
Are the Closely Related Cobetia Strains of Different Species?

 , ,
, ,
Abstract
:1. Introduction
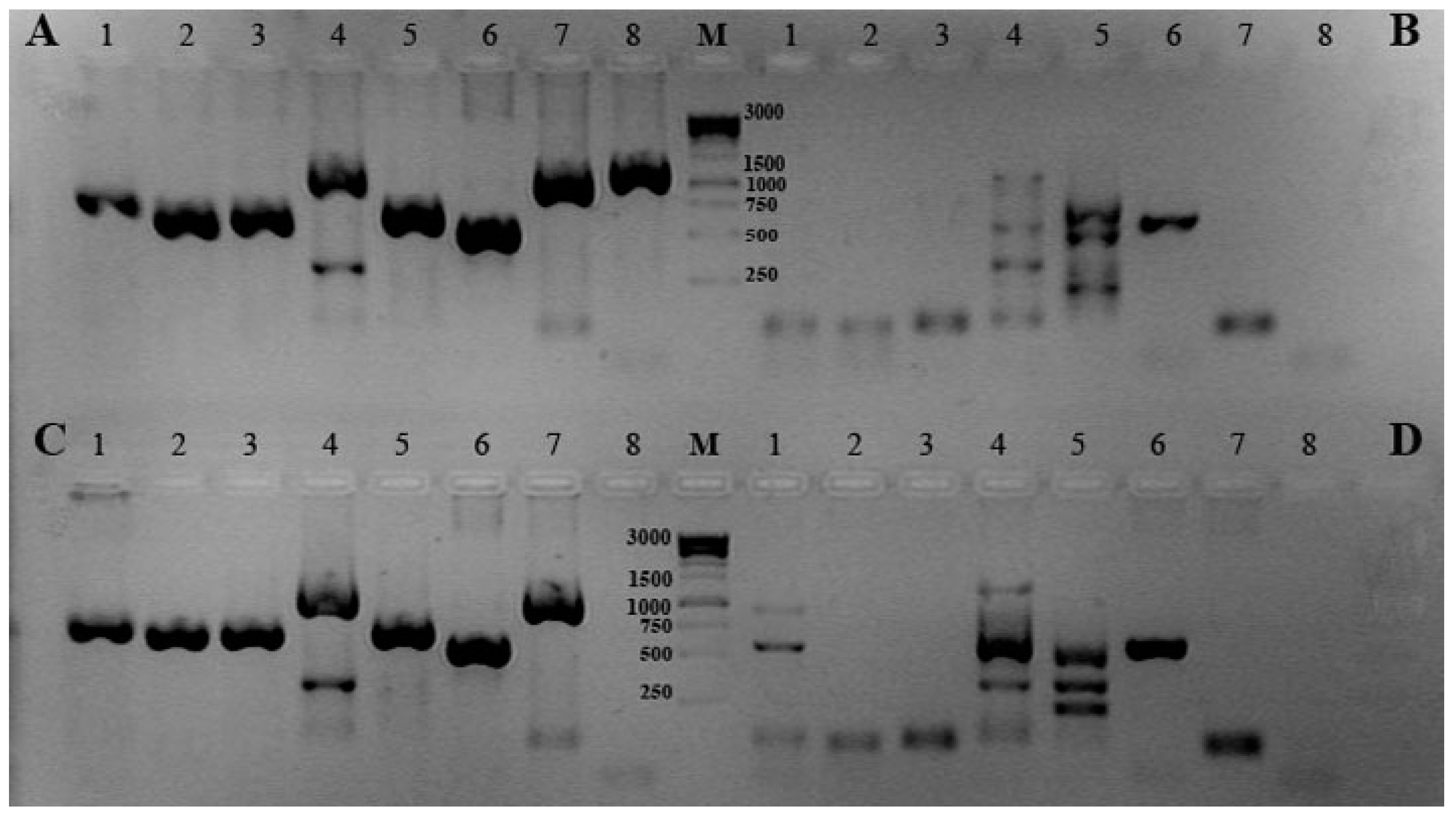
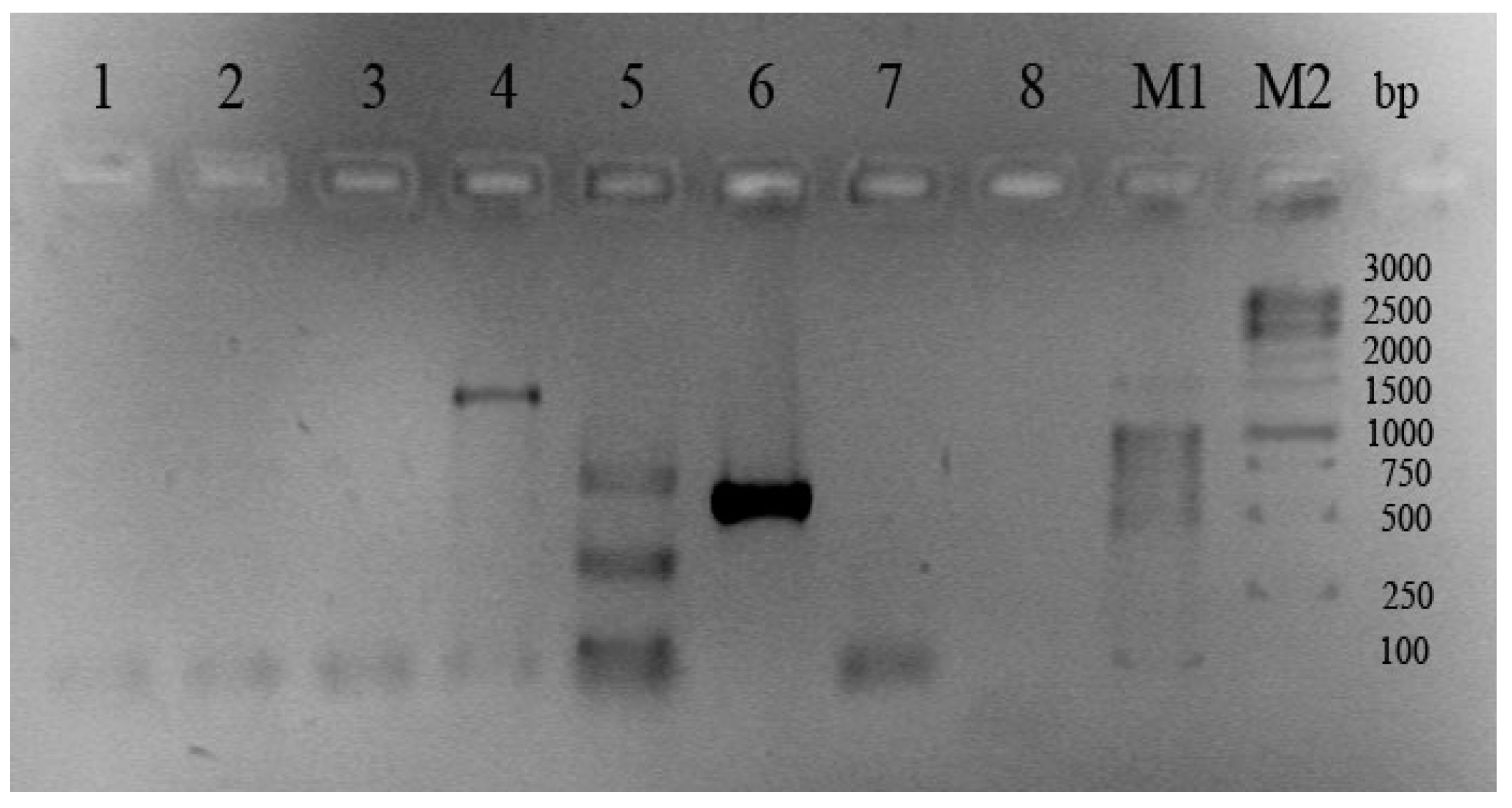
2. Results and Discussion
3. Materials and Methods
3.1. Isolation of the Strains Belonging to the Genus Cobetia
3.2. Isolation and Analysis of DNA from the Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Cobet, A.B.; Wirsen, C.J.; Jones, G.E. The effect of nickel on a marine bacterium, Arthrobacter marinus sp.nov. J. Gen. Microbiol. 1970, 62, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, L.; Baumann, P.; Mandel, M.; Allen, R.D. Taxonomy of aerobic marine eubacteria. J. Bacteriol. 1972, 110, 402–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, L.; Bowditch, R.D.; Baumann, P. Description of Deleya gen. nov. created to accommodate the marine species Alcaligenes aestus, A. pacificus, A. cupidus, A. venustus, and Pseudomonas marina. Int. J. Syst. Bacteriol. 1983, 33, 793–802. [Google Scholar] [CrossRef] [Green Version]
- Dobson, S.J.; Franzmann, P.D. Unification of the genera Deleya (Baumann et al. 1983), Halomonas (Vreeland et al. 1980), and Halovibrio (Fendrich 1988) and the species Paracoccus halodenitrificans (Robinson and Gibbons 1952) into a single genus, Halomonas, and placement of the genus Zymobacter in the family Halomonadaceae. Int. J. Syst. Bacteriol. 1996, 46, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Arahal, D.R.; Castillo, A.M.; Ludwig, W.; Schleifer, K.H.; Ventosa, A. Proposal of Cobetia marina gen. nov., comb. nov., within the family Halomonadaceae, to include the species Halomonas marina. Syst. Appl. Microbiol. 2002, 25, 207–211. [Google Scholar] [CrossRef]
- Kim, M.S.; Roh, S.W.; Bae, J.W. Cobetia crustatorum sp. nov., a novel slightly halophilic bacterium isolated from traditional fermented seafood in Korea. Int. J. Syst. Evol. Microbiol. 2010, 60, 620–626. [Google Scholar] [CrossRef] [Green Version]
- Romanenko, L.A.; Tanaka, N.; Svetashev, V.I.; Falsen, E. Description of Cobetia amphilecti sp. nov., Cobetia litoralis sp. nov. and Cobetia pacifica sp. nov., classification of Halomonas halodurans as a later heterotypic synonym of Cobetia marina and emended descriptions of the genus Cobetia and Cobetia marina. Int. J. Syst. Evol. Microbiol. 2013, 63, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Xu, K.; Han, X.; Mo, Z.; Mao, Y. Complete genome of Cobetia marina JCM 21022T and phylogenomic analysis of the family Halomonadaceae. J. Oceanol. Limnol. 2018, 36, 528–536. [Google Scholar] [CrossRef]
- Balabanova, L.A.; Golotin, V.A.; Kovalchuk, S.N.; Babii, A.V.; Shevchenko, L.S.; Son, O.M.; Kosovsky, G.Y.; Rasskazov, V.A. The Genome of the marine bacterium Cobetia marina KMM 296 isolated from the mussel Crenomytilus grayanus (Dunker, 1853). Russ. J. Mar. Biol. 2016, 42, 106–109. [Google Scholar] [CrossRef]
- Cheng, W.; Yan, X.; Xiao, J.; Chen, Y.; Chen, M.; Jin, J.; Bai, Y.; Wang, Q.; Liao, Z.; Chen, Q. Isolation, identification, and whole genome sequence analysis of the alginate-degrading bacterium Cobetia sp. cqz5-12. Sci. Rep. 2020, 2, 10920. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Cao, B.; Qiu, X.; Lin, J. Draft genome sequence of the crude oil-degrading and biosurfactant-producing strain Cobetia sp. QF-1. Genome Announc. 2018, 6, e01456-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balabanova, L.; Podvolotskaya, A.; Slepchenko, L.; Eliseikina, M.; Noskova, Y.; Nedashkovskaya, O.; Son, O.; Tekutyeva, L.; Rasskazov, V. Nucleolytic enzymes from the marine bacterium Cobetia amphilecti KMM 296 with antibiofilm activity and biopreservative effect on meat products. Food Control. 2017, 78, 270–278. [Google Scholar] [CrossRef]
- Moriya, H.; Takita, Y.; Matsumoto, A.; Yamahata, Y.; Nishimukai, M.; Miyazaki, M.; Shimoi, H.; Kawai, S.J.; Yamada, M. Cobetia sp. bacteria, which are capable of utilizing alginate or waste Laminaria sp. for poly(3-hydroxybutyrate) synthesis, isolated from a marine environment. Front. Bioeng. Biotechnol. 2020, 25, 974. [Google Scholar] [CrossRef] [PubMed]
- Mei, R.; Zhou, M.; Xu, L.; Zhang, Y.; Su, X. Characterization of a pH-tolerant strain Cobetia sp. SASS1 and its phenol degradation performance under salinity condition. Front. Microbiol. 2019, 4, 2034. [Google Scholar] [CrossRef] [Green Version]
- Coenye, T.; Vandamme, P. Intragenomic heterogeneity between multiple 16S ribosomal RNA operons in sequenced bacterial genomes. FEMS Microbiol. Lett. 2003, 228, 45–49. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Franco-Duarte, R.; Černáková, L.; Kadam, S.; Kaushik, K.S.; Salehi, B.; Bevilacqua, A.; Corbo, M.R.; Antolak, H.; Dybka-Stępień, K.; Leszczewicz, M.; et al. Advances in chemical and biological methods to identify microorganisms-from past to present. Microorganisms 2019, 7, 130. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Li, L.; Khan, M.A.; Zhu, F. Popular molecular markers in bacteria. Mol. Gen. Mikrobiol. Virusol. 2012, 3, 14–17. [Google Scholar] [CrossRef]
- Ashok Kumar, J.; Vinaya Kumar, K.; Avunje, S.; Akhil, V.; Ashok, S.; Kumar, S.; Sivamani, B.; Grover, M.; Rai, A.; Alavandi, S.V.; et al. Phylogenetic relationship among brackishwater Vibrio Species. Evol. Bioinform. Online. 2020, 16, 1176934320903288. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, E.; Christen, R.; Sawabe, T.; Alexeeva, Y.; Lysenko, A.; Chelomin, V.; Mikhailov, V. Presence of ecophysiologically diverse populations within Cobetia marina strains isolated from marine invertebrate, algae and the environments. Microbes Environ. 2005, 20, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Balabanova, L.; Nedashkovskaya, O.; Podvolotskaya, A.; Slepchenko, L.; Golotin, V.; Belik, A.; Shevchenko, L.; Son, O.; Rasskazov, V. Data supporting functional diversity of the marine bacterium Cobetia amphilecti KMM 296. Data Brief 2016, 8, 726–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 8, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plisova, E.Y.; Balabanova, L.A.; Ivanova, E.P.; Kozhemyako, V.B.; Mikhailov, V.V.; Agafonova, E.V.; Rasskazov, V.A. A highly active alkaline phosphatase from the marine bacterium Cobetia. Mar. Biotechnol. 2005, 7, 173–178. [Google Scholar] [CrossRef]
- Noskova, Y.; Likhatskaya, G.; Terentieva, N.; Son, O.; Tekutyeva, L.; Balabanova, L. A novel alkaline phosphatase/phosphodiesterase, CamPhoD, from marine bacterium Cobetia amphilecti KMM 296. Mar. Drugs. 2019, 17, 657. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Tu, Z.; Zhang, Y.; Li, J.; Feng, Y.; Yang, N.; Sang, X.; Chen, Q. Identification and characterization of DNA endonucleases in Plasmodium falciparum 3D7 clone. Malar. J. 2018, 17, 232. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, T.; Zheng, W.; Li, Z.H.; Ying, R.F.; Tang, Z.X.; Shi, L.E. Active sites and thermostability of a non-specific nuclease from Yersinia enterocolitica subsp. palearctica by site-directed mutagenesis. Biotechnol. Biotechnol. Equip. 2018, 32, 1306–1316. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, X.; Qin, J.; Cheng, S.; Yeo, W.-S.; He, L.; Ma, X.; Liu, X.; Li, M.; Bae, T. 2017 The ATP-dependent protease ClpP inhibits biofilm formation by regulating Agr and Cell wall hydrolase Sle1 in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2017, 7, 181. [Google Scholar] [CrossRef]
- Sutto-Ortiz, P.; Camacho-Ruiz, M.; Kirchmayr, M.R.; Camacho-Ruiz, R.M.; Mateos-Díaz, J.C.; Noiriel, A.; Carrière, F.; Abousalham, A.; Rodríguez, J.A. Screening of phospholipase A activity and its production by new actinomycete strains cultivated by solid-state fermentation. PeerJ 2017, 5, e3524. [Google Scholar] [CrossRef]
- Golotin, V.; Balabanova, L.; Likhatskaya, G.; Rasskazov, V. Recombinant production and characterization of a highly active alkaline phosphatase from marine bacterium Cobetia marina. Mar. Biotechnol. (NY) 2015, 17, 130–143. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]
- Vos, M. A species concept for bacteria based on adaptive divergence. Trends Microbiol. 2011, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rader, B.A.; Kremer, N.; Apicella, M.A.; Goldman, W.E.; McFall-Ngai, M.J. Modulation of symbiont lipid A signaling by host alkaline phosphatases in the squid-vibrio symbiosis. mBio. 2012, 1, e00093-e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, M.Ø.; Olderøy, M.; Zhang, Z.; Andreassen, J.-P.; Strandd, B.L.; Sikorski, P. Biocomposites prepared by alkaline phosphatase mediated mineralization of alginate microbeads. RSC Adv. 2012, 2, 1457–1465. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, E.P.; Mikhailo, V.V.; Plisova, E.J.; Balabanova, L.A.; Svetashev, V.V.; Vysockyi, M.V.; Stepanenko, V.I.; Rasskazov, V.A. Characterization of the marine bacterium Deleya marina producing highly active alkaline phosphatase and associated with the mussel Crenomytilus grayanus. Russ. J. Mar. Biol. 1994, 20, 340–545. [Google Scholar]



{kind=link}
{kind=link}
| Isolate | Isolation Source | Collection Number, KMM * | Results of Identification | 16S rRNA Genbank ID | |
|---|---|---|---|---|---|
| 16S rRNA, % Identity | PCR-Based Method | ||||
| Cobetia sp. 1AS1 | Coastal seawater, Vostok Bay, Sea of Japan | KMM 7516 | C. amphilecti, 100% | C. amphilecti | MW332480 |
| C. amphilecti KMM 296 | Coelomic liquid, mussel Crenomytilus grayanus | KMM 296 | C. amphilecti, 100% | C. amphilecti | NZ_JQJA01000078 |
| Cobetia sp. 29-18-1 | Sponge Esperiopsis digitata, Sea of Okhotsk, Is. Sakhalin, Piltun bay, 107 m. | KMM 7000 | C. amphilecti, 100%/C. litoralis, 100% | C. litoralis | MW332487 |
| Cobetia sp. 2AS | Sediments, Vostok Bay, Sea of Japan | KMM 7514 | C. amphilecti, 99.93%/C. litoralis 99.86% | C. litoralis | MW332483 |
| Cobetia sp. 2AS1 | Sediments, Vostok Bay, Sea of Japan | KMM 7005 | C. amphilecti, 100% | C. litoralis | MW332484 |
| Cobetia sp. 41-10Alg46 | The red algae Ahnfeltia tobuchiensis, collected near Is. Paramushir, Kuril Isles, Sea of Okhotsk | KMM 6284 | C. marina/C. pacifica, 100% | C. marina | MK587632 |
| Cobetia sp. 2S | Coastal seawater, Vostok Bay, Sea of Japan | KMM 7508 | C. marina/C. pacifica, 99.86% | C. pacifica | MW332481 |
| Cobetia sp. 3AS | Coastal seawater, Vostok Bay, Sea of Japan | KMM 7515 | C. marina/C. pacifica, 99.86% | C. pacifica | MW332482 |
| Cobetia sp. 11Alg1 | The red algae A. tobuchiensis (long-time cultivated), collected near Island Paramushir, Kuril Isles, Sea of Okhotsk | KMM 6816 | C. marina/C. pacifica, 100% | C. pacifica | MW332485 |
| Cobetia sp. 11Alg14 | The red algae A. tobuchiensis (long-time cultivated), collected near Island Paramushir, Kuril Isles, the Sea of Okhotsk | KMM 6818 | C. marina/C. pacifica, 100% | C. pacifica | MW332486 |
| Cobetia sp. 3AK | Coastal seawater, Vostok Bay, Sea of Japan | KMM 7505 | C. marina/C. pacifica, 100% | C. pacifica | MW332488 |
| Cobetia sp. 41-10Alg146 | The red alga A. tobuchiensis, collected near Is. Paramushir, Kuril Isles, Sea of Okhotsk | KMM 6731 | C. marina/C. pacifica, 100% | C. pacifica | KC247358 |
| Cobetia sp. 11Alg4 | The red algae A. tobuchiensis (long-time cultivated), collected near Is. Paramushir, Kuril Isles, Sea of Okhotsk | KMM 6817 | C. crustatorum, 100% | C. crustatorum | MW332489 |
| Name | Sequence | Molecular Marker * | Reference ** |
|---|---|---|---|
| 1CmNucF | 5′–TATACCATGGACGATATTCGCTCGGCCGGCCGCAA-3′ | DNA/RNA non-specific endonuclease precursor (1) and without leader peptide (2) | WP_043334786 |
| CmNucR | 5′–TATAGAGCTCTCAGTAACGTGATGGCGTACGACTG-3′ | ||
| 2CmNucF | 5′-TATACCATGGTATGGCAGGAGCGCGACTACCAGCA-3′ | ||
| CmNucR | 5′–TATAGAGCTCTCAGTAACGTGATGGCGTACGACTG -3′ | ||
| CmEEPf | 5′–TATACCATGGGACTCGACGAGACGGCACCTCCCCT -3′ | exonuclease/ endonuclease/phosphatase (EEP) | WP_084589364 |
| CmEEPr | 5′–TATAGAGCTCTTATGCTAGCCCGATCGCCTTGCGGCA-3′ | ||
| CmPhoDf | 5′–TATACCATGGAAGGACGGCGCCCGCGCATGCCCTC-3′ | alkaline phosphatase/phosphodiesterase PhoD | WP_043333989 |
| CmPhoDr | 5′–TATAGAGCTCTTAGACACTGGCGGCGGCGGGGGTC-3′ | ||
| CmPLA_f | 5′–TATACCATGGTACTCGATGAAAGCCTGGCCCAGCA-3′ | phospholipase A | WP_084589432 |
| CmPLA_r | 5′–TATAGAGCTCTTAGGTCTCTGGCGAGCCGGCGAAG-3′ | ||
| Tryp_F | 5′–TATACCATGGTACGTGAATTGCCCGACTTCACCCA-3′ | periplasmic serine peptidase Do/DeqQ | KGA03014 |
| Tryp_R | 5′–TATACTCGAGTCACTTGTCGCTGTCGGCACGCATG-3′ | ||
| CmClp_F | 5′-TATCCATGGTAAACGACTTCGACATCAAGAATGCT-3′ | ATP-dependent caseinolytic protease Clp | KGA03297 |
| CmClp_R | 5′-TATAGAGCTCTCACTCCACGTCGGGACGGCGTTCC-3′ | ||
| X-PhoN_F | 5′-TTAACCATGGCAGAGATCAAGAATGTCATTCTGAT-3′ | alkaline phosphatase PhoA | DQ435608 |
| CmAP_R | 5′-TTAAGAATTCCTTCGCTACCACTGTCTTCAGATACTGTCC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noskova, Y.; Seitkalieva, A.; Nedashkovskaya, O.; Shevchenko, L.; Tekutyeva, L.; Son, O.; Balabanova, L. Are the Closely Related Cobetia Strains of Different Species? Molecules 2021, 26, 690. https://doi.org/10.3390/molecules26030690
Noskova Y, Seitkalieva A, Nedashkovskaya O, Shevchenko L, Tekutyeva L, Son O, Balabanova L. Are the Closely Related Cobetia Strains of Different Species? Molecules. 2021; 26(3):690. https://doi.org/10.3390/molecules26030690
Chicago/Turabian StyleNoskova, Yulia, Aleksandra Seitkalieva, Olga Nedashkovskaya, Liudmila Shevchenko, Liudmila Tekutyeva, Oksana Son, and Larissa Balabanova. 2021. "Are the Closely Related Cobetia Strains of Different Species?" Molecules 26, no. 3: 690. https://doi.org/10.3390/molecules26030690
APA StyleNoskova, Y., Seitkalieva, A., Nedashkovskaya, O., Shevchenko, L., Tekutyeva, L., Son, O., & Balabanova, L. (2021). Are the Closely Related Cobetia Strains of Different Species? Molecules, 26(3), 690. https://doi.org/10.3390/molecules26030690