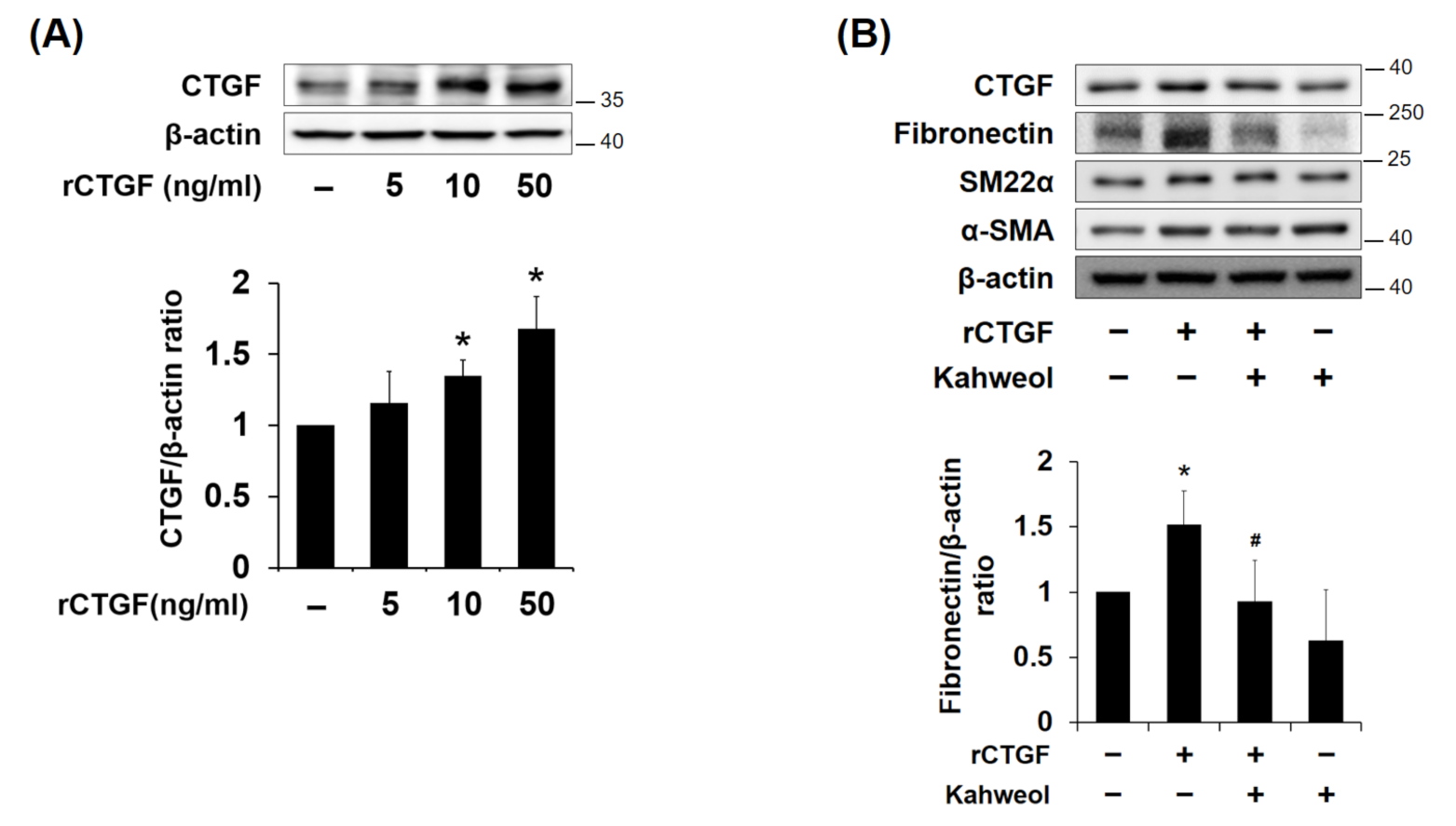
2.4. Kahweol Inhibits Fibronectin Expression by Reducing CTGF Expression in VSMCs
2.5. Kahweol Inhibits CTGF-Induced FAK Phosphorylation and Migration in VSMCs
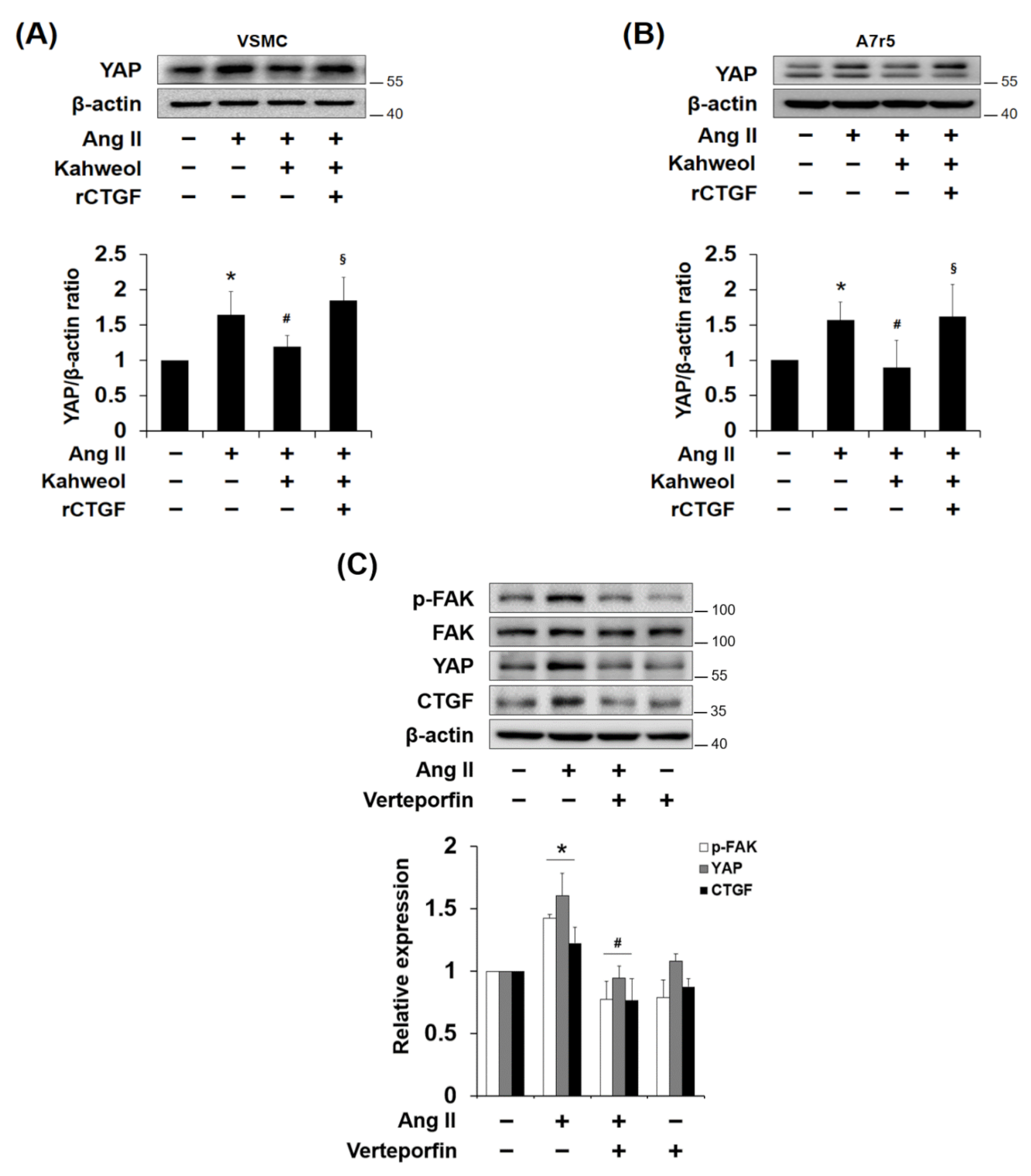
2.6. Kahweol Inhibits Ang II-Induced YAP Expression, and rCTGF Reverses the Kahweol Effect in VSMCs
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Primary Cell Culture and Drug Treatment
4.3. Western Blot Analysis
4.4. Quantitative Real-Time RT-PCR (qRT-PCR)
4.5. Wound Scratch Assay and Transwell Chamber Assay
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Sample Availability
References
- Mathieu, P.; Pibarot, P.; Despres, J.P. Metabolic Syndrome: The Danger Signal in Atherosclerosis. Vasc. Health Risk Manag. 2006, 2, 285–302. [Google Scholar] [CrossRef] [Green Version]
- Rafieian-Kopaei, M.; Setorki, M.; Doudi, M.; Baradaran, A.; Nasri, H. Atherosclerosis: Process, indicators, risk factors and new hopes. Int. J. Prev. Med. 2014, 5, 927–946. [Google Scholar] [PubMed]
- Bergheanu, S.C.; Bodde, M.C.; Jukema, J.W. Pathophysiology and treatment of atherosclerosis: Current view and future perspective on lipoprotein modification treatment. Neth. Heart J. 2017, 25, 231–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basatemur, G.L.; Jørgensen, H.F.; Clarke, M.; Bennett, M.R.; Mallat, Z. Vascular smooth muscle cells in atherosclerosis. Nat. Rev. Cardiol. 2019, 16, 727–744. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular Smooth Muscle Cells in Atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Vascular smooth muscle cell in atherosclerosis. Acta Physiol. 2015, 214, 33–50. [Google Scholar] [CrossRef]
- Louis, S.F.; Zahradka, P. Vascular smooth muscle cell motility: From migration to invasion. Exp. Clin. Cardiol. 2010, 15, e75–e85. [Google Scholar]
- Hu, D.; Yin, C.; Luo, S.; Habenicht, A.; Mohanta, S.K. Vascular Smooth Muscle Cells Contribute to Atherosclerosis Immunity. Front. Immunol. 2019, 10, 1101. [Google Scholar] [CrossRef]
- Chang, S.; Song, S.; Lee, J.; Yoon, J.; Park, J.; Choi, S.; Park, J.K.; Choi, K.; Choi, C. Phenotypic modulation of primary vascular smooth muscle cells by short-term culture on micropatterned substrate. PLoS ONE 2014, 9, e88089. [Google Scholar] [CrossRef]
- Dong, X.; Yu, L.; Sun, R.; Cheng, Y.; Cao, H.; Yang, K.; Dong, Y.; Wu, Y.; Guo, X. Inhibition of PTEN expression and activity by angiotensin II induces proliferation and migration of vascular smooth muscle cells. J. Cell Biochem. 2013, 114, 174–182. [Google Scholar] [CrossRef]
- Moraes, J.A.; Frony, A.C.; Dias, A.M.; Renovato-Martins, M.; Rodrigues, G.; Marcinkiewicz, C.; Assreuy, J.; Barja-Fidalgo, C. A1β1 and integrin-linked kinase interact and modulate angiotensin II effects in vascular smooth muscle cells. Atherosclerosis 2015, 243, 477–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.H.; Hwang, Y.P.; Choi, J.H.; Jin, S.W.; Lee, G.H.; Han, E.H.; Chung, Y.H.; Chung, Y.C.; Jeong, H.G. Kahweol inhibits proliferation and induces apoptosis by suppressing fatty acid synthase in HER2-overexpressing cancer cells. Food Chem. Toxicol. 2018, 121, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Bang, W.; Cho, J.H.; Lee, R.H.; Kim, S.H.; Kim, M.S.; Park, S.M.; Shin, J.C.; Chung, H.J.; Oh, K.B.; et al. Kahweol induces apoptosis by suppressing BTF3 expression through the ERK signaling pathway in non-small cell lung cancer cells. Int. J. Oncol. 2016, 49, 2294–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardenas, C.; Quesada, A.R.; Medina, M.A. Anti-Angiogenic and anti-inflammatory properties of kahweol, a coffee diterpene. PLoS ONE 2011, 6, e23407. [Google Scholar] [CrossRef]
- Seo, H.Y.; Kim, M.K.; Lee, S.H.; Hwang, J.S.; Park, K.G.; Jang, B.K. Kahweol Ameliorates the Liver Inflammation through the Inhibition of NF-κB and STAT3 Activation in Primary Kupffer Cells and Primary Hepatocytes. Nutrients 2018, 10, 863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.Y.; Jung, Y.A.; Lee, S.H.; Hwang, J.S.; Park, K.G.; Kim, M.K.; Jang, B.K. Kahweol decreases hepatic fibrosis by inhibiting the expression of connective tissue growth factor via the transforming growth factor-beta signaling pathway. Oncotarget 2017, 8, 87086–87094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.J.; Choi, J.H.; Jeong, H.G. Hepatoprotective and antioxidant effects of the coffee diterpenes kahweol and cafestol on carbon tetrachloride-induced liver damage in mice. Food Chem. Toxicol. 2007, 45, 2118–2125. [Google Scholar] [CrossRef] [PubMed]
- Lipson, K.E.; Wong, C.; Teng, Y.; Spong, S. CTGF is a central mediator of tissue remodeling and fibrosis and its inhibition can reverse the process of fibrosis. Fibrogenesis Tissue Repair 2012, 5, S24. [Google Scholar] [CrossRef] [Green Version]
- Perbal, B. CCN proteins: Multifunctional signalling regulators. Lancet 2004, 363, 62–64. [Google Scholar] [CrossRef]
- Rachfal, A.W.; Brigstock, D.R. Structural and functional properties of CCN proteins. Vitam. Horm. 2005, 70, 69–103. [Google Scholar]
- Ruperez, M.; Lorenzo, O.; Blanco-Colio, L.M.; Esteban, V.; Egido, J.; Marta Ruiz-Ortega, M. Connective tissue growth factor is a mediator of angiotensin II-induced fibrosis. Circulation 2003, 108, 1499–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi-Wen, X.; Leask, A.; Abraham, D. Regulation and function of connective tissue growth factor/CCN2 in tissue repair, scarring and fibrosis. Cytokine Growth Factor Rev. 2008, 19, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, D.P.; Farias, G.C.; Sousa, E.B.; Coelho-Aguiar, J.M.; Lobo, J.C.; Casado, P.L.; Duart, M.E.; Abreu, J.G. New strategy to control cell migration and metastasis regulated by CCN2/CTGF. Cancer Cell Int. 2014, 14, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Abraham, D.J.; Shi-Wen, X.; Pearson, J.D.; Black, C.M.; Lyons, K.M.; Leask, A. CCN2 (connective tissue growth factor) promotes fibroblast adhesion to fibronectin. Mol. Biol. Cell 2004, 15, 5635–5646. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.C.; Chen, B.C.; Hsu, M.J.; Tsai, C.T.; Hong, C.Y.; Lin, C.H. Thrombin induced connective tissue growth factor expression in rat vascular smooth muscle cells via the PAR-1/JNK/AP-1 pathway. Acta Pharmacol. Sin. 2012, 33, 49–56. [Google Scholar] [CrossRef]
- Hwang, A.R.; Nam, J.O.; Kang, Y.J. Fluvastatin inhibits advanced glycation end products-induced proliferation, migration, and extracellular matrix accumulation in vascular smooth muscle cells by targeting connective tissue growth factor. Korean J. Physiol. Pharmacol. 2018, 22, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Cicha, I.; Yilmaz, A.; Klein, M.; Raithel, D.; Brigstock, D.R.; Daniel, W.G.; Goppelt-Struebe, M.; Garlichs, C.D. Connective tissue growth factor is overexpressed in complicated atherosclerotic plaques and induces mononuclear cell chemotaxis in vitro. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1008–1013. [Google Scholar] [CrossRef]
- Fan, W.H.; Pech, M.; Karnovsky, M.J. Connective tissue growth factor (CTGF) stimulates vascular smooth muscle cell growth and migration in vitro. Eur. J. Cell Biol. 2000, 79, 915–923. [Google Scholar] [CrossRef]
- Branchetti, E.; Poggio, P.; Sainger, R.; Shang, E.; Grau, J.B.; Jackson, B.M.; Lai, E.K.; Parmacek, M.S.; Gorman, R.C.; Gorman, J.H.; et al. Oxidative stress modulates vascular smooth muscle cell phenotype via CTGF in thoracic aortic aneurysm. Cardiovasc. Res. 2013, 100, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Lachowski, D.; Cortes, E.; Robinson, B.; Rice, A.; Rombouts, K.; Hernandez, A.E. FAK controls the mechanical activation of YAP, a transcriptional regulator required for durotaxis. FASEB J. 2018, 32, 1099–1107. [Google Scholar] [CrossRef] [Green Version]
- Ha, Y.M.; Lee, D.H.; Kim, M.; Kang, Y.J. High glucose induces connective tissue growth factor expression and extracellular matrix accumulation in rat aorta vascular smooth muscle cells via extracellular signal-regulated kinase 1/2. Korean J. Physiol. Pharmacol. 2013, 17, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Schmidt-Ott, K.M.; Kagiyama, S.; Phillips, M.I. The multiple actions of angiotensin II in atherosclerosis. Regul. Pept. 2000, 93, 65–77. [Google Scholar] [CrossRef]
- Weiss, D.; Sorescu, D.; Taylor, W.R. Angiotensin II and atherosclerosis. Am. J. Cardiol. 2001, 87, 25C–32C. [Google Scholar] [CrossRef]
- Riser, D.; Riser, B.L.; Denichilo, M.; Cortes, P.; Baker, C.; Grondin, J.M.; Yee, J.; Narins, R.G. Regulation of connective tissue growth factor activity in cultured rat mesangial cells and its expression in experimental diabetic glomerulosclerosis. J. Am. Soc. Nephrol. 2000, 11, 25–38. [Google Scholar]
- Tong, L.; Qi, G. Crocin prevents plateletderived growth factor BBinduced vascular smooth muscle cells proliferation and phenotypic switch. Mol. Med. Rep. 2018, 17, 7595–7602. [Google Scholar] [PubMed] [Green Version]
- Iwamoto, H.; Izumi, K.; Natsagdorj, A.; Naito, R.; Makino, T.; Kadomoto, S.; Hiratsuka, K.; Shigehara, K.; Kadono, Y.; Narimoto, K.; et al. Coffee diterpenes kahweol acetate and cafestol synergistically inhibit the proliferation and migration of prostate cancer cells. Prostate 2019, 79, 468–479. [Google Scholar] [CrossRef]
- Lopez-Colome, A.M.; Lee-Rivera, I.; Benavides-Hidalgo, R.; Lopez, E. Paxillin: A crossroad in pathological cell migration. J. Hematol. Oncol. 2017, 10, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, J.T.; Martin, K.H.; Slack, J.K.; Taylor, J.M.; Weed, S.A. Focal adhesion kinase: A regulator of focal adhesion dynamics and cell movement. Oncogene 2000, 19, 5606–5613. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Zaal, E.A.; Berkers, C.R.; Lemeer, S.; Heck, A.J. CTGF/VEGFA-Activated Fibroblasts Promote Tumor Migration through Micro-Environmental Modulation. Mol. Cell Proteom. 2018, 17, 1502–1514. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.C.; Hsu, C.J.; Fong, Y.C.; Chuang, S.M.; Tang, C.H. CTGF induces monocyte chemoattractant protein-1 expression to enhance monocyte migration in human synovial fibroblasts. Biochim. Biophys. Acta 2013, 1833, 1114–1124. [Google Scholar] [CrossRef] [Green Version]
- Boopathy, G.T.K.; Hong, W. Role of Hippo Pathway-YAP/TAZ Signaling in Angiogenesis. Front. Cell Dev. Biol. 2019, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Bao, Q.; Yan, M.; Liang, J.; Zhu, Y.; Wang, C.; Ai, D. The role of Hippo/yes-associated protein signalling in vascular remodelling associated with cardiovascular disease. Br. J. Pharmacol. 2018, 175, 1354–1361. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Yuan, W.; Su, Z.; Lin, C.; Huang, T.; Chen, Y.; Wang, J. Yes-Associated protein mediates angiotensin II-induced vascular smooth muscle cell phenotypic modulation and hypertensive vascular remodelling. Cell Prolif. 2018, 51, e12517. [Google Scholar] [CrossRef] [Green Version]
- Nardone, G.; Cruz, J.O.; Vrbsky, J.; Martini, C.; Pribyl, J.; Skladal, P.; Pesl, M.; Caluori, G.; Pagliari, S.; Martino, F.; et al. YAP regulates cell mechanics by controlling focal adhesion assembly. Nat. Commun. 2017, 8, 15321. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Cao, B.; Wang, Y.; Ma, C.; Zeng, Z.; Liu, L.; Li, X.; Tao, D.; Gong, J.; Xie, D. Hippo component YAP promotes focal adhesion and tumour aggressiveness via transcriptionally activating THBS1/FAK signalling in breast cancer. J. Exp. Clin. Cancer Res. 2018, 37, 175. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Jin, G.N.; Ma, M.; Liang, H.F.; Zhang, B.X.; Chen, X.P.; Ding, Z.Y. Taurocholate Induces Connective Tissue Growth Factor Expression in Hepatocytes Through ERK-YAP Signaling. Cell Physiol. Biochem. 2018, 50, 1711–1725. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.H.; Choi, S.T.; Kang, Y.J. Kahweol, a Diterpenoid Molecule, Inhibits CTGF-Dependent Synthetic Phenotype Switching and Migration in Vascular Smooth Muscle Cells. Molecules 2021, 26, 640. https://doi.org/10.3390/molecules26030640
Lee JH, Choi ST, Kang YJ. Kahweol, a Diterpenoid Molecule, Inhibits CTGF-Dependent Synthetic Phenotype Switching and Migration in Vascular Smooth Muscle Cells. Molecules. 2021; 26(3):640. https://doi.org/10.3390/molecules26030640
Chicago/Turabian StyleLee, Jeong Hee, Seok Tae Choi, and Young Jin Kang. 2021. "Kahweol, a Diterpenoid Molecule, Inhibits CTGF-Dependent Synthetic Phenotype Switching and Migration in Vascular Smooth Muscle Cells" Molecules 26, no. 3: 640. https://doi.org/10.3390/molecules26030640
APA StyleLee, J. H., Choi, S. T., & Kang, Y. J. (2021). Kahweol, a Diterpenoid Molecule, Inhibits CTGF-Dependent Synthetic Phenotype Switching and Migration in Vascular Smooth Muscle Cells. Molecules, 26(3), 640. https://doi.org/10.3390/molecules26030640