
Dipeptidyl Peptidase 3 Activity as a Promising Biomarker of Bone Fragility in Postmenopausal Women

 , , , ,
, , , ,
Abstract
:1. Introduction
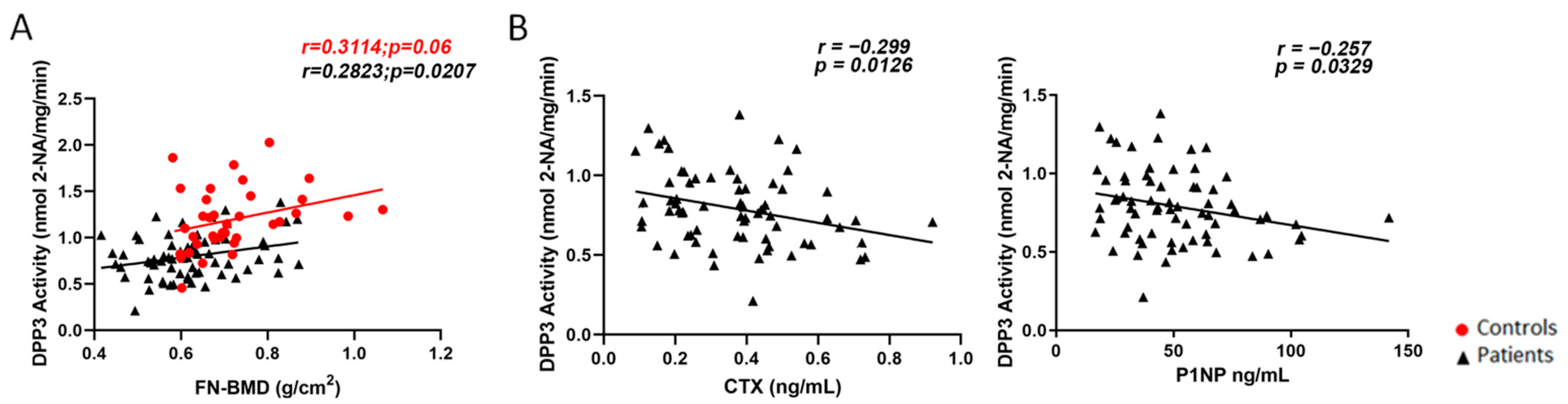
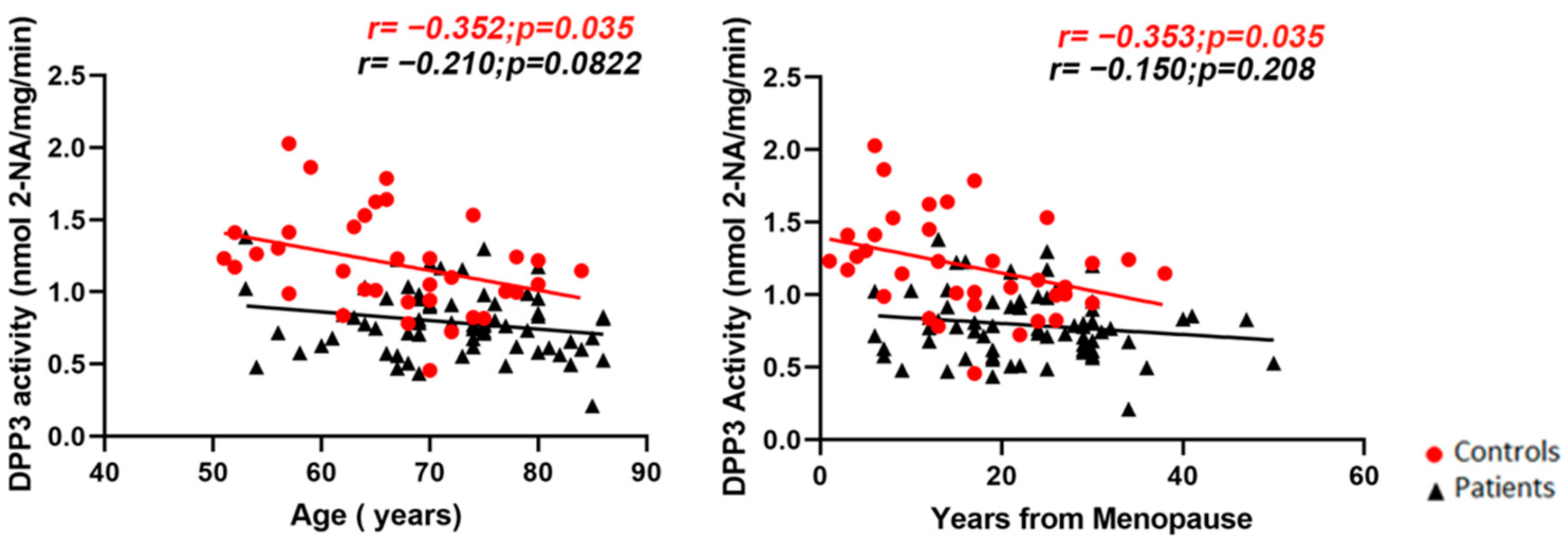
2. Results
2.1. Cross-Sectional Evaluation
2.2. Prospective Evaluation
3. Discussion
4. Materials and Methods
4.1. Study Population and Design
4.2. Biochemical Analysis
4.3. Dpp3 Activity Measurement
4.4. Dual-Energy X-ray Absorptiometry (DXA)
4.5. Vertebral Fracture Assessment
4.6. Ethics
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| Dpp3 | Dipeptidyl peptidase 3 |
| Keap1 | Kelch-like ECH associated protein-1 |
| Nrf2 | Nuclear factor erythroid 2-related factor-2 |
| CTX | C-terminal telopeptide |
| P1NP | N-terminal pro-peptide of type 1 procollagen |
| BMD | Bone mineral density |
| DXA | Dual-energy X-ray absorptiometry |
| SD | Standard deviation |
| PTH | Parathyroid hormone |
| GFR | Glomerular filtration rate |
| FN | Femoral neck |
| TH | Total hip |
| 2-NA | β-Naphthylamine |
| RANKL | Receptor activator of NFkB ligand |
| HO-1 | Heme oxygenase 1 |
| RAS | Renin–angiotensin system |
References
- Prajapati, S.C.; Chauhan, S.S. Dipeptidyl Peptidase III: A Multifaceted Oligopeptide N-End Cutter. FEBS J. 2011, 278, 3256–3276. [Google Scholar] [CrossRef]
- Kumar, P.; Reithofer, V.; Reisinger, M.; Wallner, S.; Pavkov-Keller, T.; Macheroux, P.; Gruber, K. Substrate Complexes of Human Dipeptidyl Peptidase III Reveal the Mechanism of Enzyme Inhibition. Sci. Rep. 2016, 6, 23787. [Google Scholar] [CrossRef] [Green Version]
- Hast, B.E.; Goldfarb, D.; Mulvaney, K.M.; Hast, M.A.; Siesser, P.F.; Yan, F.; Hayes, D.N.; Major, M.B. Proteomic Analysis of Ubiquitin Ligase KEAP1 Reveals Associated Proteins That Inhibit NRF2 Ubiquitination. Cancer Res. 2013, 73, 2199–2210. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Alcivar, A.L.; Ma, J.; Foo, T.K.; Zywea, S.; Mahdi, A.; Huo, Y.; Kensler, T.W.; Gatza, M.L.; Xia, B. NRF2 Induction Supporting Breast Cancer Cell Survival Is Enabled by Oxidative Stress-Induced Dpp3-KEAP1 Interaction. Cancer Res. 2017, 77, 2881–2892. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Yu, J.; Guo, L.; Ma, H. Dipeptidyl-Peptidase 3 Protects Oxygen-Glucose Deprivation/Reoxygenation-Injured Hippocampal Neurons by Suppressing Apoptosis, Oxidative Stress and Inflammation via Modulation of Keap1/Nrf2 Signaling. Int. Immunopharmacol. 2021, 96, 107595. [Google Scholar] [CrossRef]
- Tong, Y.; Huang, Y.; Zhang, Y.; Zeng, X.; Yan, M.; Xia, Z.; Lai, D. Dpp3/CDK1 Contributes to the Progression of Colorectal Cancer through Regulating Cell Proliferation, Cell Apoptosis, and Cell Migration. Cell Death Dis. 2021, 12, 529. [Google Scholar] [CrossRef]
- Menale, C.; Robinson, L.J.; Palagano, E.; Rigoni, R.; Erreni, M.; Almarza, A.J.; Strina, D.; Mantero, S.; Lizier, M.; Forlino, A.; et al. Absence of Dipeptidyl Peptidase 3 Increases Oxidative Stress and Causes Bone Loss. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2019, 34, 2133–2148. [Google Scholar] [CrossRef]
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporosis. Lancet Lond. Engl. 2019, 393, 364–376. [Google Scholar] [CrossRef]
- Choksi, P.; Jepsen, K.J.; Clines, G.A. The Challenges of Diagnosing Osteoporosis and the Limitations of Currently Available Tools. Clin. Diabetes Endocrinol. 2018, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Rehfeld, L.; Funk, E.; Jha, S.; Macheroux, P.; Melander, O.; Bergmann, A. Novel Methods for the Quantification of Dipeptidyl Peptidase 3 (Dpp3) Concentration and Activity in Human Blood Samples. J. Appl. Lab. Med. 2019, 3, 943–953. [Google Scholar] [CrossRef]
- Dépret, F.; Amzallag, J.; Pollina, A.; Fayolle-Pivot, L.; Coutrot, M.; Chaussard, M.; Santos, K.; Hartmann, O.; Jully, M.; Fratani, A.; et al. Circulating Dipeptidyl Peptidase-3 at Admission Is Associated with Circulatory Failure, Acute Kidney Injury and Death in Severely Ill Burn Patients. Crit. Care Lond. Engl. 2020, 24, 168. [Google Scholar] [CrossRef] [Green Version]
- Takagi, K.; Blet, A.; Levy, B.; Deniau, B.; Azibani, F.; Feliot, E.; Bergmann, A.; Santos, K.; Hartmann, O.; Gayat, E.; et al. Circulating Dipeptidyl Peptidase 3 and Alteration in Haemodynamics in Cardiogenic Shock: Results from the OptimaCC Trial. Eur. J. Heart Fail. 2020, 22, 279–286. [Google Scholar] [CrossRef]
- Ellis, S.; Nuenke, J.M. Dipeptidyl Arylamidase III of the Pituitary. Purification and Characterization. J. Biol. Chem. 1967, 242, 4623–4629. [Google Scholar] [CrossRef]
- Malovan, G.; Hierzberger, B.; Suraci, S.; Schaefer, M.; Santos, K.; Jha, S.; Macheroux, P. The Emerging Role of Dipeptidyl Peptidase 3 in Pathophysiology. FEBS J. 2022. [CrossRef]
- Black, D.M.; Cauley, J.A.; Wagman, R.; Ensrud, K.; Fink, H.A.; Hillier, T.A.; Lui, L.-Y.; Cummings, S.R.; Schousboe, J.T.; Napoli, N. The Ability of a Single BMD and Fracture History Assessment to Predict Fracture Over 25 Years in Postmenopausal Women: The Study of Osteoporotic Fractures. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2018, 33, 389–395. [Google Scholar] [CrossRef]
- Singh, K.B.; Rai, R.; Khanka, S.; Singh, D. Discontinuation of PTH Therapy Amplifies Bone Loss by Increasing Oxidative Stress: An Event Ameliorated by Sequential IL-17 Neutralizing Antibody Therapy. Biomed. Pharmacother. 2022, 145, 112390. [Google Scholar] [CrossRef]
- Reis, J.; Ramos, A. In Sickness and in Health: The Oxygen Reactive Species and the Bone. Front. Bioeng. Biotechnol. 2021, 9, 745911. [Google Scholar] [CrossRef]
- Mačak Šafranko, Ž.; Sobočanec, S.; Šarić, A.; Jajčanin-Jozić, N.; Krsnik, Ž.; Aralica, G.; Balog, T.; Abramić, M. The Effect of 17β-Estradiol on the Expression of Dipeptidyl Peptidase III and Heme Oxygenase 1 in Liver of CBA/H Mice. J. Endocrinol. Investig. 2015, 38, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Voiosu, A.M.; Wiese, S.; Goetze, J.P.; Hartmann, O.; Voiosu, T.; Santos, K.; Møller, S. Dipeptidyl Peptidase-3 Is Associated with Severity of Liver Disease and Circulatory Complications in Patients with Cirrhosis. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2022, 27, 196–204. [Google Scholar] [CrossRef]
- Van Lier, D.; Beunders, R.; Kox, M.; Pickkers, P. The Value of Bioactive Adrenomedullin and Dipeptidyl Peptidase 3 to Predict Short-Term Unfavourable Outcomes after Cardiac Surgery: A Prospective Cohort Study. Eur. J. Anaesthesiol. 2022, 39, 342–351. [Google Scholar] [CrossRef]
- Deniau, B.; Picod, A.; Van Lier, D.; Vaittinada Ayar, P.; Santos, K.; Hartmann, O.; Gayat, E.; Mebazaa, A.; Blet, A.; Azibani, F. High Plasma Dipeptidyl Peptidase 3 Levels Are Associated with Mortality and Organ Failure in Shock: Results from the International, Prospective and Observational FROG-ICU Cohort. Br. J. Anaesth. 2022, 128, e54–e57. [Google Scholar] [CrossRef]
- Frigyesi, A.; Lengquist, M.; Spångfors, M.; Annborn, M.; Cronberg, T.; Nielsen, N.; Levin, H.; Friberg, H. Circulating Dipeptidyl Peptidase 3 on Intensive Care Unit Admission Is a Predictor of Organ Dysfunction and Mortality. J. Intensive Care 2021, 9, 52. [Google Scholar] [CrossRef]
- Thorgeirsdóttir, B.; Levin, H.; Spångfors, M.; Annborn, M.; Cronberg, T.; Nielsen, N.; Lybeck, A.; Friberg, H.; Frigyesi, A. Plasma Proenkephalin A 119-159 and Dipeptidyl Peptidase 3 on Admission after Cardiac Arrest Help Predict Long-Term Neurological Outcome. Resuscitation 2021, 163, 108–115. [Google Scholar] [CrossRef]
- Boorsma, E.M.; Ter Maaten, J.M.; Damman, K.; van Veldhuisen, D.J.; Dickstein, K.; Anker, S.D.; Filippatos, G.; Lang, C.C.; Metra, M.; Santos, K.; et al. Dipeptidyl Peptidase 3, a Marker of the Antagonist Pathway of the Renin-Angiotensin-Aldosterone System in Patients with Heart Failure. Eur. J. Heart Fail. 2021, 23, 947–953. [Google Scholar] [CrossRef]
- Blet, A.; Deniau, B.; Santos, K.; van Lier, D.P.T.; Azibani, F.; Wittebole, X.; Chousterman, B.G.; Gayat, E.; Hartmann, O.; Struck, J.; et al. Monitoring Circulating Dipeptidyl Peptidase 3 (Dpp3) Predicts Improvement of Organ Failure and Survival in Sepsis: A Prospective Observational Multinational Study. Crit. Care Lond. Engl. 2021, 25, 61. [Google Scholar] [CrossRef]
- Han, J.; Yang, K.; An, J.; Jiang, N.; Fu, S.; Tang, X. The Role of NRF2 in Bone Metabolism-Friend or Foe? Front. Endocrinol. 2022, 13, 813057. [Google Scholar] [CrossRef]
- Kanis, J.A.; Kanis, J.A.; WHO Study Group. Assessment of Fracture Risk and Its Application to Screening for Postmenopausal Osteoporosis: Synopsis of a WHO Report. Osteoporos. Int. J. Establ. Result Coop. Eur. Found. Osteoporos. Natl. Osteoporos. Found. USA 1994, 4, 368–381. [Google Scholar] [CrossRef]
- Lewiecki, E.M.; Baim, S.; Binkley, N.; Bilezikian, J.P.; Kendler, D.L.; Hans, D.B.; Silverman, S.; International Society for Clinical Densitometry. Report of the International Society for Clinical Densitometry 2007 Adult Position Development Conference and Official Positions. South. Med. J. 2008, 101, 735–739. [Google Scholar] [CrossRef]
- Genant, H.K.; Wu, C.Y.; van Kuijk, C.; Nevitt, M.C. Vertebral Fracture Assessment Using a Semiquantitative Technique. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 1993, 8, 1137–1148. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients (n = 69) | Controls (n = 36) | p Value | |
|---|---|---|---|
| Age, years | 71.93 ± 8.21 | 66.64 ± 8.73 | 0.003 |
| Age at menopause, years | 49.01 ± 5.16 | 50.22 ± 2.95 | 0.131 |
| Calcium, mg/dL (8.4–10.2) | 9.36 ± 0.43 | 9.45 ± 0.37 | 0.256 |
| Phosphate, mg/dL (2.3–4.7) | 3.63 ± 0.47 | 3.81 ± 0.41 | 0.057 |
| 25-OH Vitamin D, ng/mL | 30.74 ± 13.25 | 29.91 ± 15.14 | 0.773 |
| PTH, pg/mL (13–85) | 52.81 ± 24.42 | 49.06 ± 15.32 | 0.338 |
| GFR, mL/min/m2 | 77.78 ± 21.42 | 88.51 ± 18.82 | 0.013 |
| CTX, ng/mL | 0.371 ± 0.18 | n.a. | n.a. |
| P1NP, ng/mL | 50.51 ± 24.57 | n.a. | n.a. |
| L1–L4 BMD, g/cm2 | 0.800 ± 0.158 | 0.939 ± 0.131 | 0.001 |
| FN-BMD, g/cm2 | 0.624 ± 0.108 | 0.718 ± 0.112 | 0.001 |
| TH-BMD, g/cm2 | 0.697 ± 0.126 | 0.874 ± 0.078 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menale, C.; Tabacco, G.; Naciu, A.M.; Schiavone, M.L.; Cannata, F.; Morenghi, E.; Sobacchi, C.; Palermo, A. Dipeptidyl Peptidase 3 Activity as a Promising Biomarker of Bone Fragility in Postmenopausal Women. Molecules 2022, 27, 3929. https://doi.org/10.3390/molecules27123929
Menale C, Tabacco G, Naciu AM, Schiavone ML, Cannata F, Morenghi E, Sobacchi C, Palermo A. Dipeptidyl Peptidase 3 Activity as a Promising Biomarker of Bone Fragility in Postmenopausal Women. Molecules. 2022; 27(12):3929. https://doi.org/10.3390/molecules27123929
Chicago/Turabian StyleMenale, Ciro, Gaia Tabacco, Anda Mihaela Naciu, Maria Lucia Schiavone, Francesca Cannata, Emanuela Morenghi, Cristina Sobacchi, and Andrea Palermo. 2022. "Dipeptidyl Peptidase 3 Activity as a Promising Biomarker of Bone Fragility in Postmenopausal Women" Molecules 27, no. 12: 3929. https://doi.org/10.3390/molecules27123929
APA StyleMenale, C., Tabacco, G., Naciu, A. M., Schiavone, M. L., Cannata, F., Morenghi, E., Sobacchi, C., & Palermo, A. (2022). Dipeptidyl Peptidase 3 Activity as a Promising Biomarker of Bone Fragility in Postmenopausal Women. Molecules, 27(12), 3929. https://doi.org/10.3390/molecules27123929