Abstract
Cancer is one of the world’s most burdensome diseases, with increasing prevalence and a high mortality rate threat. Tumor recurrence and metastasis due to treatment resistance are two of the primary reasons that cancers have been so difficult to treat. The epithelial–mesenchymal transition (EMT) is essential for tumor drug resistance. EMT causes tumor cells to produce mesenchymal stem cells and quickly adapt to various injuries, showing a treatment-resistant phenotype. In addition, multiple signaling pathways and regulatory mechanisms are involved in the EMT, resulting in resistance to treatment and hard eradication of the tumors. The purpose of this study is to review the link between EMT, therapeutic resistance, and the molecular process, and to offer a theoretical framework for EMT-based tumor-sensitization therapy.
1. Introduction
The epithelial–mesenchymal transition (EMT) is a cellular process that often occurs during normal or pathological processes. It is essential in embryonic development and wound healing and is crucial for tumor growth [1]. Many factors trigger EMT in tumor cells, such as tumor microenvironment [2,3] and inflammatory cytokines [4]. Indeed, EMT is associated with increased cancer stem cells, inadequate anti-tumor immunity, and resistance to oncology therapies. Consequently, EMT is an essential component of the invasion-metastasis cascade. Despite the diversity of cancer types, the EMT process in patients with low-grade gliomas might have significant prognostic consequences [5].
When tumor cells undergo EMT, they lose their epithelioid characteristics and exhibit poor intercellular adhesion and polarity, resulting in enhanced tumor cell motility and invasion [6]. EMT is also associated with abnormal molecular expression patterns, including decreased epithelioid marker expression, increased mesenchymal marker expression, and EMT-related transcription factors. Given that epigenetic processes such as DNA methylation and chromatin changes may impact EMT, a complex but reversible process, they are required to maintain its regulation. Loss of the cell cycle regulator p21 has been associated with increased EMT characteristics and a rise in ZEB1, the master EMT transcription factor [7,8,9].
Tumor cells adapt to cytotoxicity and develop therapeutic resistance due to EMT-induced anti-apoptotic abilities, improved DNA damage repair, and a modified drug metabolism pathway [10]. At present, distinct EMT processes have been seen in tumor tissues and cells that are resistant to chemotherapy or radiation, suggesting that EMT is intrinsically linked to tumor treatment resistance [11,12,13]. In this review, we addressed the characteristics of EMT; further discussed tumor therapeutic resistance, discussed EMT-mediated therapeutic resistance and its molecular mechanism, and appraised the level of research on EMT and tumor therapeutic resistance.
2. Overview
Epithelial cells undergo a shift to mesenchymal cells during EMT. Their gel-cultured lens epithelial cells lose their polarity, extend their pseudopodia, and acquire mesenchymal features, in a process known as the epithelial–mesenchymal transition. EMT often develops due to physiological or pathological processes occurring inside the human body. According to the biological setting in which EMT occurs, it may be categorized as follows:
Type 1 EMT is a term that refers to an EMT that occurs during embryogenesis and organ development [14].
Type 2 EMT is defined as an EMT that responds to wound healing, tissue regeneration, or organ fibrosis [15,16].
Type 3 EMT refers to the conversion of epithelioid cells to mesenchymal cells that occurs during the development of malignant tumors and is associated with treatment resistance and distant metastasis [17,18]. This review discusses the phenotypic and molecular processes underlying the tumor treatment resistance generated by type 3 EMT.
3. Therapeutic Resistance of Tumors
Surgery, chemotherapy, radiation, and targeted molecular therapy are the principal therapies for solid tumors. These strategies have been shown to successfully remove, damage, and induce cell death, hence resulting in disease remission. A combination of one or more treatments may be used depending on the type of tumor and tumor stage. Patients with advanced cancers typically miss out on major surgery due to the extra chemotherapy, radiation therapy, and targeted therapy treatments required. While some degree of control is achievable, cancer chemotherapeutic drug resistance, radiation resistance, and targeted treatment resistance contribute to the failure to produce a radical curative effect, posing a substantial barrier to cancer eradication. Tumor treatment resistance is a complex process that is impacted by various factors, including the tumor microenvironment, non-coding RNA, and EMT [19,20,21,22,23].
Due to the heterogeneity of tumors, treatment-resistant mutations are filtered out throughout therapy, or latent, initially drug-resistant tumor stem cell subsets are awakened, resulting in treatment failure and tumor recurrence or metastasis. Additionally, tumors in the same area are varied in terms of cell type, clinical stage, and degree of differentiation. Furthermore, homogeneous tumors demonstrate significant molecular variety, including differences in gene expression patterns, network regulation, and mutation profiles [24,25]. The heterogeneity of patients with the same kind of tumor is reflected in the inconsistency of the changed gene spectrum and biological features across cells, reflecting malignant tumors’ enormous complexity and diversity during their lifetime. More importantly, tumor heterogeneity may contribute to tumor cell resistance to treatment by modifying epigenetic factors such as messenger RNA, the transcriptome, and the proteome. Simultaneously, tumor stem cells are related to resistance to cancer treatment [26,27,28]. Most cancer treatments, including anticancer drugs, are modulated by innate processes that function as a defense against environmental toxins. Defense mechanisms include drug efflux mediated by ATP binding box (ABC) transporters. The ABC transporters is the most common transmembrane transporter for drug efflux. The human genome consists of seven subfamilies of ABC genes (ABCA-ABCG). ABCB1, ABCC1, and ABCG2 are essential for developing cancer chemotherapy resistance (MDR) [29,30]. ABCB 1 was highly expressed on the surface of some tumor cells. In this case, the cytoplasm is able to pump the chemotherapeutic substance of the intracellular ABCB 1 substrate out of the cytoplasm. Overexpression of ABCB 1 on the cytoplasm surface further amplifies this response. This is one of the main mechanisms of reduced intracellular drug accumulation and the generation of multidrug resistance in tumors [31]. ABCG2, named Breast cancer resistance protein, is the main drug efflux transporter that leads to resistance in breast cancer. ABCG2 is thought to be a marker of cancer stem cells (CSCs) in some cancers and is to blame for the side-population effect. It can move medicines with either a positive or a negative electrical charge, starting with chemotherapeutic agents. ABCG2 substrate chemotherapeutic substances in tumor cells can be transferred to the outside of tumor cells, which reduces the sensitivity of tumor cells to chemotherapy drugs. This leads to the emergence of multidrug resistance [32,33]. For instance, the expression of the ABC transporters was similarly increased after TGF-β or Twist in breast cancer cells, and zeb1 exhibited the reverse-transition EMT and its resistance to the virobistar [34,35]. Similarly, it was reported that the transcription factors that govern EMT might be utilized to regulate the high expression of the ABC transporter protein by regulating the transmission-drug pump gene, therefore decreasing the drug concentration in the cell and making the cells susceptible to drug treatment. The EMT transcription factor may also be modulated by interfering with the apoptotic pathway to render tumor cells drug-resistant [36,37].
Recent advances in the tumor microenvironment study have shown that the tumor microenvironment is intricately linked to tumor treatment resistance. Indeed, the tumor microenvironment is composed of cellular and noncellular elements, including tumor cells, fibroblasts, endothelial cells, and immune cells [38,39,40]. The change from fibroblasts into tumor-associated fibroblasts in the tumor microenvironment may result in tumor resistance to chemotherapy. The infiltration of regulatory T cells into the tumor microenvironment may result in immunological resistance to targeted anti-VEGF treatment. When protected by the tumor microenvironment, non-tumor stromal cells and cell stroma provide an optimal habitat for surviving tumor cells and favorable conditions for tumor recurrence and metastasis [41,42].
Even though 95% of the genes in the human genome do not code for proteins, they do transcribe non-coding RNAs. Non-coding RNAs may be classified as housekeeping or regulatory RNAs based on their expression and functional properties. The former is essential for cell life activities and has a somewhat stable composition, enabling component expression. The latter has a high degree of selectivity, is often transiently expressed, and works as a transcriptional and translational regulator, affecting tumor treatment resistance. Among them, the association between long non-coding RNAs (lncRNAs), short non-coding RNAs (miRNAs), and circular non-coding RNAs (circRNAs) and tumor treatment resistance is gaining in attention. LncRNAs contribute to DNA repair and cell-cycle progression, drug metabolism and efflux, cell death, and EMT through encoding small-molecule polypeptides, contributing to tumor treatment resistance [43,44]. By augmenting the target gene’s mRNA with 6–7 bases and decreasing the target gene’s protein production level, miRNAs may confer resistance to malignancy therapy. Oncogenic miRNAs, in general, may reduce tumor suppressor gene expression, interfere with drug efferent pump molecules and cell cycle and apoptotic regulatory molecules, and ultimately result in tumor treatment resistance [45]. CircRNAs may function as an miRNA biomarker and therapeutic target for prognosis in malignancies via endogenous competition for miRNA and target mRNA binding sites or directly interact with proteins [46].
4. EMT-Induced Tumor Therapy Resistance
EMT was previously only associated with cancer cell invasion and metastasis. Recent research has shown that tumor cells undergoing EMT also have enhanced anti-apoptotic activity, which significantly contributes to tumor treatment resistance [47,48,49]. In addition, tumor cells that undergo EMT may acquire resistance to apoptosis, which directly influences the efficacy of radiation, chemotherapy, targeted therapy, and immunotherapy. Reversing EMT or eradicating tumor cells with EMT abnormalities has been proposed as a potential tumor therapy strategy [50].
4.1. EMT-Mediated Tumor Chemotherapy Resistance
Chemotherapy is a regularly utilized treatment approach for malignant tumors, although tumor cell resistance to chemotherapy drugs usually results in chemotherapy failure. Chemotherapy resistance mechanisms in tumor cells have previously been linked to cell cycle and phase specificity, drug uptake and efflux mechanisms, intracellular target substance concentrations, structural modifications, and enhanced DNA damage repair [51,52,53]. EMT has garnered significant attention as a potential mechanism of chemotherapeutic resistance in recent years due to its ability to turn tumor cells into tumor stem cells and confer resistance on tumor cells with resistance chemotherapy [54,55]. Without therapy, tumors are composed of clusters of chemically resistant cells that are dormant, dry, and well-coordinated in their EMT. These cells become more numerous during drug treatments and ultimately acquire resistance to chemotherapy. Similarly, EMT-mediated tumor phenotypic plasticity greatly influences resistance to chemotherapy in cancers [17].
Additionally, incomplete EMT or mesenchymal cells raised cancer stem cell markers and lowered the sensitivity to FAK inhibitors in MDA-MB-468 tumors treated with paclitaxel-nilotinib. Similar tumor stem cell markers were elevated in patients with metastatic prostate cancer treated with the PARP inhibitor tarazoparil [56,57]. Thus, the phenotypic plasticity conferred to tumor cells by EMT may result in tumor cells quickly responding to cytotoxicity or targeted molecular therapy, resulting in acquired drug resistance. By inhibiting FOXM1-mediated EMT and DNA repair, the expression of miR-671-5p dynamically decreases during the oncogenic transition of breast cancer. Therefore, miR-671-5p may be a promising biomarker for early breast cancer detection and a therapeutic target for breast cancer treatment [58].
Cisplatin is commonly used as a chemotherapeutic agent for oral squamous cell carcinoma. It was reported that EMT plays a significant role in the development of resistance to cisplatin therapy in oral squamous cell carcinoma. Mir-155, an exocrine compound, induced an EMT-mediated drug resistance phenotype in cisplatin-sensitive oral squamous cell carcinoma cells [59]. Through the induction of EMT, aberrant LncRNA expression may result in chemotherapeutic resistance. The lncRNA HOXA-AS3 induces EMT in NSCLC cells by targeting HOXA3 and increasing Twist expression [60]. Similarly, cisplatin treatment for gastric cancer may activate the estrogen receptor GPR30, leading to gastric cancer cells developing resistance to cisplatin through EMT [61,62]. In addition, EMT is inextricably connected to chemotherapy resistance in colorectal cancer. Snail, a molecule involved in EMT regulation, was significantly elevated in colorectal cancer tissues. The direct control of ABCB1 increased tumor cell resistance to 5-FU, indicating a probable relationship between EMT and treatment resistance [63] (Figure 1).
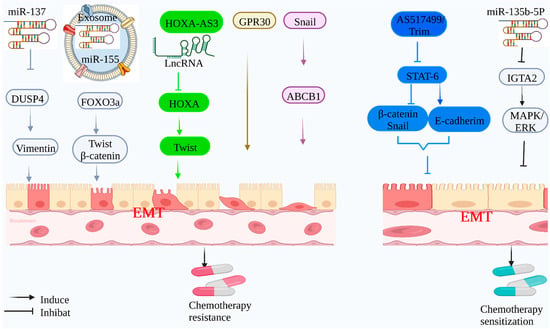
Figure 1.
Mechanism of EMT-mediated chemotherapy resistance: Tumor-associated genes can induce EMT to mediate chemotherapy resistance of tumor cells. Mir-137 can inhibit the expression of target protein DUSP4 and induce EMT, which activates vimentin; exosomal Mir-155 can target the expression of FOXO3a and up-regulate the expression of β-catenin and Twist; lncRNA HOXA-AS3 can inhibit the expression of HOXA3 and up-regulate the expression of Twist, as well as participating in mediating adriamycin and cisplatin drug resistance; tumor-associated protein GPR30 can induce EMT and mediate cisplatin resistance. The tumor-related protein Snail regulates ABCB1 expression, induces EMT, and mediates 5-FU resistance. At the same time, 5-FU combined with AS1517499 and Trim inhibited the phosphorylation of STAT-6, up-regulated the expression of E-cadherin, and down-regulated the expression of Snail and β-catenin, delaying the progression of EMT, and enhanced the sensitivity of 5-FU to EMT-mediated chemotherapy resistance. Mir-135b-5p can target the expression of IGTA2, down-regulate the MAPK/ERK signaling pathway, delay the progression of EMT, and enhance the sensitivity of 5-FU.
Given the crucial role of EMT in tumor chemotherapy resistance, it is desirable to target EMT and reverse it to MET to improve chemotherapy efficacy. 5-FU is the main treatment for colorectal cancer, and its combination with other adjuvant therapies (AS1517499 and Trim) has been demonstrated to enhance chemotherapy efficacy. When combined with 5-FU, this adjuvant treatment lowered STAT-6 phosphorylation, raised epithelial marker E-cadherin expression, decreased mesenchymal marker β-catenin and Snail1 expression, induced apoptosis, and enhanced 5-FU sensitivity [64]. The mir-135B-5p/ITGA2 signaling axis has been demonstrated to reduce the degree of EMT in gastric cancer chemotherapy trials by inhibiting the MAPK/ERK pathway, restoring gastric cancer cells’ chemotherapeutic sensitivity, and boosting mortality [65].
4.2. Tumor Radiation Resistance Mediated by EMT
Tumor radiotherapy is a strategy for locally treating tumor lesions with radiation, mainly killing or weakening tumor cells by inducing apoptosis and mitotic instability. Radiation resistance in tumors is a sophisticated process involving several genes and pathways that are tightly tied to cell cycle arrest, apoptosis resistance, tumor microenvironment changes, autophagy, and EMT, among other factors [66,67,68]. EMT-induced local recurrences and metastasis are critical causes of radiation failure. Numerous studies have shown a relationship between ionizing radiation-induced EMT in tumor cells, drug resistance in tumor cells, and the formation of tumor stem cells, although the process is not entirely known at the time [69,70].
EMT is intrinsically connected to tumor radiation resistance, and as EMT proceeds, tumor radiation sensitivity gradually decreases. TRIP4 overexpression in cervical cancer cells and tumor tissues may promote EMT and activate the PI3K/Akt and MAPK/ERK signaling pathways, enhancing radiation resistance in cervical cancer [71]. Radiation resistance is a frequent cause of the recurrence of glioblastoma. CXCL1 overexpression induces the mesenchymal transition and imparts radiation resistance to glioblastoma cells by stimulating the NF-B signaling pathway in glioblastoma cells [72].
Similarly, it was reported that EVI1, which is overexpressed in Nasopharyngeal carcinoma, could bind to Snail and HDAC1 to suppress E-cadherin synthesis and thus enhance the EMT of nasopharyngeal cancer cells. Meanwhile, EVI1 may directly interact with the promoter of β-catenin, boosting the stem cell properties of nasopharyngeal carcinoma (NPC) cells. EVI1 plays a role in nasopharyngeal cancer patients’ radiation resistance [73]. Individuals with recurrent nasopharyngeal cancer had more distant metastases and were more likely to develop radiation resistance after a second treatment than those with first diagnosed nasopharyngeal cancer. CCL2 expression was significantly increased in HONe1-IR cells and recurrent NPC tumors and contributed to radiation resistance through EMT. Additionally, inhibiting the mitochondrial pyruvate carrier (MPC) has been shown to promote EMT and confer radiation resistance in pancreatic and colorectal cancer cells [74].
Radiation resistance is inextricably linked to EMT and DNA damage repair in advanced colorectal cancer, and mir-130a may play a role in the radiation resistance mechanism in advanced colorectal cancer [75]. Similarly, mir-1275 improves esophageal cancer (EC) cells’ susceptibility to irradiation by suppressing EMT. Nonetheless, mir-1275 expression was down-regulated in the radiation-resistant esophageal cancer cell line KYSE-150R, and EMT was seen, leading to increased radiotherapy resistance in esophageal cancer cells [76] (Figure 2).
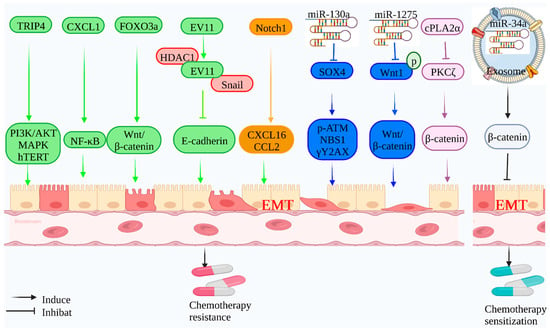
Figure 2.
Mechanism of EMT-mediated radiotherapy resistance: Tumor-associated genes can mediate the radiation resistance of tumor cells by inducing EMT. Tumor-related proteins (TRIP4, CXCL1, FOXO3a, Notch1, and cPLA2α) can induce EMT through related signaling pathways (PI3K/Akt, NF-κB, Wnt/β-catenin, CCL2/CXCL16, and PKCζ/β-catenin) and enhance the radiotherapy tolerance of cervical cancer, glioblastoma, and nasopharyngeal carcinoma. EVI1 inhibits E-cadherin expression and induces EMT by forming a co-inhibitory complex with Snail and HDAC1 and enhances the radiotherapy tolerance of nasopharyngeal carcinoma. Tumor-associated non-coding RNAs (Mir-130a and Mir-1275) inhibit target proteins (SOX4 and WNT1), further influencing related proteins (NBS1, P-ATM, and γH2AX) and related signaling pathways (Wnt/β-catenin) to induce EMT and enhance the radiotherapy sensitivity of colorectal cancer and esophageal cancer. Meanwhile, in response to EMT-mediated radiotherapy resistance, exosome Mir-34A can target the expression of β-catenin, delay the progression of EMT, and improve the radiotherapy sensitivity of NPC.
Inhibiting or reversing EMT may increase the radiosensitivity of cancers. Radiotherapy resistance in cervical cancer is intimately linked to cervical cancer stem cells. cPLA2 was discovered to regulate the reversible transition of cervical cancer stem cells between mesenchymal and epithelial states via the atypical protein kinase PKCζ, regulate cancer cell EMT state changes, and maintain various embryonic stem cells cell characteristics via the interaction between β-catenin and E-cadherin [77]. Consequently, EMT targeting cPLA2α to promote cervical cancer stem cells is a unique approach to radiosensitization therapy. Additionally, the exosome delivery of exogenous mir-34C to nasopharyngeal cancer may help to reduce EMT progression and increase radiation sensitivity [78].
4.3. EMT-Mediated Tumor-Targeted Therapy Resistance
With the rapid growth in molecular biology in cancer, molecular targeted therapy has been used in clinical practice and has gained growing importance in cancer medical treatment. While novel targets are rapidly being discovered, drug resistance remains a serious hurdle to the development of targeted therapies [79]. Cancer molecular targeted therapy is directed against tumor cells’ marker molecules in order to intervene in their carcinogenesis, such as inhibiting tumor cell proliferation, interfering with the cell cycle, inducing tumor cell differentiation and apoptosis, and inhibiting tumor cell metastasis and tumor vascular survival, to accomplish the purpose of tumor therapy. EMT may lead to drug resistance in tumor cells by modifying their behavior and function (for example, by promoting tumor spread, increasing drug efflux, or decreasing apoptotic signals), which is a crucial factor in drug resistance to targeted therapy [80,81].
At present, Gefitinib is the first-line therapeutic choice for patients with non-small cell lung cancer (NSCLC) who have an EGFR mutation. Gefitinib can promote tumor cell death and angiogenesis inhibition, but it may also delay the establishment of EMT in lung cancer cells by altering the link between HOTAIR and mir-34A-5p, hence exerting an additional effect antitumor activity [82]. This suggests that EMT may be closely related to the efficacy of EGFR-Tyrosine kinase inhibitors (EGFR-TKI). EMT is often associated with resistance to Gefitinib in lung cancer cells. By boosting interleukin-6 (IL-6) production in gefitinib-resistant non-small cell lung cancer cells, highly expressed LINC00460 acts as a competitive bait for mir-149-5p. Oct4 and Nanog coexpression affects drug resistance and EMT phenotypes by stimulating the Wnt/-βcatenin signaling pathway [83]. TGF-β1 overexpression has been demonstrated to promote ITG3 expression in patients with acquired Gefitinib or Oxitinib-resistant lung cancer. Antagonistic ITG3 inhibits lung cancer cell proliferation and EMT phenotypes, hence enhancing EGFR-TKI sensitivity [84]. Similar results were seen in gefitinib-resistant cell lines (HCC827GR and PC 9GR), where TGF-β1 -induced EMT contributed to gefitinib resistance through the mir-625-3p/AXL axis, and re-expression of the mir-625-3p partially restored gefitinib resistance. This further contributes to our knowledge of EGFR-TKI resistance [85].
Cetuximab, a monoclonal antibody directed against the EGFR, has been widely used to treat metastatic colorectal cancer; nevertheless, many patients who initially respond to cetuximab develop resistance [86]. Mir-141-3p participates in EMT by modulating the expression of E-cadherin, N-cadherin, Snail, and Vimentin. It also enhances the sensitivity of colorectal cancer cells to cetuximab by suppressing EGFR expression [87]. Meanwhile, RAS mutations limit the effectiveness of anti-EGFR monoclonal antibodies in patients receiving chemotherapy for metastatic colorectal cancer [88].
Curcumin is a plant polyphenol derived from the rhizome of turmeric, and its antitumor effect has been shown in liver cancer, lung cancer, and other malignant tumors. EMT is a critical biological link in the pathological process of esophageal cancer; it regulates the migration and invasion of cancer cells and has an indirect effect on the proliferation of cancer cells, chemotherapy resistance, and other biological behaviors [89]. During EMT, the epithelial phenotype is diminished along with the expression of the marker gene E-cadherin and cell polarity, enhancement of interstitial phenotype, increased expression of marker genes N-cadherin and Vimentin, and increased cell motility. Similarly, curcumin may enhance the expression of epithelial marker gene E-cadherin and reduce the expression of interstitial marker gene N-cadherin Vimentin; the greater the dosage of curcumin, the more pronounced the impact on gene expression regulation. These findings indicate that curcumin may greatly prevent EMT in esophageal cancer cells. EMT occurs in various malignant tumor lesions and is controlled by Notch and Wnt signaling pathways, according to publication number [90]. Notch1, Jagged1, Hes, Wnt1, and β-catenin were also reduced by curcumin in esophageal cancer cells. The Notch and Wnt pathway genes were evaluated after treatment with various dosages of curcumin, which increased the expression of GSK-3. These findings indicate that curcumin can greatly limit the activation of Notch and Wnt pathways in esophageal cancer cells and that curcumin may impede EMT by inhibiting Notch and Wnt pathways.
According to studies, resveratrol may reduce tumor invasion and metastasis by blocking EMT-related signaling pathways [91]. Resveratrol was reported to suppress the TGF-β-induced EMT process in lung cancer A549 cells by raising the expression of E-cadherin and lowering the expression of fibronectin, vimentin, and EMT-induced transcription factors Snail and Slug, consequently preventing cancer cell metastasis [92]. Likewise, Resveratrol suppresses TGF-β-induced EMT in colorectal cancer via reducing Smad protein, consequently limiting the invasion and metastasis of colorectal cancer cells. EMT is also proliferative vitreoretinopathy in retinal pigment epithelial cells. Important aspects of the pathogenesis Resveratrol may deacetylate Smad protein, hence reducing the EMT process generated by TGF-β and preventing proliferative vitreoretinopathy [93,94]. Table 1 below summarizes some molecular mechanisms by which EMT mediates cancer therapeutic resistance.

Table 1.
EMT-mediated therapeutic resistance.
4.4. EMT-Mediated Tumor Immunotherapy Resistance
Immune checkpoint inhibition is a significant advancement in tumor immunotherapy and is effective against a broad spectrum of advanced malignant tumors. However, low response rates and treatment resistance are essential hurdles to the advancement of immunotherapy. EMT has been associated with activating various immune checkpoint molecules, including programmed cell death ligand 1 (PD-L1) [97]. Thus, further exploration of the regulatory relationship between EMT and immune checkpoint molecules will improve immunotherapy’s sensitivity (Figure 3).
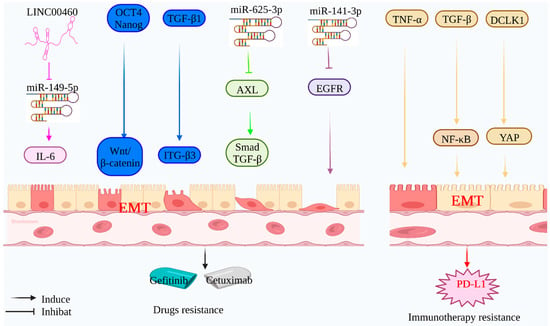
Figure 3.
Mechanism of EMT-mediated targeting and immunotherapy resistance: Tumor-associated genes can induce EMT-mediated tumor cell targeting and immunotherapy resistance. LINC00460 can competitively inhibit the expression of Mir-149-5p and induce EMT by regulating the expression of IL-6, thus mediating gefitinib resistance. Mir-625-3p inhibited the expression of target gene AXL and activated the TGF-β/Smad signaling pathway to induce EMT. Mir-141-3p can inhibit the expression of target gene EGFR and induce EMT, thus mediating the resistance of gefitinib and cetuximab. Tumor-associated proteins (Oct4, Nanog, and TGF-β1) can induce EMT through related signaling pathways (Wnt/β-catenin and ITGβ3) and mediate gefitinib resistance. Tumor-associated proteins (TNF-α, TGF-β1, and DCLK1) can induce EMT through related signaling pathways (NF-κB and YAP), thereby mediating immunotherapy resistance.
EMT endows tumor cells with the potential to avoid the immune system, which is controlled by immune checkpoint molecules in the majority of malignancies. This demonstrates a possible link between EMT and immunological checkpoint molecules. In hepatoma cell lines Hep3B and PLC/PRF/5, TNF-induced EMT increased the expression of PD-L1, PD-L2, CD73, and B7-H3, while reversing EMT reduced the expression of PD-L1, PD-L2, CD73, and B7-H3. Additionally, TNF was linked with a rise in PD-L1 expression in 422 HCC patients [96]. This study establishes a substantial link between the expression of immune checkpoint molecules and TNF-α-induced EMT. Similar incidences of gastric and pancreatic cancer have been reported. EMT may enhance the migration and invasion potential of gastric cancer cells, depending on the amount to which PD-L1 expression is enhanced due to NF-кB activation [95]. DCLK1 is integrally engaged in tumor cell EMT and may also regulate the expression of PD-L1 through the Hippo pathway-associated protein (YAP) [98].
Regulating EMT and the tumor immune response may be a potential strategy for increasing the efficacy of immunotherapy. CMTM6 is a PD-L1 regulatory molecule, and it has been shown that silencing CMTM6 decreases PD-L1 expression and increases CD8+ and CD4+ T cell infiltration. Meanwhile, via the Wnt/β-catenin signaling pathway, CMTM6 may increase tumor cell dehydration and EMT [99]. CMTM6 may be a potential target for immune sensitization therapy due to its capacity to simultaneously regulate tumor cell dryness, EMT, and immune response. REDOX states are present in melanoma and non-small cell lung cancer cell lines and are regulated by numerous aldehyde dehydrogenases (ALDHs). ALDH3A1 has been demonstrated to trigger EMT in melanoma and non-small cell lung cancer cells, while the overexpression of ALDH3A1 enhances PD-L1 expression and lowers monocyte proliferation in peripheral blood [90]. These results imply that ALDH3A1 is inextricably linked to EMT and PD-L1 and regulates tumor immune output.
5. Therapeutic Strategies for EMT-Mediated Tumor Therapeutic Resistance
Given the crucial role of EMT in tumor resistance, reversing or inhibiting EMT represents a unique idea for therapeutic tumor sensitization. MET is thought to be the diametric opposite of EMT. It leads to the suppression of mesenchymal characteristics of tumor cells and the re-expression of epithelial markers, which improves tumor cells’ resistance to therapy [100,101,102]. The conversion of mesenchymal tumor cells to an epithelioid phenotype may provide a unique method for cancer treatment resistance.
MET may be formed due to the reduced expression of EMT transcription factors (EMT-TRAN and transcription of EMT-TFS), and targeting EMT-TFS may be a strategy for treating malignancies that have developed resistance to treatment [103]. Ionizing radiation has been found to promote EMT and increase the radiation resistance of hypopharyngeal cancer cells through the AKT/GSK-3/Snail signaling pathway. However, silencing Snail reverses EMT and significantly lowers the radiation resistance of hypopharyngeal cancer cells [104]. It has been shown that silencing Slug increases cancer cells’ radiation sensitivity in oral squamous cell carcinoma [105]. These results suggest that Snail and Slug may be potential EMT-mediated radiation resistance therapy targets. Twist may form a dimer or heterodimer with the E-cadherin promoter region, inhibiting its expression and promoting the initiation of EMT [106]. When sitwist-MSN-HA, a novel nanoparticle delivery platform, interfered with Twist expression, naked mice were more vulnerable to cisplatin in epithelial ovarian cancer, showing that the technology may have been exploited in malignancies with the overexpression of Twist [107]. Not only is ZEB1 required for EMT activation, but it also plays a crucial role in the development of chemotherapeutic resistance. Interfering with ZEB1 expression has been demonstrated to increase the sensitivity of colorectal cancer cells to chemotherapy, suggesting that reducing ZEB1 expression may help restore sensitivity in ZEB1-mediated chemotherapy resistance [108].
Numerous signaling pathways are implicated in tumor cell EMT, and inhibitors of these signaling pathways may effectively delay EMT to a certain extent, hence diminishing tumor resistance to treatment. TGF-β/Smad signaling is a well-characterized signaling mechanism that regulates EMT. The TGF-receptor inhibitors LV2109761 and LV364947 have been demonstrated to block EMT generated by ionizing radiation in gastric cancer and glioma cells, hence boosting tumor cells’ irradiation sensitivity [109,110,111]. Additionally, tumor EMT and treatment resistance are related to the Wnt/β-catenin signaling pathway [112]. Indeed, the overexpression of the Wnt/β-catenin signaling pathway has been demonstrated to promote tumor cell EMT and resistance to treatment. WNT974 inhibits the Wnt/β-catenin signaling system, which may significantly increase lymphoma chemotherapeutic sensitivity to driamycin. The inhibitor XAV939 has dramatically increased cervical cancer cells’ radiation sensitivity [113,114].
Several recent studies demonstrated a correlation between EMT and the regulation of certain ribosomal proteins in various forms of cancer. Some ribosomal proteins have shown the potential to regulate cell migration, altering the EMT process and eventually resulting in chemoresistance [115,116]. The down-regulation of ribosomal protein uL3 is associated with increased cell migration and an EMT resulting in chemoresistance, while the silencing of ribosomal protein RPL34 is sufficient to reduce the EMT phenotype, preventing esophageal cancer cell migration and invasion [117,118]. The translation of ribosomal protein-coding mRNAs (RP-mRNAs) causes the creation of ribosomes; nevertheless, the mechanisms that regulate RP-mRNA translation in conjunction with other cellular activities are not well understood. The RP-mRNAs are localized in actin-rich cell protrusions during cell migration into their surroundings. RNA-binding protein La-related protein 6 is responsible for this localization (LARP6). Cell migration depends on LARP6-mediated mRNA localization, and this pathway is related to cancer development during EMT [116,119].
6. Outlooks
The mechanism by which tumor cells develop treatment resistance is complex, and therapeutic efficiency and resistance processes differ significantly across individuals with diverse tumors. The heterogeneity and biological mechanisms such as EMT, autophagy, cell cycle arrest, DNA damage repair, drug metabolism, and efflux all contribute to tumor treatment resistance. Additionally, the tumor microenvironment and other small-molecule compounds in the circulatory system may contribute to the resistance management of tumor therapy. However, intervening in only one of the steps outlined above is insufficient to completely reverse tumor cell treatment resistance: future research should focus on the underlying mechanism of tumor cells from the treatment-resistant phenotype. EMT is one of the most common mechanisms underlying resistance to cancer treatment. As research has advanced, the role of EMT and the molecular mechanism by which EMT occurs have become well understood. Numerous clinical studies have shown a clear link between EMT, chemotherapy tolerance, and radiation resistance in cancer patients. However, there is currently a shortage of therapeutic drugs or treatment techniques that may boost the EMT process and hence the efficiency of tumor therapy. Although cell and animal studies have shown that targeting specific locations may reverse EMT and boost tumor therapy sensitivity, there are still considerable risks and obstacles to long-term human success. As a result, a more comprehensive and detailed study of the relationship between EMT and tumor treatment resistance is necessary. It is still necessary to coordinate basic experiments, clinical research, and development with translational medicine at the core to develop a new plan for personalized tumor treatment in the future.
Author Contributions
Conceptualization Z.X. and Y.Z.; investigation Z.X., Y.Z. and H.D.; original draft preparation Z.X., Y.Z., H.D. and B.H.; review and editing Z.X. and B.H.; funding acquisition Z.X. and B.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the International Cooperation Project of Science and Technology Development Jilin Province (20190701035GH). Project of Jilin Provincial Department of Finance (jcsz2021893–19).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial-mesenchymal transition and implica-tions for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Buhrmann, C.; Brockmueller, A.; Harsha, C.; Kunnumakkara, A.B.; Kubatka, P.; Aggarwal, B.B.; Shakibaei, M. Evidence That Tumor Microenvironment Initiates Epithelial-To-Mesenchymal Transition and Calebin A can Suppress it in Colorectal Cancer Cells. Front. Pharmacol. 2021, 12, 699842. [Google Scholar] [CrossRef]
- Khodabandeh, Z.; Valilo, M.; Velaei, K.; Pirpour Tazehkand, A. The potential role of nicotine in breast cancer initiation, development, angiogenesis, invasion, metastasis, and resistance to therapy. Breast Cancer 2022, 1–12. [Google Scholar] [CrossRef]
- Yang, R.; Yi, M.; Xiang, B. Novel Insights on Lipid Metabolism Alterations in Drug Resistance in Cancer. Front. Cell Dev. Biol. 2022, 10, 875318. [Google Scholar] [CrossRef]
- Wu, J.; He, J.; Zhang, J.; Ji, H.; Wang, N.; Ma, S.; Yan, X.; Gao, X.; Du, J.; Liu, Z.; et al. Identification of EMT-Related Genes and Prognostic Signature With Significant Implications on Biological Properties and Oncology Treatment of Lower Grade Gliomas. Front. Cell Dev. Biol. 2022, 10, 887693. [Google Scholar] [CrossRef]
- Lee, C.C.; Cheng, Y.C.; Chang, C.Y.; Lin, C.M.; Chang, J.Y. Alpha-tubulin acetyltransferase/MEC-17 regulates cancer cell migration and invasion through epithelial-mesenchymal transition suppression and cell polarity disruption. Sci. Rep. 2018, 8, 17477. [Google Scholar] [CrossRef]
- Kong, D.; Zhou, H.; Neelakantan, D.; Hughes, C.J.; Hsu, J.Y.; Srinivasan, R.R.; Lewis, M.T.; Ford, H.L. VEGF-C mediates tumor growth and metastasis through promoting EMT-epithelial breast cancer cell crosstalk. Oncogene 2021, 40, 964–979. [Google Scholar] [CrossRef]
- Lindner, P.; Paul, S.; Eckstein, M.; Hampel, C.; Muenzner, J.K.; Erlenbach-Wuensch, K.; Ahmed, H.P.; Mahadevan, V.; Brabletz, T.; Hartmann, A.; et al. EMT transcription factor ZEB1 alters the epigenetic landscape of colorectal cancer cells. Cell Death Dis. 2020, 11, 147. [Google Scholar] [CrossRef]
- Caramel, J.; Ligier, M.; Puisieux, A. Pleiotropic Roles for ZEB1 in Cancer. Cancer Res. 2018, 78, 30–35. [Google Scholar] [CrossRef]
- Song, H.; Liu, D.; Dong, S.; Zeng, L.; Wu, Z.; Zhao, P.; Zhang, L.; Chen, Z.S.; Zou, C. Epitranscriptomics and epiproteomics in cancer drug resistance: Therapeutic implications. Signal Transduct. Target. Ther. 2020, 5, 193. [Google Scholar] [CrossRef]
- Weadick, B.; Nayak, D.; Persaud, A.K.; Hung, S.W.; Raj, R.; Campbell, M.J.; Chen, W.; Li, J.; Williams, T.M.; Govindarajan, R. EMT-Induced Gemcitabine Resistance in Pancreatic Cancer Involves the Functional Loss of Equilibrative Nucleoside Transporter 1. Mol. Cancer Ther. 2021, 20, 410–422. [Google Scholar] [CrossRef]
- Luo, M.; Wu, C.; Guo, E.; Peng, S.; Zhang, L.; Sun, W.; Liu, D.; Hu, G.; Hu, G. FOXO3a knockdown promotes radioresistance in nasopharyngeal carcinoma by inducing epithelial-mesenchymal transition and the Wnt/beta-catenin signaling pathway. Cancer Lett. 2019, 455, 26–35. [Google Scholar] [CrossRef]
- Takaoka, Y.; Konno, M.; Koseki, J.; Colvin, H.; Asai, A.; Tamari, K.; Satoh, T.; Mori, M.; Doki, Y.; Ogawa, K.; et al. Mitochondrial pyruvate carrier 1 expression controls cancer epithelial-mesenchymal transition and radioresistance. Cancer Sci. 2019, 110, 1331–1339. [Google Scholar] [CrossRef]
- Owusu-Akyaw, A.; Krishnamoorthy, K.; Goldsmith, L.T.; Morelli, S.S. The role of mesenchymal-epithelial transition in endometrial function. Hum. Reprod. Update 2019, 25, 114–133. [Google Scholar] [CrossRef]
- Hill, C.; Li, J.; Liu, D.; Conforti, F.; Brereton, C.J.; Yao, L.; Zhou, Y.; Alzetani, A.; Chee, S.J.; Marshall, B.G.; et al. Autophagy inhibition-mediated epithelial-mesenchymal transition augments local myofibroblast differentiation in pulmonary fibrosis. Cell Death Dis. 2019, 10, 591. [Google Scholar] [CrossRef]
- VanDussen, K.L.; Stojmirovic, A.; Li, K.; Liu, T.C.; Kimes, P.K.; Muegge, B.D.; Simpson, K.F.; Ciorba, M.A.; Perrigoue, J.G.; Friedman, J.R.; et al. Abnormal Small Intestinal Epithelial Microvilli in Patients with Crohn’s Disease. Gastroenterology 2018, 155, 815–828. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, M.; Zhou, C.; Wang, W.; Yang, H.; Ye, W. The role of epithelial-mesenchymal transition in regulating radioresistance. Crit. Rev. Oncol. Hematol. 2020, 150, 102961. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Xue, H.; Li, C.; Liu, Q.; Zhou, Y.; Wang, T.; Wang, H.; Qian, H.; Wen, T. A TGF-beta-MTA1-SOX4-EZH2 signaling axis drives epithelial-mesenchymal transition in tumor metastasis. Oncogene 2020, 39, 2125–2139. [Google Scholar] [CrossRef]
- Du, F.; Yu, L.; Wu, Y.; Wang, S.; Yao, J.; Zheng, X.; Xie, S.; Zhang, S.; Lu, X.; Liu, Y.; et al. miR-137 alleviates doxorubicin resistance in breast cancer through inhibition of epithelial-mesenchymal transition by targeting DUSP4. Cell Death Dis. 2019, 10, 922. [Google Scholar] [CrossRef]
- Cheng, J.; Meng, J.; Zhu, L.; Peng, Y. Exosomal noncoding RNAs in Glioma: Biological functions and potential clinical applications. Mol. Cancer 2020, 19, 66. [Google Scholar] [CrossRef]
- Wei, L.; Sun, J.; Zhang, N.; Zheng, Y.; Wang, X.; Lv, L.; Liu, J.; Xu, Y.; Shen, Y.; Yang, M. Noncoding RNAs in gastric cancer: Implications for drug resistance. Mol. Cancer 2020, 19, 62. [Google Scholar] [CrossRef]
- Hu, Q.; Huang, Z.; Duan, Y.; Fu, Z.; Bin, L. Reprogramming Tumor Microenvironment with Photothermal Therapy. Bioconjug. Chem. 2020, 31, 1268–1278. [Google Scholar] [CrossRef]
- Zeng, D.; Liang, Y.K.; Xiao, Y.S.; Wei, X.L.; Lin, H.Y.; Wu, Y.; Bai, J.W.; Chen, M.; Zhang, G.J. Inhibition of Notch1 reverses EMT and chemoresistance to cisplatin via direct downregulation of MCAM in triple-negative breast cancer cells. Int. J. Cancer 2020, 147, 490–504. [Google Scholar] [CrossRef]
- El Hout, M.; Cosialls, E.; Mehrpour, M.; Hamai, A. Crosstalk between autophagy and metabolic regulation of cancer stem cells. Mol. Cancer 2020, 19, 27. [Google Scholar] [CrossRef]
- Li, Y.; He, Y.; Butler, W.; Xu, L.; Chang, Y.; Lei, K.; Zhang, H.; Zhou, Y.; Gao, A.C.; Zhang, Q.; et al. Targeting cellular heterogeneity with CXCR2 blockade for the treatment of therapy-resistant prostate cancer. Sci. Transl. Med. 2019, 11, eaax0428. [Google Scholar] [CrossRef]
- Martins-Neves, S.R.; Cleton-Jansen, A.M.; Gomes, C.M.F. Therapy-induced enrichment of cancer stem-like cells in solid human tumors: Where do we stand? Pharmacol. Res. 2018, 137, 193–204. [Google Scholar] [CrossRef]
- Marzagalli, M.; Raimondi, M.; Fontana, F.; Montagnani Marelli, M.; Moretti, R.M.; Limonta, P. Cellular and molecular biology of cancer stem cells in melanoma: Possible therapeutic implications. Semin. Cancer Biol. 2019, 59, 221–235. [Google Scholar] [CrossRef]
- Fung, S.W.; Cheung, P.F.; Yip, C.W.; Ng, L.W.; Cheung, T.T.; Chong, C.C.; Lee, C.; Lai, P.B.; Chan, A.W.; Tsao, G.S.; et al. The ATP-binding cassette transporter ABCF1 is a hepatic oncofetal protein that promotes chemoresistance, EMT and cancer stemness in hepatocellular carcinoma. Cancer Lett. 2019, 457, 98–109. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Chen, X. Drug resistance and combating drug resistance in cancer. Cancer Drug Resist. 2019, 2, 141–160. [Google Scholar] [CrossRef]
- Alfarouk, K.O.; Stock, C.M.; Taylor, S.; Walsh, M.; Muddathir, A.K.; Verduzco, D.; Bashir, A.H.; Mohammed, O.Y.; El-hassan, G.O.; Harguindey, S.; et al. Resistance to cancer chemotherapy: Failure in drug response from ADME to P-gp. Cancer Cell Int. 2015, 15, 71. [Google Scholar] [CrossRef]
- Sui, H.; Fan, Z.Z.; Li, Q. Signal transduction pathways and transcriptional mechanisms of ABCB1/Pgp-mediated multiple drug resistance in human cancer cells. J. Int. Med. Res. 2012, 40, 426–435. [Google Scholar] [CrossRef]
- Liu, X. ABC Family Transporters. Adv. Exp. Med. Biol. 2019, 1141, 13–100. [Google Scholar] [CrossRef]
- Zattoni, I.F.; Delabio, L.C.; Dutra, J.P.; Kita, D.H.; Scheiffer, G.; Hembecker, M.; Pereira, G.D.S.; Moure, V.R.; Valdameri, G. Targeting breast cancer resistance protein (BCRP/ABCG2): Functional inhibitors and expression modulators. Eur. J. Med. Chem. 2022, 237, 114346. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; Stephens, M.A.; Pathak, H.; Rangarajan, A. Transcription factors that mediate epithelial-mesenchymal transition lead to multidrug resistance by upregulating ABC transporters. Cell Death Dis. 2011, 2, e179. [Google Scholar] [CrossRef]
- Li, Q.Q.; Xu, J.D.; Wang, W.J.; Cao, X.X.; Chen, Q.; Tang, F.; Chen, Z.Q.; Liu, X.P.; Xu, Z.D. Twist1-mediated adriamy-cin-induced epithelial-mesenchymal transition relates to multidrug resistance and invasive potential in breast cancer cells. Clin. Cancer Res. 2009, 15, 2657–2665. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.S.; Sun, Y.Z.; Wang, S.M.; Ruan, J.S. Epithelial-mesenchymal transition: Potential regulator of ABC trans-porters in tumor progression. J. Cancer 2017, 8, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Aberuyi, N.; Rahgozar, S.; Pourabutaleb, E.; Ghaedi, K. Selective dysregulation of ABC transporters in methotrex-ate-resistant leukemia T-cells can confer cross-resistance to cytarabine, vincristine and dexamethasone, but not doxorubicin. Curr. Res. Transl. Med. 2021, 69, 103269. [Google Scholar] [CrossRef]
- Tan, S.; Xia, L.; Yi, P.; Han, Y.; Tang, L.; Pan, Q.; Tian, Y.; Rao, S.; Oyang, L.; Liang, J.; et al. Exosomal miRNAs in tumor microenvironment. J. Exp. Clin. Cancer Res. 2020, 39, 67. [Google Scholar] [CrossRef]
- Xiao, Y.; Yu, D. Tumor microenvironment as a therapeutic target in cancer. Pharmacol. Ther. 2021, 221, 107753. [Google Scholar] [CrossRef]
- Krisnawan, V.E.; Stanley, J.A.; Schwarz, J.K.; DeNardo, D.G. Tumor Microenvironment as a Regulator of Radiation Therapy: New Insights into Stromal-Mediated Radioresistance. Cancers 2020, 12, 2916. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, T.; Liu, R.; Ning, T.; Yang, H.; Liu, D.; Zhang, Q.; Lin, D.; Ge, S.; Bai, M.; et al. CAF secreted miR-522 suppresses ferroptosis and promotes acquired chemo-resistance in gastric cancer. Mol. Cancer 2020, 19, 43. [Google Scholar] [CrossRef] [PubMed]
- Okoye, I.S.; Coomes, S.M.; Pelly, V.S.; Czieso, S.; Papayannopoulos, V.; Tolmachova, T.; Seabra, M.C.; Wilson, M.S. MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity 2014, 41, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Gu, Y.; Lu, P.; Li, J.; Cao, H.; Li, X.; Qian, X.; Yu, C.; Yang, Y.; Yang, X.; et al. Exosome-mediated lncRNA AFAP1-AS1 promotes trastuzumab resistance through binding with AUF1 and activating ERBB2 translation. Mol. Cancer 2020, 19, 26. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Qin, Z.; Cai, S.; Yu, L.; Hu, H.; Zeng, S. The role of non-coding RNAs in ABC transporters regulation and their clinical implications of multidrug resistance in cancer. Expert Opin. Drug Metab. Toxicol. 2021, 17, 291–306. [Google Scholar] [CrossRef]
- Zeng, F.; Wang, Q.; Wang, S.; Liang, S.; Huang, W.; Guo, Y.; Peng, J.; Li, M.; Zhu, W.; Guo, L. Linc00173 promotes chemoresistance and progression of small cell lung cancer by sponging miR-218 to regulate Etk expression. Oncogene 2020, 39, 293–307. [Google Scholar] [CrossRef]
- Hong, X.; Liu, N.; Liang, Y.; He, Q.; Yang, X.; Lei, Y.; Zhang, P.; Zhao, Y.; He, S.; Wang, Y.; et al. Circular RNA CRIM1 functions as a ceRNA to promote nasopharyngeal carcinoma metastasis and docetaxel chemoresistance through upregulating FOXQ1. Mol. Cancer 2020, 19, 33. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.Y.; Zu, Y.X.; Jiang, X.W.; Sun, X.T.; Liu, T.Y.; Li, R.L.; Wu, Q.; Zhang, Y.S.; Zhao, Q.C. Novel ADAM-17 inhibitor ZLDI-8 inhibits the proliferation and metastasis of chemo-resistant non-small-cell lung cancer by reversing Notch and epithelial mesenchymal transition in vitro and in vivo. Pharmacol. Res. 2019, 148, 104406. [Google Scholar] [CrossRef]
- Williams, E.D.; Gao, D.; Redfern, A.; Thompson, E.W. Controversies around epithelial-mesenchymal plasticity in cancer metastasis. Nat. Rev. Cancer 2019, 19, 716–732. [Google Scholar] [CrossRef]
- Blazquez, R.; Rietkotter, E.; Wenske, B.; Wlochowitz, D.; Sparrer, D.; Vollmer, E.; Muller, G.; Seegerer, J.; Sun, X.; Dettmer, K.; et al. LEF1 supports metastatic brain colonization by regulating glutathione metabolism and increasing ROS resistance in breast cancer. Int. J. Cancer 2020, 146, 3170–3183. [Google Scholar] [CrossRef]
- Pattabiraman, D.R.; Bierie, B.; Kober, K.I.; Thiru, P.; Krall, J.A.; Zill, C.; Reinhardt, F.; Tam, W.L.; Weinberg, R.A. Activation of PKA leads to mesenchymal-to-epithelial transition and loss of tumor-initiating ability. Science 2016, 351, aad3680. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, Y.; Dong, Z.; Shi, Z.; Zhang, J.T. Single-nucleotide polymorphisms in a short basic motif in the ABC transporter ABCG2 disable its trafficking out of endoplasmic reticulum and reduce cell resistance to anticancer drugs. J. Biol. Chem. 2019, 294, 20222–20232. [Google Scholar] [CrossRef] [PubMed]
- Nedeljkovic, M.; Damjanovic, A. Mechanisms of Chemotherapy Resistance in Triple-Negative Breast Cancer-How We Can Rise to the Challenge. Cells 2019, 8, 957. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Li, S.; Tang, X.; Wang, Y.; Guo, W.; Cao, G.; Chen, K.; Zhang, M.; Guan, M.; Yang, D. Copy Number Amplification of DNA Damage Repair Pathways Potentiates Therapeutic Resistance in Cancer. Theranostics 2020, 10, 3939–3951. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, D.; Wu, J.Y.; Xing, K.; Yeo, E.; Li, C.; Zhang, L.; Holland, E.; Yao, L.; Qin, L.; et al. Wnt-mediated endothelial transformation into mesenchymal stem cell-like cells induces chemoresistance in glioblastoma. Sci. Transl. Med. 2020, 12, eaay7522. [Google Scholar] [CrossRef] [PubMed]
- Giannone, G.; Attademo, L.; Scotto, G.; Genta, S.; Ghisoni, E.; Tuninetti, V.; Aglietta, M.; Pignata, S.; Valabrega, G. Endometrial Cancer Stem Cells: Role, Characterization and Therapeutic Implications. Cancers 2019, 11, 1820. [Google Scholar] [CrossRef]
- Horn, L.A.; Fousek, K.; Palena, C. Tumor Plasticity and Resistance to Immunotherapy. Trends Cancer 2020, 6, 432–441. [Google Scholar] [CrossRef]
- Navas, T.; Kinders, R.J.; Lawrence, S.M.; Ferry-Galow, K.V.; Borgel, S.; Hollingshead, M.G.; Srivastava, A.K.; Alcoser, S.Y.; Makhlouf, H.R.; Chuaqui, R.; et al. Clinical Evolution of Epithelial-Mesenchymal Transition in Human Carcinomas. Cancer Res. 2020, 80, 304–318. [Google Scholar] [CrossRef]
- Tan, X.; Li, Z.; Ren, S.; Rezaei, K.; Pan, Q.; Goldstein, A.T.; Macri, C.J.; Cao, D.; Brem, R.F.; Fu, S.W. Dynamically decreased miR-671-5p expression is associated with oncogenic transformation and radiochemoresistance in breast cancer. Breast Cancer Res. 2019, 21, 89. [Google Scholar] [CrossRef]
- Kirave, P.; Gondaliya, P.; Kulkarni, B.; Rawal, R.; Garg, R.; Jain, A.; Kalia, K. Exosome mediated miR-155 delivery confers cisplatin chemoresistance in oral cancer cells via epithelial-mesenchymal transition. Oncotarget 2020, 11, 1157–1171. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, R.; An, X.; Li, Z.; Fang, C.; Pan, B.; Chen, W.; Xu, G.; Han, W. LncRNA HOXA-AS3 confers cisplatin resistance by interacting with HOXA3 in non-small-cell lung carcinoma cells. Oncogenesis 2019, 8, 60. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Z.; Sun, J.; Lv, H.; Wang, Y.; Ni, Y.; Chen, S.; Hu, C.; Wang, L.; Chen, W.; et al. Cisplatin resistance in gastric cancer cells is involved with GPR30-mediated epithelial-mesenchymal transition. J. Cell. Mol. Med. 2020, 24, 3625–3633. [Google Scholar] [CrossRef] [PubMed]
- Lionarons, D.A.; Hancock, D.C.; Rana, S.; East, P.; Moore, C.; Murillo, M.M.; Carvalho, J.; Spencer-Dene, B.; Herbert, E.; Stamp, G.; et al. RAC1(P29S) Induces a Mesenchymal Phenotypic Switch via Serum Response Factor to Promote Melanoma Development and Therapy Resistance. Cancer Cell 2019, 36, 68–83.e69. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, J.M.; Wei, W.; Yang, R.; Chen, D.; Ma, X.D.; Jiang, G.M.; Wang, B.L. Regulation of ATP-binding cassette subfamily B member 1 by Snail contributes to chemoresistance in colorectal cancer. Cancer Sci. 2020, 111, 84–97. [Google Scholar] [CrossRef]
- Mendoza-Rodriguez, M.G.; Sanchez-Barrera, C.A.; Callejas, B.E.; Garcia-Castillo, V.; Beristain-Terrazas, D.L.; Delgado-Buenrostro, N.L.; Chirino, Y.I.; Leon-Cabrera, S.A.; Rodriguez-Sosa, M.; Gutierrez-Cirlos, E.B.; et al. Use of STAT6 Phosphorylation Inhibitor and Trimethylglycine as New Adjuvant Therapies for 5-Fluorouracil in Colitis-Associated Tumorigenesis. Int. J. Mol. Sci. 2020, 21, 2130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cao, T.; Guo, K.; Zhou, Y.; Liu, H.; Pan, Y.; Hou, Q.; Nie, Y.; Fan, D.; Lu, Y.; et al. Regulation of Integrin Subunit Alpha 2 by miR-135b-5p Modulates Chemoresistance in Gastric Cancer. Front. Oncol. 2020, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Schulz, A.; Meyer, F.; Dubrovska, A.; Borgmann, K. Cancer Stem Cells and Radioresistance: DNA Repair and Beyond. Cancers 2019, 11, 862. [Google Scholar] [CrossRef]
- Simonet, S.; Rodriguez-Lafrasse, C.; Beal, D.; Gerbaud, S.; Malesys, C.; Tillement, O.; Lux, F.; Fayyad-Kazan, H.; Rachidi, W.; Ardail, D. Gadolinium-Based Nanoparticles Can Overcome the Radioresistance of Head and Neck Squamous Cell Carcinoma Through the Induction of Autophagy. J. Biomed. Nanotechnol. 2020, 16, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liu, H.; Zheng, Y.; Han, Y.; Wang, T.; Zhang, H.; Sun, Q.; Li, Z. Overcoming Radioresistance in Tumor Therapy by Alleviating Hypoxia and Using the HIF-1 Inhibitor. ACS Appl. Mater. Interfaces 2020, 12, 4231–4240. [Google Scholar] [CrossRef]
- Tsao, T.; Beretov, J.; Ni, J.; Bai, X.; Bucci, J.; Graham, P.; Li, Y. Cancer stem cells in prostate cancer radioresistance. Cancer Lett. 2019, 465, 94–104. [Google Scholar] [CrossRef]
- Yadav, S.; Kowolik, C.M.; Lin, M.; Zuro, D.; Hui, S.K.; Riggs, A.D.; Horne, D.A. SMC1A is associated with radioresistance in prostate cancer and acts by regulating epithelial-mesenchymal transition and cancer stem-like properties. Mol. Carcinog. 2019, 58, 113–125. [Google Scholar] [CrossRef]
- Che, Y.; Li, Y.; Zheng, F.; Zou, K.; Li, Z.; Chen, M.; Hu, S.; Tian, C.; Yu, W.; Guo, W.; et al. TRIP4 promotes tumor growth and metastasis and regulates radiosensitivity of cervical cancer by activating MAPK, PI3K/AKT, and hTERT signaling. Cancer Lett. 2019, 452, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Alafate, W.; Li, X.; Zuo, J.; Zhang, H.; Xiang, J.; Wu, W.; Xie, W.; Bai, X.; Wang, M.; Wang, J. Elevation of CXCL1 indicates poor prognosis and radioresistance by inducing mesenchymal transition in glioblastoma. CNS Neurosci. Ther. 2020, 26, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liang, Y.; Zheng, X.; Deng, X.; Huang, W.; Zhang, G. EVI1 promotes epithelial-to-mesenchymal transition, cancer stem cell features and chemo-/radioresistance in nasopharyngeal carcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 82. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Wang, F.; Diao, Y.; Zhang, Z.; Chen, Q.; Qian, C.N.; Keller, E.T.; Zhang, J.; Lu, Y. Knockdown of Notch1 inhibits nasopharyngeal carcinoma cell growth and metastasis via downregulation of CCL2, CXCL16, and uPA. Mol. Carcinog. 2019, 58, 1886–1896. [Google Scholar] [CrossRef] [PubMed]
- Ha Thi, H.T.; Kim, H.Y.; Kim, Y.M.; Hong, S. MicroRNA-130a modulates a radiosensitivity of rectal cancer by targeting SOX4. Neoplasia 2019, 21, 882–892. [Google Scholar] [CrossRef]
- Xie, C.; Wu, Y.; Fei, Z.; Fang, Y.; Xiao, S.; Su, H. MicroRNA-1275 induces radiosensitization in oesophageal cancer by regulating epithelial-to-mesenchymal transition via Wnt/beta-catenin pathway. J. Cell. Mol. Med. 2020, 24, 747–759. [Google Scholar] [CrossRef]
- He, Y.; Xiao, M.; Fu, H.; Chen, L.; Qi, L.; Liu, D.; Guo, P.; Chen, L.; Luo, Y.; Xiao, H.; et al. cPLA2alpha reversibly regulates different subsets of cancer stem cells transformation in cervical cancer. Stem Cells 2020, 38, 487–503. [Google Scholar] [CrossRef]
- Wan, F.Z.; Chen, K.H.; Sun, Y.C.; Chen, X.C.; Liang, R.B.; Chen, L.; Zhu, X.D. Exosomes overexpressing miR-34c inhibit malignant behavior and reverse the radioresistance of nasopharyngeal carcinoma. J. Transl. Med. 2020, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Weygant, N.; Chandrakesan, P.; Houchen, C.W.; Peng, J.; Qu, D. Tuft and Cancer Stem Cell Marker DCLK1: A New Target to Enhance Anti-Tumor Immunity in the Tumor Microenvironment. Cancers 2020, 12, 3801. [Google Scholar] [CrossRef]
- Bonavida, B.; Baritaki, S. Dual role of NO donors in the reversal of tumor cell resistance and EMT: Downregulation of the NF-kappaB/Snail/YY1/RKIP circuitry. Nitric Oxide 2011, 24, 1–7. [Google Scholar] [CrossRef]
- Liu, H.; Chen, C.; Zeng, J.; Zhao, Z.; Hu, Q. MicroRNA-210-3p is transcriptionally upregulated by hypoxia induction and thus promoting EMT and chemoresistance in glioma cells. PLoS ONE 2021, 16, e0253522. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Li, J.; Ma, C.; Tang, X.; Tang, Q.; Wu, J.; Chai, X.; Xie, J.; Yang, X.B.; Hann, S.S. Novel regulation of miR-34a-5p and HOTAIR by the combination of berberine and gefitinib leading to inhibition of EMT in human lung cancer. J. Cell. Mol. Med. 2020, 24, 5578–5592. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhu, H.; Liao, Y.; Wu, W.; Liu, L.; Liu, L.; Wu, Y.; Sun, F.; Lin, H.W. Inhibition of Wnt/beta-catenin pathway reverses multi-drug resistance and EMT in Oct4(+)/Nanog(+) NSCLC cells. Biomed. Pharmacother. 2020, 127, 110225. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, T.; Lv, D.; Li, L.; Yue, J.; Chen, H.Z.; Xu, L. Acquired Resistance to EGFR TKIs Mediated by TGFbeta1/Integrin beta3 Signaling in EGFR-Mutant Lung Cancer. Mol. Cancer Ther. 2019, 18, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Sun, L.; Liu, T.; Zhu, J.; Zeng, Y.; Zhang, Y.; Wang, X.; Liu, Z.; Huang, J.A. The miR6253p/AXL axis induces nonT790M acquired resistance to EGFRTKI via activation of the TGFbeta/Smad pathway and EMT in EGFRmutant nonsmall cell lung cancer. Oncol. Rep. 2020, 44, 185–195. [Google Scholar] [CrossRef]
- Gao, L.; Xu, J.; He, G.; Huang, J.; Xu, W.; Qin, J.; Zheng, P.; Ji, M.; Chang, W.; Ren, L.; et al. CCR7 high expression leads to cetuximab resistance by cross-talking with EGFR pathway in PI3K/AKT signals in colorectal cancer. Am. J. Cancer Res. 2019, 9, 2531–2543. [Google Scholar]
- Xing, Y.; Jing, H.; Zhang, Y.; Suo, J.; Qian, M. MicroRNA-141-3p affected proliferation, chemosensitivity, migration and invasion of colorectal cancer cells by targeting EGFR. Int. J. Biochem. Cell Biol. 2020, 118, 105643. [Google Scholar] [CrossRef]
- Birkman, E.M.; Avoranta, T.; Algars, A.; Korkeila, E.; Lintunen, M.; Lahtinen, L.; Kuopio, T.; Ristamaki, R.; Carpen, O.; Sundstrom, J. EGFR gene copy number decreases during anti-EGFR antibody therapy in colorectal cancer. Hum. Pathol. 2018, 82, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Hosoda, K.; Yashima, K.; Tamoto, A.; Yamamoto, S.; Kawata, S.; Ikebuchi, Y.; Matsumoto, K.; Kawaguchi, K.; Harada, K.; Murawaki, Y.; et al. Expression of methylation-modulated tumor-related genes in endoscopically resected early esophageal squamous neoplasia. Oncol. Lett. 2017, 14, 737–742. [Google Scholar] [CrossRef][Green Version]
- Yokozaki, H.; Koma, Y.I.; Shigeoka, M.; Nishio, M. Cancer as a tissue: The significance of cancer-stromal interactions in the development, morphogenesis and progression of human upper digestive tract cancer. Pathol. Int. 2018, 68, 334–352. [Google Scholar] [CrossRef]
- Liu, C.Y.; Lin, H.H.; Tang, M.J.; Wang, Y.K. Vimentin contributes to epithelial-mesenchymal transition cancer cell mechanics by mediating cytoskeletal organization and focal adhesion maturation. Oncotarget 2015, 6, 15966–15983. [Google Scholar] [CrossRef] [PubMed]
- Uttamsingh, S.; Bao, X.; Nguyen, K.T.; Bhanot, M.; Gong, J.; Chan, J.L.; Liu, F.; Chu, T.T.; Wang, L.H. Synergistic effect between EGF and TGF-beta1 in inducing oncogenic properties of intestinal epithelial cells. Oncogene 2008, 27, 2626–2634. [Google Scholar] [CrossRef]
- Ji, Q.; Liu, X.; Han, Z.; Zhou, L.; Sui, H.; Yan, L.; Jiang, H.; Ren, J.; Cai, J.; Li, Q. Resveratrol suppresses epithelial-to-mesenchymal transition in colorectal cancer through TGF-beta1/Smads signaling pathway mediated Snail/E-cadherin expression. BMC Cancer 2015, 15, 97. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, H.; Tang, L.; Chen, H.; Wu, C.; Zhao, M.; Yang, Y.; Chen, X.; Liu, G. Resveratrol inhibits TGF-beta1-induced epithelial-to-mesenchymal transition and suppresses lung cancer invasion and metastasis. Toxicology 2013, 303, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Li, J.; Li, R.Y.; Lan, T.; Xiao, C.; Gong, P. PD-L1 Expression Is Regulated By NF-kappaB During EMT Signaling In Gastric Carcinoma. Onco Targets Ther. 2019, 12, 10099–10105. [Google Scholar] [CrossRef]
- Shrestha, R.; Bridle, K.R.; Crawford, D.H.G.; Jayachandran, A. TNFalphamediated epithelialtomesenchymal transition regulates expression of immune checkpoint molecules in hepatocellular carcinoma. Mol. Med. Rep. 2020, 21, 1849–1860. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhan, H. Communication between EMT and PD-L1 signaling: New insights into tumor immune evasion. Cancer Lett. 2020, 468, 72–81. [Google Scholar] [CrossRef]
- Yan, R.; Li, J.; Zhou, Y.; Yao, L.; Sun, R.; Xu, Y.; Ge, Y.; An, G. Inhibition of DCLK1 down-regulates PD-L1 expression through Hippo pathway in human pancreatic cancer. Life Sci. 2020, 241, 117150. [Google Scholar] [CrossRef]
- Chen, L.; Yang, Q.C.; Li, Y.C.; Yang, L.L.; Liu, J.F.; Li, H.; Xiao, Y.; Bu, L.L.; Zhang, W.F.; Sun, Z.J. Targeting CMTM6 Suppresses Stem Cell-Like Properties and Enhances Antitumor Immunity in Head and Neck Squamous Cell Carcinoma. Cancer Immunol. Res. 2020, 8, 179–191. [Google Scholar] [CrossRef]
- Terzuoli, E.; Bellan, C.; Aversa, S.; Ciccone, V.; Morbidelli, L.; Giachetti, A.; Donnini, S.; Ziche, M. ALDH3A1 Overexpression in Melanoma and Lung Tumors Drives Cancer Stem Cell Expansion, Impairing Immune Surveillance through Enhanced PD-L1 Output. Cancers 2019, 11, 1963. [Google Scholar] [CrossRef]
- Bakir, B.; Chiarella, A.M.; Pitarresi, J.R.; Rustgi, A.K. EMT, MET, Plasticity, and Tumor Metastasis. Trends Cell Biol. 2020, 30, 764–776. [Google Scholar] [CrossRef] [PubMed]
- van Staalduinen, J.; Baker, D.; Ten Dijke, P.; van Dam, H. Epithelial-mesenchymal-transition-inducing transcription factors: New targets for tackling chemoresistance in cancer? Oncogene 2018, 37, 6195–6211. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Durand, S.; Dalle, S.; Caramel, J. EMT-Inducing Transcription Factors, Drivers of Melanoma Phenotype Switching, and Resistance to Treatment. Cancers 2020, 12, 2154. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Z.; Li, Y.; Lu, T.; Hu, G. Silencing Snail Reverses Epithelial-Mesenchymal Transition and Increases Radiosensitivity in Hypopharyngeal Carcinoma. Onco Targets Ther. 2020, 13, 497–511. [Google Scholar] [CrossRef]
- Jiang, F.; Zhou, L.; Wei, C.; Zhao, W.; Yu, D. Slug inhibition increases radiosensitivity of oral squamous cell carcinoma cells by upregulating PUMA. Int. J. Oncol. 2016, 49, 709–719. [Google Scholar] [CrossRef]
- Li, J.; Alvero, A.B.; Nuti, S.; Tedja, R.; Roberts, C.M.; Pitruzzello, M.; Li, Y.; Xiao, Q.; Zhang, S.; Gan, Y.; et al. CBX7 binds the E-box to inhibit TWIST-1 function and inhibit tumorigenicity and metastatic potential. Oncogene 2020, 39, 3965–3979. [Google Scholar] [CrossRef]
- Shahin, S.A.; Wang, R.; Simargi, S.I.; Contreras, A.; Parra Echavarria, L.; Qu, L.; Wen, W.; Dellinger, T.; Unternaehrer, J.; Tamanoi, F.; et al. Hyaluronic acid conjugated nanoparticle delivery of siRNA against TWIST reduces tumor burden and enhances sensitivity to cisplatin in ovarian cancer. Nanomedicine 2018, 14, 1381–1394. [Google Scholar] [CrossRef]
- Wang, M.; He, S.F.; Liu, L.L.; Sun, X.X.; Yang, F.; Ge, Q.; Wong, W.K.; Meng, J.Y. Potential role of ZEB1 as a DNA repair regulator in colorectal cancer cells revealed by cancer-associated promoter profiling. Oncol. Rep. 2017, 38, 1941–1948. [Google Scholar] [CrossRef]
- Hao, Y.; Baker, D.; Ten Dijke, P. TGF-beta-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. Int. J. Mol. Sci. 2019, 20, 2767. [Google Scholar] [CrossRef]
- Yang, T.; Huang, T.; Zhang, D.; Wang, M.; Wu, B.; Shang, Y.; Sattar, S.; Ding, L.; Liu, Y.; Jiang, H.; et al. TGF-beta receptor inhibitor LY2109761 enhances the radiosensitivity of gastric cancer by inactivating the TGF-beta/SMAD4 signaling pathway. Aging 2019, 11, 8892–8910. [Google Scholar] [CrossRef]
- Bayin, N.S.; Ma, L.; Thomas, C.; Baitalmal, R.; Sure, A.; Fansiwala, K.; Bustoros, M.; Golfinos, J.G.; Pacione, D.; Snuderl, M.; et al. Patient-Specific Screening Using High-Grade Glioma Explants to Determine Potential Radiosensitization by a TGF-beta Small Molecule Inhibitor. Neoplasia 2016, 18, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Teeuwssen, M.; Fodde, R. Wnt Signaling in Ovarian Cancer Stemness, EMT, and Therapy Resistance. J. Clin. Med. 2019, 8, 1658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Si, J.; Gan, L.; Guo, M.; Yan, J.; Chen, Y.; Wang, F.; Xie, Y.; Wang, Y.; Zhang, H. Inhibition of Wnt signalling pathway by XAV939 enhances radiosensitivity in human cervical cancer HeLa cells. Artif. Cells Nanomed. Biotechnol. 2020, 48, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yuan, X.; Xu, H.; Yi, M.; Liu, S.; Wen, F. WNT974 Inhibits Proliferation, Induces Apoptosis, and Enhances Chemosensitivity to Doxorubicin in Lymphoma Cells by Inhibiting Wnt/beta-Catenin Signaling. Med. Sci. Monit. 2020, 26, e923799. [Google Scholar] [CrossRef] [PubMed]
- Prakash, V.; Carson, B.B.; Feenstra, J.M.; Dass, R.A.; Sekyrova, P.; Hoshino, A.; Petersen, J.; Guo, Y.; Parks, M.M.; Kurylo, C.M.; et al. Ribosome biogenesis during cell cycle arrest fuels EMT in development and disease. Nat. Commun. 2019, 10, 2110. [Google Scholar] [CrossRef]
- Dermit, M.; Dodel, M.; Lee, F.C.Y.; Azman, M.S.; Schwenzer, H.; Jones, J.L.; Blagden, S.P.; Ule, J.; Mardakheh, F.K. Subcellular mRNA Localization Regulates Ribosome Biogenesis in Migrating Cells. Dev. Cell 2020, 55, 298–313.e210. [Google Scholar] [CrossRef]
- Pecoraro, A.; Carotenuto, P.; Russo, G.; Russo, A. Ribosomal protein uL3 targets E2F1 and Cyclin D1 in cancer cell response to nucleolar stress. Sci. Rep. 2019, 9, 15431. [Google Scholar] [CrossRef]
- Fan, H.; Li, J.; Jia, Y.; Wu, J.; Yuan, L.; Li, M.; Wei, J.; Xu, B. Silencing of Ribosomal Protein L34 (RPL34) Inhibits the Proliferation and Invasion of Esophageal Cancer Cells. Oncol. Res. 2017, 25, 1061–1068. [Google Scholar] [CrossRef]
- Mattijssen, S.; Arimbasseri, A.G.; Iben, J.R.; Gaidamakov, S.; Lee, J.; Hafner, M.; Maraia, R.J. LARP4 mRNA codon-tRNA match contributes to LARP4 activity for ribosomal protein mRNA poly(A) tail length protection. eLife 2017, 6, e28889. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).