
Proteome Analysis of Aflibercept Intervention in Experimental Central Retinal Vein Occlusion

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
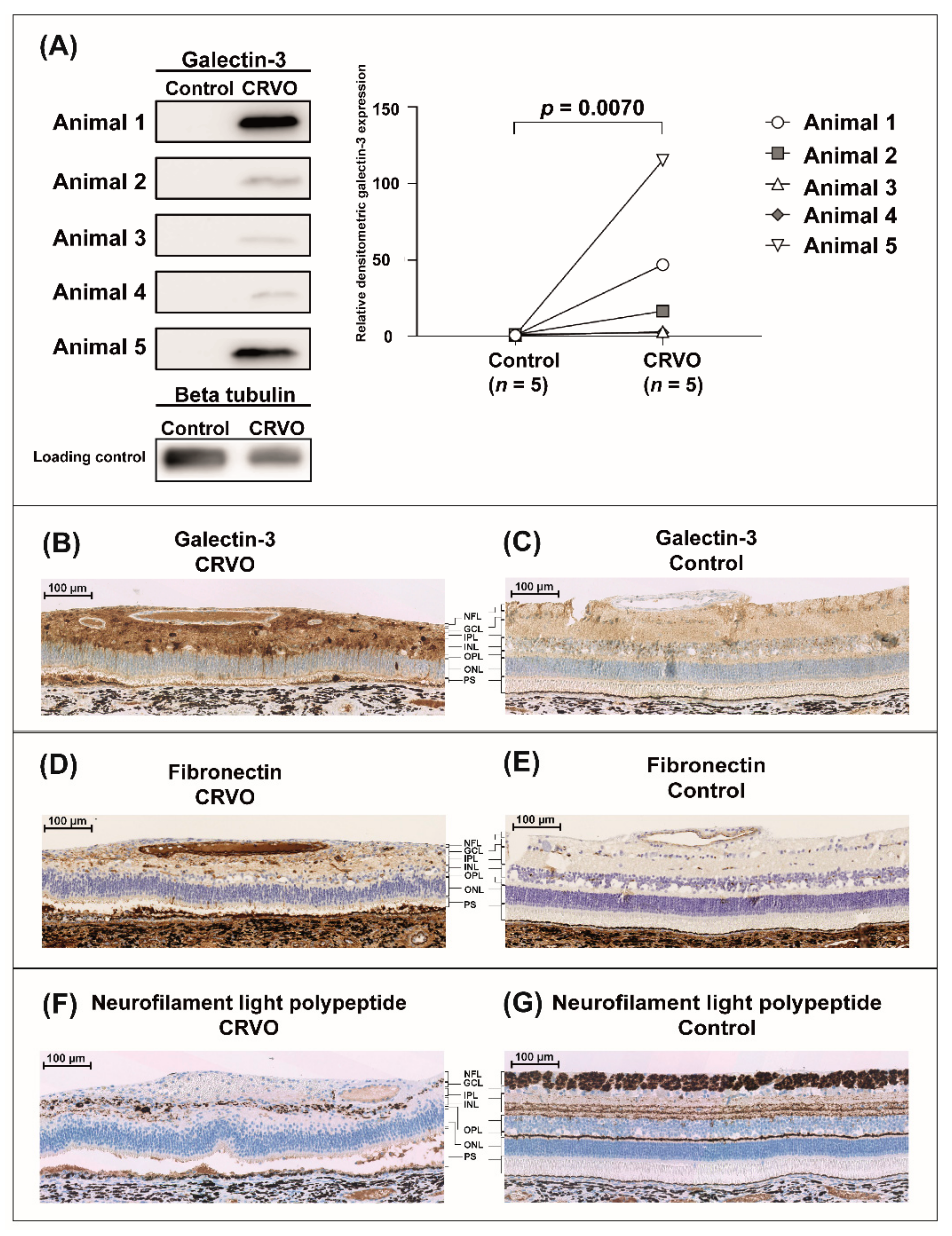
2.1. Evaluation of Experimental CRVO Model
2.2. Retinal Proteome Changes Following Aflibercept Intervention in Experimental CRVO
3. Discussion
3.1. Evaluation of Experimental CRVO Model
3.2. Aflibercept Intervention in CRVO
4. Materials and Methods
4.1. Animal Preparation
4.2. Experimental CRVO
4.3. Sample Preparation for Mass Spectrometry
4.4. Quantification with Tandem Mass Tag-Based Mass Spectrometry
4.5. Filtration of Proteins and Statistics
4.6. Immunohistochemistry
4.7. Western Blotting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Green, W.R.; Chan, C.C.; Hutchins, G.M.; Terry, J.M. Central retinal vein occlusion: A prospective histopathologic study of 29 eyes in 28 cases. Trans. Am. Ophthalmol. Soc. 1981, 79, 371–422. [Google Scholar] [CrossRef] [PubMed]
- Hayreh, S.S.; Podhajsky, P.A.; Zimmerman, M.B. Natural history of visual outcome in central retinal vein occlusion. Ophthalmology 2011, 118, 119–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noma, H.; Mimura, T.; Yasuda, K.; Shimura, M. Role of soluble vascular endothelial growth factor receptor signaling and other factors or cytokines in central retinal vein occlusion with macular edema. Investig. Ophthalmol. Vis. Sci. 2015, 56, 1122–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campochiaro, P.A.; Akhlaq, A. Sustained suppression of VEGF for treatment of retinal/choroidal vascular diseases. Prog. Retin. Eye Res. 2020, 83, 100921. [Google Scholar] [CrossRef] [PubMed]
- Petri, A.S.; Boysen, K.; Cehofski, L.J.; van Dijk, E.H.C.; Dysli, C.; Fuchs, J.; Mastropasqua, R.; Subhi, Y. Intravitreal Injections with Vascular Endothelial Growth Factor Inhibitors: A Practical Approach. Ophthalmol. Ther. 2020, 9, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holz, F.G.; Roider, J.; Ogura, Y.; Korobelnik, J.F.; Simader, C.; Groetzbach, G.; Vitti, R.; Berliner, A.J.; Hiemeyer, F.; Beckmann, K.; et al. VEGF Trap-Eye for macular oedema secondary to central retinal vein occlusion: 6-month results of the phase III GALILEO study. Br. J. Ophthalmol. 2013, 97, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Korobelnik, J.F.; Larsen, M.; Eter, N.; Bailey, C.; Wolf, S.; Schmelter, T.; Allmeier, H.; Chaudhary, V. Efficacy and Safety of Intravitreal Aflibercept Treat-and-Extend for Macular Edema in Central Retinal Vein Occlusion: The CENTERA Study. Am. J. Ophthalmol. 2021, 227, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Braithwaite, T.; Nanji, A.A.; Lindsley, K.; Greenberg, P.B. Anti-vascular endothelial growth factor for macular oedema secondary to central retinal vein occlusion. Cochrane Database Syst. Rev. 2014, 5, CD007325. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.W.; Le Couter, J.; Strauss, E.C.; Ferrara, N. Vascular endothelial growth factor a in intraocular vascular disease. Ophthalmology 2013, 120, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Cehofski, L.J.; Kruse, A.; Alsing, A.N.; Sejergaard, B.F.; Nielsen, J.E.; Pedersen, S.; Muttuvelu, D.V.; Kirkeby, S.; Honoré, B.; Vorum, H. Aflibercept Intervention in Experimental Branch Retinal Vein Occlusion Results in Upregulation of DnaJ Homolog Subfamily C Member 17. J. Ophthalmol. 2021, 2021, 6690260. [Google Scholar] [CrossRef] [PubMed]
- Cehofski, L.J.; Honore, B.; Vorum, H. A Review: Proteomics in Retinal Artery Occlusion, Retinal Vein Occlusion, Diabetic Retinopathy and Acquired Macular Disorders. Int. J. Mol. Sci. 2017, 18, 907. [Google Scholar] [CrossRef] [PubMed]
- Cehofski, L.J.; Mandal, N.; Honore, B.; Vorum, H. Analytical platforms in vitreoretinal proteomics. Bioanalysis 2014, 6, 3051–3066. [Google Scholar] [CrossRef] [PubMed]
- Cehofski, L.J.; Kojima, K.; Terao, N.; Kitazawa, K.; Thineshkumar, S.; Grauslund, J.; Vorum, H.; Honoré, B. Aqueous Fibronectin Correlates with Severity of Macular Edema and Visual Acuity in Patients with Branch Retinal Vein Occlusion: A Proteome Study. Investig. Ophthalmol. Vis. Sci. 2020, 61, 6. [Google Scholar] [CrossRef] [PubMed]
- Cehofski, L.J.; Kruse, A.; Kirkeby, S.; Alsing, A.N.; Ellegaard Nielsen, J.; Kojima, K.; Honore, B.; Vorum, H. IL-18 and S100A12 Are Upregulated in Experimental Central Retinal Vein Occlusion. Int. J. Mol. Sci. 2018, 19, 2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Bai, Y.; Huang, L.; Zhou, P.; Yu, W.; Zhao, M. Knockout of αA-crystallin inhibits ocular neovascularization. Investig. Ophthalmol. Vis. Sci. 2015, 56, 816–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cehofski, L.J.; Kruse, A.; Magnusdottir, S.O.; Alsing, A.N.; Nielsen, J.E.; Kirkeby, S.; Honore, B.; Vorum, H. Dexamethasone intravitreal implant downregulates PDGFR-alpha and upregulates caveolin-1 in experimental branch retinal vein occlusion. Exp. Eye Res. 2018, 171, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Cehofski, L.J.; Kruse, A.; Kjaergaard, B.; Stensballe, A.; Honore, B.; Vorum, H. Proteins involved in focal adhesion signaling pathways are differentially regulated in experimental branch retinal vein occlusion. Exp. Eye Res. 2015, 138, 87–95. [Google Scholar] [CrossRef]
- Cehofski, L.J.; Kruse, A.; Bogsted, M.; Magnusdottir, S.O.; Stensballe, A.; Honore, B.; Vorum, H. Retinal proteome changes following experimental branch retinal vein occlusion and intervention with ranibizumab. Exp. Eye Res. 2016, 152, 49–56. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein IDs | Protein Name | Gene Name | p-Value | Fold Change Aflibercept/NaCl |
|---|---|---|---|---|
| P17948 | Vascular endothelial growth factor receptor 1 * | FLT1 | p < 0.001 | 46.42 |
| P0DOX5;P01857 | Ig gamma-1 chain C region ** | IGHG1 | p < 0.001 | 18.90 |
| P35968 | Vascular endothelial growth factor receptor 2 *** | KDR | p < 0.001 | 5.07 |
| O43823 | A-kinase anchor protein 8 | AKAP8 | 0.027 | 1.19 |
| O60831 | PRA1 family protein 2 | PRAF2 | 0.039 | 0.91 |
| Q29092;P14625 | Endoplasmin | HSP90B1 | 0.041 | 0.91 |
| Q9NR30-2;Q9NR30 | Nucleolar RNA helicase 2 | DDX21 | 0.046 | 0.90 |
| O15258 | Protein RER1 | RER1 | 0.0091 | 0.90 |
| O60518 | Ran-binding protein 6 | RANBP6 | 0.030 | 0.90 |
| Q99615 | DnaJ homolog subfamily C member 7 | DNAJC7 | 0.0057 | 0.89 |
| Q92522 | Histone H1x | H1FX | 0.046 | 0.88 |
| Q9HCM4 | Band 4.1-like protein 5 | EPB41L5 | 0.033 | 0.88 |
| Q9HCS7 | Pre-mRNA-splicing factor SYF1 | XAB2 | 0.019 | 0.87 |
| Q53SF7 | Cordon-bleu protein-like 1 | COBLL1 | 0.036 | 0.87 |
| P86791;P86790 | Vacuolar fusion protein CCZ1 homolog; Vacuolar fusion protein CCZ1 homolog B | CCZ1;CCZ1B | 0.029 | 0.87 |
| Q9NP97 | Dynein light chain roadblock-type 1 | DYNLRB1 | 0.027 | 0.85 |
| P36955 | Pigment epithelium-derived factor (PEDF) | SERPINF1 | 0.020 | 0.84 |
| Q5VZL5 | Zinc finger MYM-type protein 4 | ZMYM4 | 0.030 | 0.83 |
| P83916 | Chromobox protein homolog 1 | CBX1 | 0.046 | 0.82 |
| P62272;P62269 | 40S ribosomal protein S18 | RPS18 | 0.030 | 0.76 |
| Q9Y2R9 | 28S ribosomal protein S7, mitochondrial | MRPS7 | 0.026 | 0.70 |
| Protein IDs | Protein Name | Gene Name | p-Value | Fold Change Aflibercept/NaCl |
|---|---|---|---|---|
| P17948 | Vascular endothelial growth factor receptor 1 * | FLT1 | p < 0.001 | 46.42 |
| P0DOX5; P01857 | Ig gamma-1 chain C region ** | IGHG1 | p < 0.001 | 18.9 |
| P35968 | Vascular endothelial growth factor receptor 2 *** | KDR | p < 0.001 | 5.07 |
| P13618 | ATP synthase-coupling factor 6, mitochondria | ATP5PF | 0.13 | 0.47 |
| Q95274; P62328 | Thymosin beta-4; Hematopoietic system regulatory peptide | TMSB4X | 0.16 | 0.43 |
| P02475 | Alpha-crystallin A chain | CRYAA | 0.07 | 0.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cehofski, L.J.; Kruse, A.; Alsing, A.N.; Sejergaard, B.F.; Nielsen, J.E.; Schlosser, A.; Sorensen, G.L.; Grauslund, J.; Honoré, B.; Vorum, H. Proteome Analysis of Aflibercept Intervention in Experimental Central Retinal Vein Occlusion. Molecules 2022, 27, 3360. https://doi.org/10.3390/molecules27113360
Cehofski LJ, Kruse A, Alsing AN, Sejergaard BF, Nielsen JE, Schlosser A, Sorensen GL, Grauslund J, Honoré B, Vorum H. Proteome Analysis of Aflibercept Intervention in Experimental Central Retinal Vein Occlusion. Molecules. 2022; 27(11):3360. https://doi.org/10.3390/molecules27113360
Chicago/Turabian StyleCehofski, Lasse Jørgensen, Anders Kruse, Alexander Nørgaard Alsing, Benn Falch Sejergaard, Jonas Ellegaard Nielsen, Anders Schlosser, Grith Lykke Sorensen, Jakob Grauslund, Bent Honoré, and Henrik Vorum. 2022. "Proteome Analysis of Aflibercept Intervention in Experimental Central Retinal Vein Occlusion" Molecules 27, no. 11: 3360. https://doi.org/10.3390/molecules27113360