Abstract
Wetlands are of a considerable environmental value as they provide food and habitat for plants and animals. Several important chemical transformations take place in wetland media, including the conversion of inorganic mercury (Hg) to monomethylmercury (MeHg), a toxic compound with a strong tendency for bioconcentration. Considering the fact that wetlands are hotspots for Hg methylation, we investigated, for the first time, Hg methylation and demethylation rates in an old growth cypress wetland at Sky Lake in the Mississippi Delta. The Sky Lake ecosystem undergoes large-scale water level fluctuations causing alternating periods of oxic and anoxic conditions in the sediment. These oscillating redox conditions, in turn, can influence the transformation, speciation, and bioavailability of Hg. In the present study, sediment cores from the wetland and Sky Lake itself were spiked with enriched stable isotope tracers of inorganic Hg and MeHg and allowed to incubate (in-situ) before freezing, sectioning, and analysis. Methylation rates (day−1) ranged from 0.012 ± 0.003 to 0.054 ± 0.019, with the lowest rate in the winter and the highest in the summer. Demethylation rates were about two orders of magnitude higher, and also greater in the warmer seasons (e.g., 1.84 ± 0.78 and 4.63 ± 0.51 for wetland sediment in the winter and summer, respectively). Methylation rates were generally higher in the open water sediment compared to wetland sediment, with the latter shaded and cooler. Both methylation (r = 0.76, p = 0.034) and demethylation (0.97, p = 0.016) rates (day−1) were positively correlated with temperature, but not with most other water quality parameters. MeHg concentration in the water was correlated with pH (r = 0.80, p < 0.05), but methylation rates were only marginally correlated (r = 0.71). Environmental factors driving microbial production of MeHg in the system include warm temperatures, high levels of labile natural organic matter, and to a lesser extent the relatively low pH and the residence time of the water. This study also provides baseline data that can be used to quantify the impacts of modifying the natural flow of water to the system on Hg methylation and demethylation rates.
1. Introduction
Methylmercury (MeHg) is a neurotoxin species produced primarily by sulfate reducing bacteria in aquatic sediments that readily biomagnifies up freshwater and marine food chains [1]. Consumption of fish containing high levels of MeHg can lead to severe adverse health effects in both humans and wildlife [2]. To understand the factors controlling the environmental concentration of MeHg in natural water bodies, there is a need to measure both inorganic (Hg+2) and organic (MeHg) forms, as well as the relative rates of methylation and demethylation. Thus, enriched stable isotopes have been used to simultaneously study the methylation and demethylation reactions [3].
Mercury entering naturally occurring aquatic systems stems from both natural and anthropogenic sources, with the latter primarily from combustion of fossil fuels, as well as contaminated from legacy Hg mines, soda-PVC plants, artisanal gold mining, etc. [4]. Inorganic Hg can be transformed to MeHg by abiotic and biotic methylation processes [4,5,6]. The majority of Hg methylation is carried out by sulfate-reducing bacteria (SRB), with a minor contribution from Fe-reducing bacteria. These bacteria use vitamin B-12 for transferring a CH3 group via radical reactions [7].
Wetlands are an important resource because they provide many positive environmental benefits, including habitat for wildlife, buffering for flood-prone areas, and a natural filtering effect of some water impurities [8,9]. Wetlands also play a critical role in the transformation of Hg within landscapes and are considered hotspots for Hg methylation, in part because SRB thrive in wetland sediments [10]. Wetlands have been shown to be net sources of MeHg to ecosystems and may, in part, explain the high concentrations of Hg found in fish in remote and near-pristine regions [11].
MeHg diffuses out of the microorganism and enters the water column where it readily accumulates in plankton and biomagnifies in the food web where it can have ecological and human health impacts. Environmental factors that seem to promote methylation, all of which can be found in wetlands, include warm temperatures, low pH, the presence of humic materials, and long water residence time [12,13]. Previous studies have shown that humic materials can accumulate heavy metals, including Hg, and have been shown to increase natural bacterial assemblages, serving as a carbon and energy source [14,15].
As wetlands are documented to exhibit elevated MeHg production compared to other aquatic habitats, it is of paramount importance to assess Hg methylation in different types of wetlands and within important watersheds. Considering the significance, we set out to: (1) measure methylation/demethylation rates seasonally in an ancient cypress wetland and adjacent open water areas in Sky Lake, (2) correlate methylation/demethylation rates with ancillary data (pH, dissolved oxygen, temperature, conductivity, etc.), and (3) determine the factors driving Hg methylation in the system. Finally, this study also provides baseline data that can be used to quantify the impacts of altering the natural flow of water to the system on Hg methylation and demethylation rates. Such changes may occur with climate change or more abruptly with changes in land use.
2. Materials and Methods
2.1. Site Descriptions
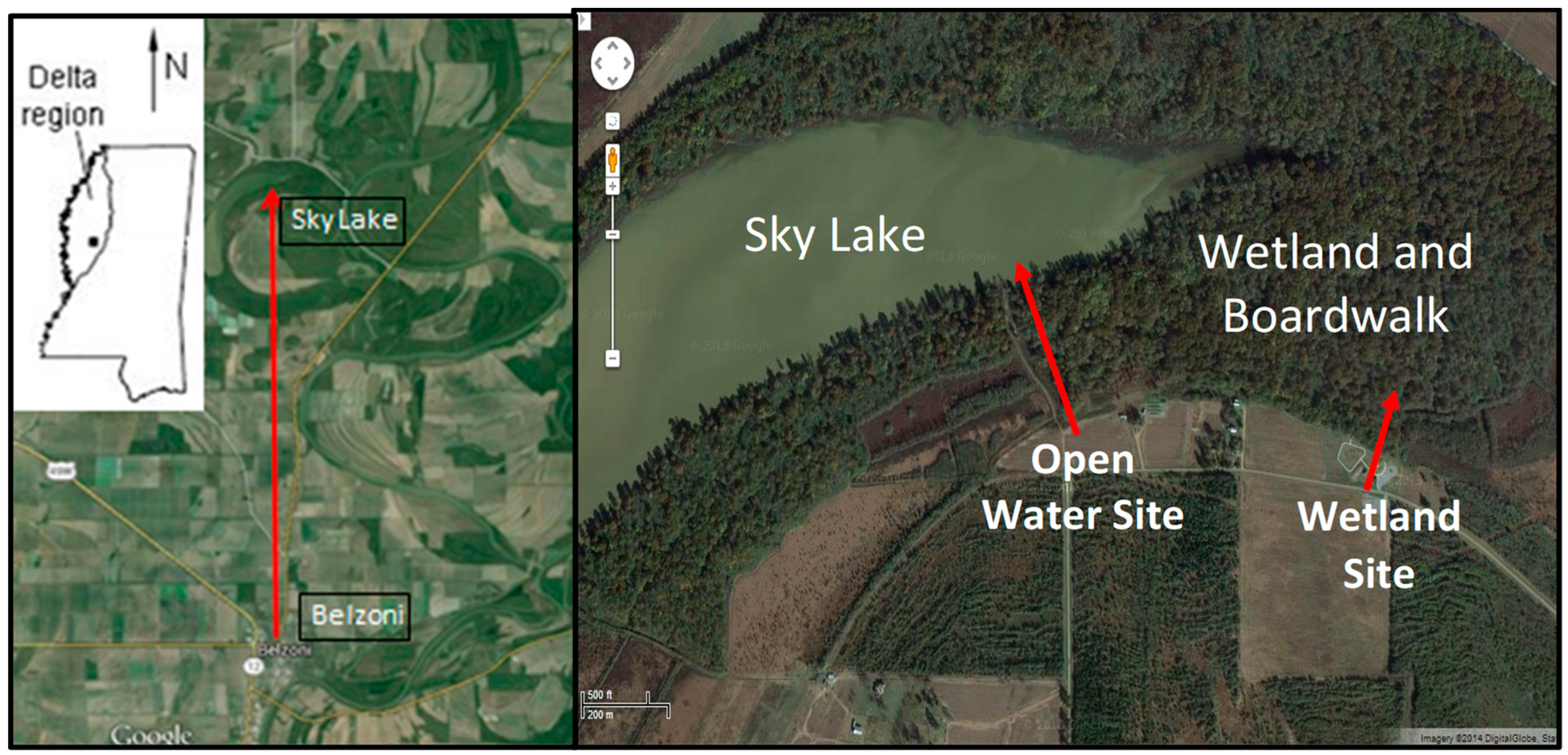
Sky Lake is an oxbow lake located in the Mississippi Delta between the Yazoo and the Big Sunflower Rivers (Figure 1). A wetland associated with the lake contains one of the few old growth Cypress forests remaining in the world, 10 km from the town of Belzoni (Figure 2). The site has an elevated boardwalk that stands atop a portion of the wetland. The cypress wetland at Sky Lake undergoes large-scale water level fluctuations that are thought to affect the redox conditions in the sediment resulting in alternating periods of oxic and anoxic conditions. These oscillations in redox conditions may influence the methylation and demethylation rates, which affect speciation and bioavailability of Hg in the system. We were granted permission to access the site and setup instrumentation.
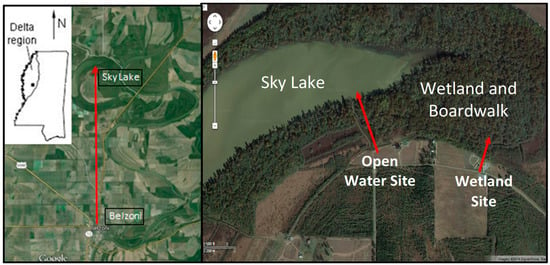
Figure 1.
Sky Lake study area located in the Mississippi Delta. The satellite images depict the location of the open water and wetland study sites.

Figure 2.
Sky Lake Wildlife Management Area. The large cypress tree in the foreground is several hundred years old. The arrow indicates a recent water level, demonstrating seasonal water level fluctuations of several meters in the wetland.
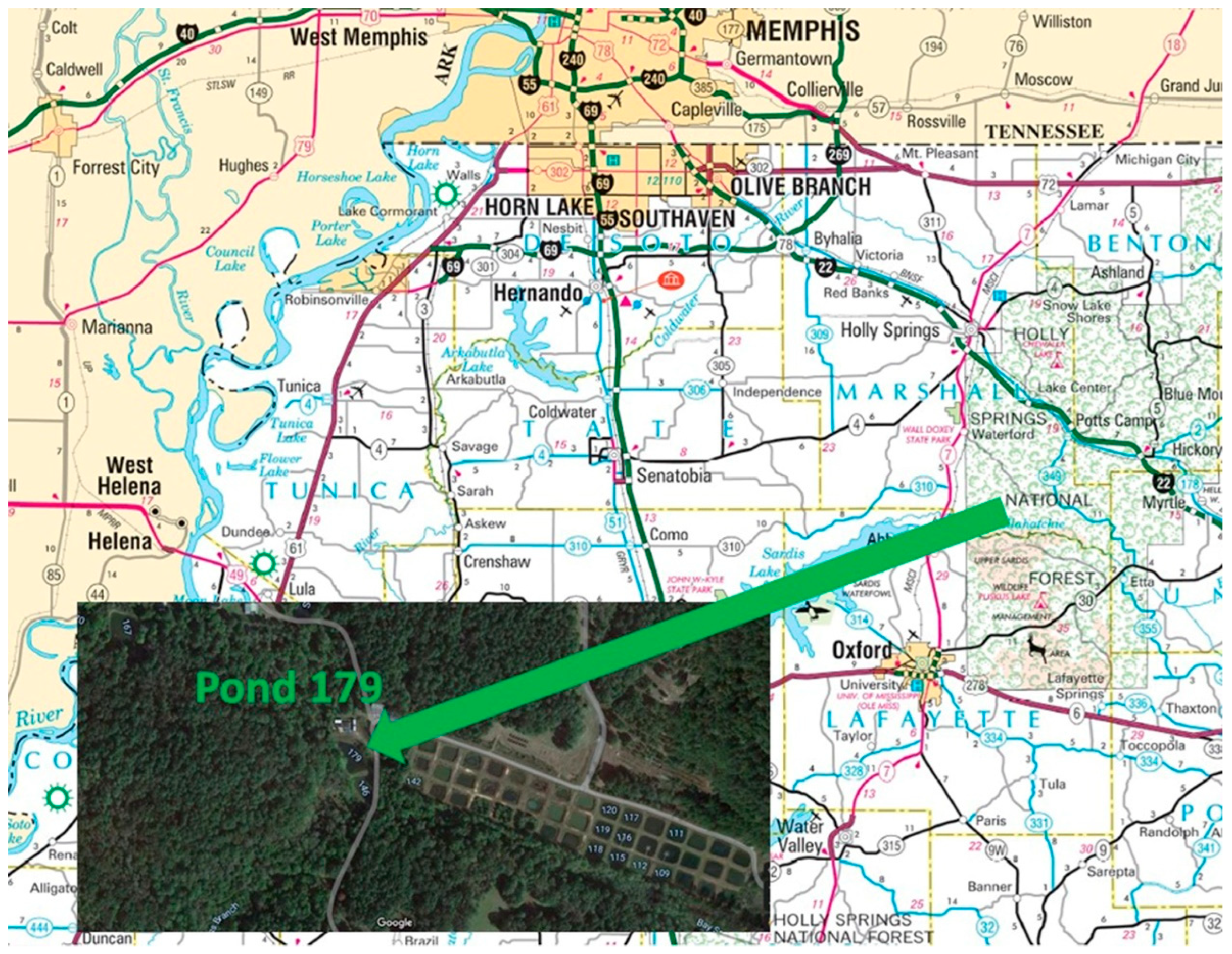
The UM Field Station is located 11 miles northeast of the UM Oxford campus off County Road 202. The Field Station is located on 740 acres of land and consists of 200 experimental ponds ranging from 0.1 to 2 acres. Ponds are typically about 1 m in depth. The UM Field Station site was chosen as a control site due to the water level stability and was conducted at pond 179 (Figure 3). A prior study with sediments from this site observed the effect of activated carbon, biochar, and humus/manure on Hg methylation rates [14]. All maps were provided by Google Map image except Figure 3 which comes from the Mississippi Department of Transportation.
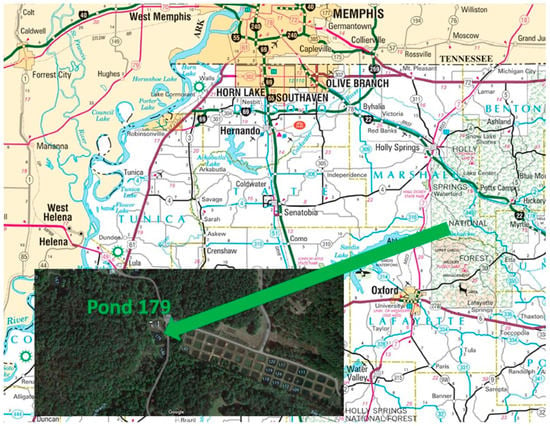
Figure 3.
University of Mississippi Field Station—Pond 179.
2.2. Enriched Stable Isotopes of Hg and Reference Materials
Enriched isotopes of 199HgO, 200HgO, and 201HgO were purchased from Oak Ridge National Laboratories (ORNL) (Oak Ridge, Tennessee, TN, USA) and dissolved in 10% optima grade nitric acid. MeHg containing enriched isotopes was subsequently synthesized using the methylcobalamine method [16]. All other reagents used were reagent grade. Ultrapure water (>18 ΜΩ cm−1) was obtained from a Barnstead Nanopure Diamond system. For reference materials, we used DORM-3 (dogfish muscle) for total Hg, and CC-580 (Estuarine Sediment) for MeHg. DORM-3 was obtained from the National Research Council of Canada (Ottawa), and CC-580 was obtained from the European Commission Institute for Reference Materials and Measurements (IRMM) in Belgium.
2.3. Sediment Collection and Processing
Two sediment cores were collected seasonally from both wetland (W) and open water (OW) areas at Sky Lake during the years 2013–2014. MeHg production is related to temperature and microbial activity, and has been shown to vary seasonally [15,17]. Wetland sediments are typically anoxic or become anoxic within the first cm or less, and thus Hg methylation is generally highest in the top layers of the sediment [15]. We chose to measure Hg methylation rates in the top 6 cm of the sediment, but on occasion even deeper. Sediment cores were collected from Sky Lake and the Biological Field Station using plastic core liners (PVC 2″ × 36″) purchased from AMS, Inc. Holes were drilled every 1 cm down the length of each tube to allow enriched isotopic tracers (Me199Hg+ and 200Hg2+) to be injected throughout the sediment core for the tracer study (Figure 4). The holes were filled with clear waterproof silicone.

Figure 4.
Injecting Hg tracer isotopes into a sediment core.
Two (duplicate) sediment cores were taken at each sample site. Stable isotope spike solutions of 200Hg2+ and 199CH3Hg+ were prepared at concentrations ranging from ~10–100% of ambient concentrations by diluting the standard with bottom water from the site. The mixture was left to equilibrate for an hour prior to usage to attain natural conditions/speciation [18]. Approximately 0.2 mL of spike solution was injected into each 1 cm increment down to approximately 20 cm (Figure 4). After spiking, the sediment cores were placed back into the wetland and open water sites to incubate for 2 h before being transported in a cooler to the laboratory. At the lab, a modified wood extruder was used to extrude the cores onto a plastic cutting board. A stainless-steel knife was used to section the top 6 cm of the core and to divide the top 6 cm into 0–2, 2–4, and 4–6 cm sections. The samples were taken wearing gloves and stored overnight in a −80 °C freezer and subsequently lyophilized for 5 days.
2.4. Extracting and Isolating MeHg from the Sediment
Before distillation, all equipment used was acid-washed in a 15% HCl bath, rinsed with Ultrapure water, and dried under a laminar flow hood. The acetate buffer and 1% sodium tetraethyl-borate were prepared according to EPA Method 1630 [19]. The sediment extraction and distillation procedure is based on Method 1630 and can be found in the Supplementary Material.
2.5. Determination of MeHg Species in Sediments by GC-ICP-MS
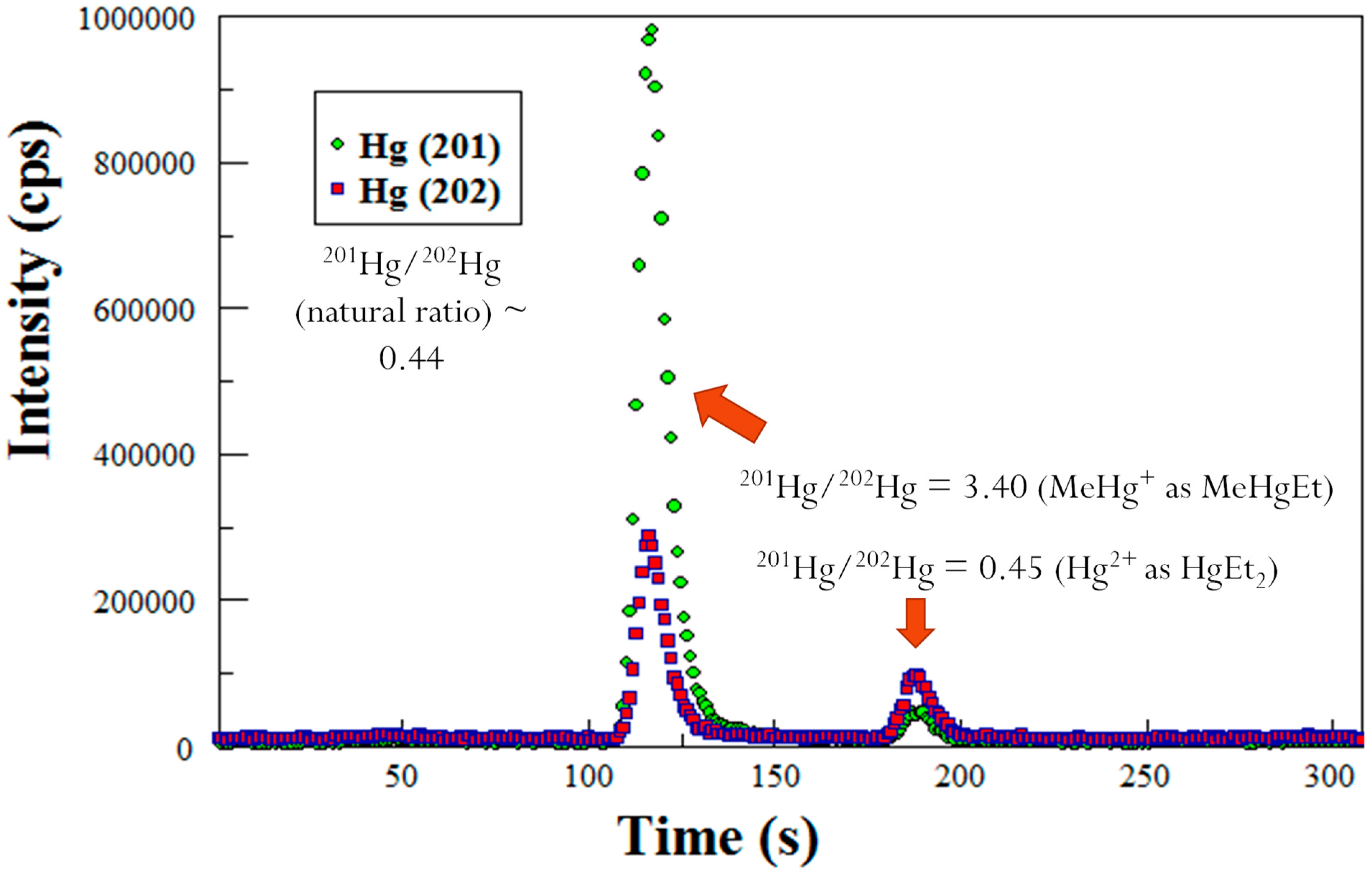
To determine the amount of MeHg in the sediments, GC-ICP-MS was utilized as described by Lambertsson [20]. Briefly, a GC column from a purge and trap Tekran 2700 MeHg analyzer was used for the separation of the different species of Hg. The effluent coming from the atomic fluorescence cell of the Tekran 2700 is coupled via a Teflon line to the sample injector to the ICP-MS. The isotopes of Hg were detected by the electron multiplier. Prior to analysis, the solution was ethylated such that all Hg compounds became volatilized although Hg0 is volatile by itself. The mercuric ion becomes diethyl Hg, and MeHg becomes MeEtHg. The Hg species were separated out by the GC that is contained in the Tekran 2700. The Hg species flowed through the atomic fluorescence detector, and were introduced into the plasma of the ICP-MS which is a technique, that provides excellent separation. A typical GC-ICP-MS chromatogram is shown in Figure 5. Each data point on the chromatogram is a mass spectrum that was generated from the ICP-MS. The chromatogram shows that when Me201Hg is spiked into a sample mixture, the resulting ratios of 201/202 Hg are changed to 3.40. The naturally occurring ratio of 201/202 is 0.44 [21].
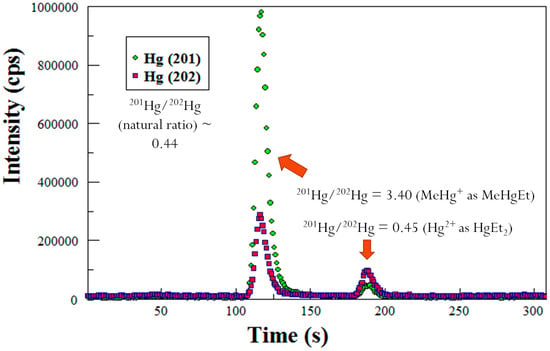
Figure 5.
GC-ICP-MS chromatogram showing isotope ratios for Hg species: Hg+2 with natural abundance (peak ~ 185 s) and an enriched isotope 201MeHg spike (peak ~ 115 s).
2.6. Determination of Total-Hg and Hg Isotopes in Sediments by DMA-ICP-MS
DMA coupled to ICP-MS offers several advantages over the DMA alone for the determination of mercury in methods that require distinguishing the individual isotopes [22]. These include increased sensitivity, lower detection limits, decreased potential for sample contamination, and applicability to Hg stable isotope tracer studies. As mercury isotopes are required for the methylation and demethylation calculations, total Hg concentrations and Hg isotopes in sediment were determined by DMA-ICP-MS as described in Bussan et al. [22]. Briefly, sample boats were cleaned prior to use by running the boats as blanks in the DMA. Concentrations were calculated using the single spike isotope dilution equation using 201Hg2+ and 201CH3Hg+1 (Equation (1)) [23]. Ambient MeHg concentrations in the sediment were also determined by GC-ICP-MS using the same equation
The abbreviations of the constants are as follows: Cn: concentration of sample; Cspike: concentration of spike; Ms: weight of the spike; Mn: weight of the sample; WN: atomic weight of the element in the sample; WS: atomic weight of the element in the spike AbAs: abundance of A in the spike; AbBs: abundance of B in the spike; AbAN: abundance of A in the sample; AbBN: abundance of B in the sample; and Rm: ratio of the mixture.
2.7. Determination of Hg Methylation and MeHg Demethylation Rates
Hg methylation and demethylation rates were determined following Hintelmann et al. [18,23]. Briefly, to determine the amount of methylated and demethylated Hg at least three isotopes of Hg need to be monitored. One isotope represents the newly produced MeHg from the added inorganic Hg tracer (200Hg in this study); another represents the demethylation of the added MeHg tracer (Me199Hg); and the third represents changes in the MeHg concentrations derived from the Hg originally present in the sample (e.g., 202Hg) [1].
The net production of methylation is represented by Equation (2).
where [CH3XHg+] = concentration of CH3XHg+ newly generated from the XHg2+ tracer (in ng/g), km = specific methylation rate constant (in d−1), [XHg2+] = concentration of added XHg2+ (in ng/g), kd = specific demethylation rate constant (in d−1), and t = incubation time (in days). By having a large excess of XHg2+, we can solve for the specific methylation constant assuming a pseudo-first-order reaction; Equation (2) becomes Equation (3):
Likewise, when CH3199Hg+ is spiked to the sediment, the 200Hg2+ from the demethylation is negligible, and Equation (3) reduces down to:
Integration of Equation (4) yields Equation (5), which can be used to determine the demethylation rate constant (kd).
2.8. Instrumental Parameters
The DMA-ICP-MS instrumental parameters used in this study are given in Table 1.

Table 1.
DMA-ICP-MS instrumental parameters.
2.9. Water Quality Parameters
Water quality parameters were measured with a portable YSI Professional Plus meter. Dissolved oxygen (DO) (mg L−1), pH, total dissolved solids (TDS) (mg L−1) using the default instrument conversion constant, oxidation reduction potential (ORP) (mV), and temperature (°C) were measured (Table 2). Total suspended solids (TSS) (mg L−1) were measured using a portable suspended solids analyzer (InsitelG, model 3150). Sulfide (S2-) (mV) was measured with an Orion silver/sulfide ion selective electrode using Pb (ClO4)2 as the titrant. Loss on ignition (LOI) was calculated using a relationship established in our previous work between LOI and total carbon and which is specific for the sampling sites [24].
Table 2.
Water quality parameters for Sky Lake open water and wetland areas during sampling.
2.10. Statistical Analysis
Statistical evaluation was performed using STAT PLUS 2009 software. Data were evaluated for correlations using the Pearson Linear correlation test. In addition, p-values or observed significance values were calculated for the different parameters measured. A p-value < 0.05 is considered to be of statistical significance. Univariate analysis of variance (ANOVA) with post hoc Tukey HSD test was also used as a statistical test to assess the differences among groups based on a set of dependent variables.
3. Results and Discussion
A summary of Hg methylation and demethylation rates is given in Table 3, while total Hg, MeHg, and organic matter (LOI) for the sediment and MeHg data for the water is provided in Table 4. Ancillary water quality data, including temperature, dissolved oxygen, total dissolved solids, oxidation reduction potential, pH, total suspended solids, conductivity, and sulfide concentration is given in Table 2. These results are discussed in the subsections below.
Table 3.
Sediment Hg methylation (Km) and MeHg demethylation (Kd) rates for sediment in Sky Lake (open water and wetland areas) and the University of Mississippi’s Biological Field Station (n = 3; ± 1 SD) n, bulk samples of the same sediment.
Table 4.
Concentrations of bulk samples of the same sediment for total Hg and MeHg in sediment and water from Sky Lake wetland and open water areas, as well as loss on ignition (LOI), an estimate of organic matter in the sediment.
Sediment total Hg on a dry weight basis ranged from 12.5 ng g−1 to 189.0 ng g−1 at the open water site and from 20.0 ng g−1 to 121.6 ng g−1 at the wetland water site. The large variation of total mercury may be attributed to the rate of conversion of inorganic mercury being converted to methyl mercury by microscopic organisms in sediments and water. Sediment MeHg ranged from 0.36 ng g−1 to 1.34 ng g−1 at the open water site and ranged from 0.43 ng g−1 to 1.43 ng g−1 at the wetland water site. These results are similar to Hugget et al. who reported sediment Hg levels ranging from 27 ng g−1 to 168 ng g−1 in three Mississippi reservoirs (Enid, Sardis and Grenada Lakes) [25]. There are no major source points in either of these two different watersheds, so the likely source of the Hg is atmospheric deposition from both natural and distant anthropogenic sources, along with some leaching of Hg from the mineral environment.
Hall et al. (2008) sampled surface water and pore water in a variety of wetlands in southern Louisiana and the Gulf of Mexico region between August of 2003 and May 2005. Their study revealed that total Hg and MeHg concentrations in surface waters were appreciably greater in freshwater and brackish wetlands, as these sites are expected to support microbial methylation of Hg, while the values were lower in marine wetlands and brackish and marine open water systems. In particular, for surface waters, they reported MeHg levels as high as 2.15 ng L−1 and total Hg levels as high as 5.58 ng L−1 [26]. Both values are higher than the ones reported in the present study. In addition, the authors determined that MeHg concentrations were greatest in wetland water compared to non-vegetated regions of the lake and river mainstems. However, we found slightly lower concentrations for MeHg in water at the wetland site (5.93 ± 0.87 pg g−1) compared to the open water site (7.41 ± 0.83 pg g−1), although we have too few data to make a robust statistical comparison.
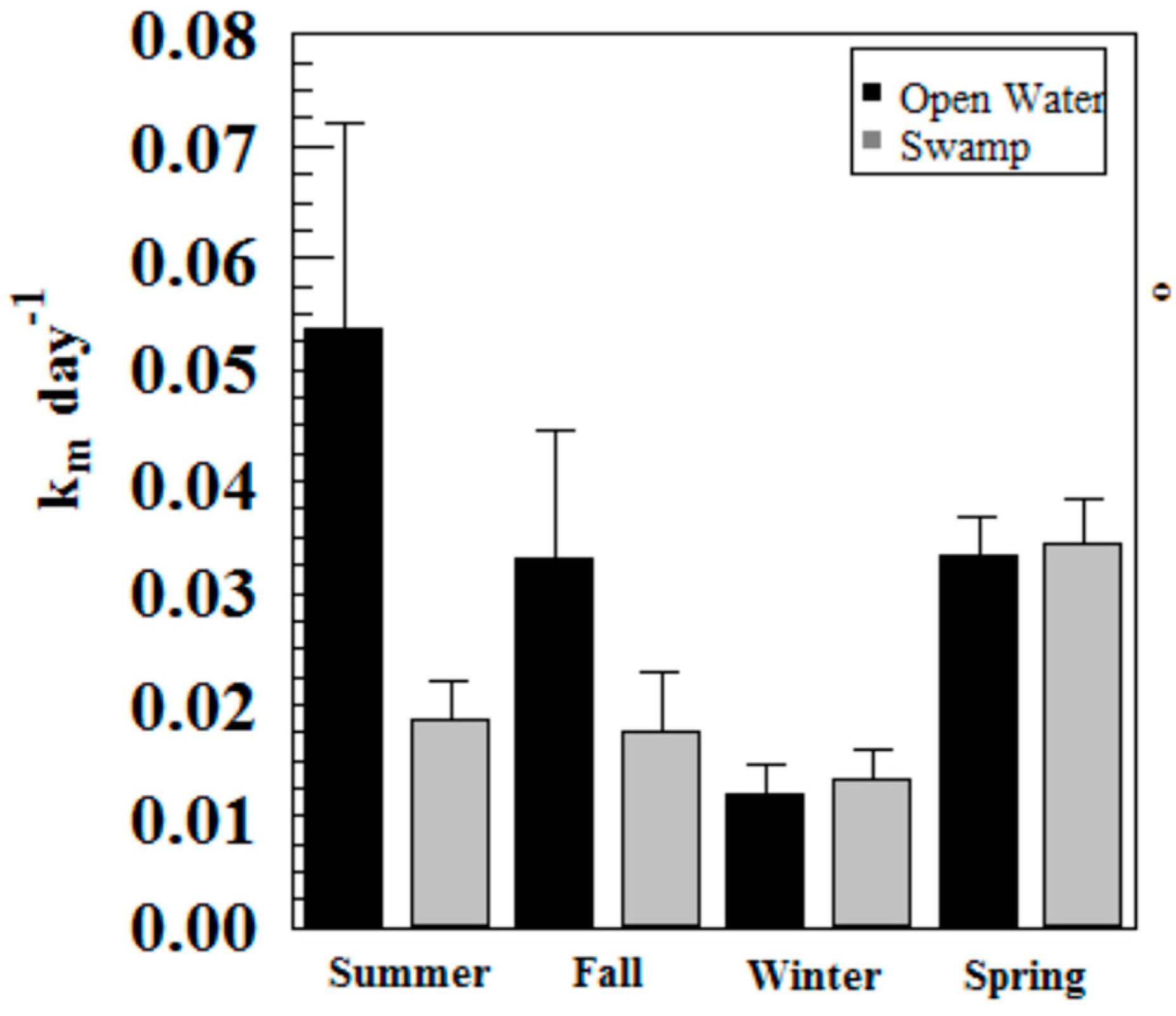
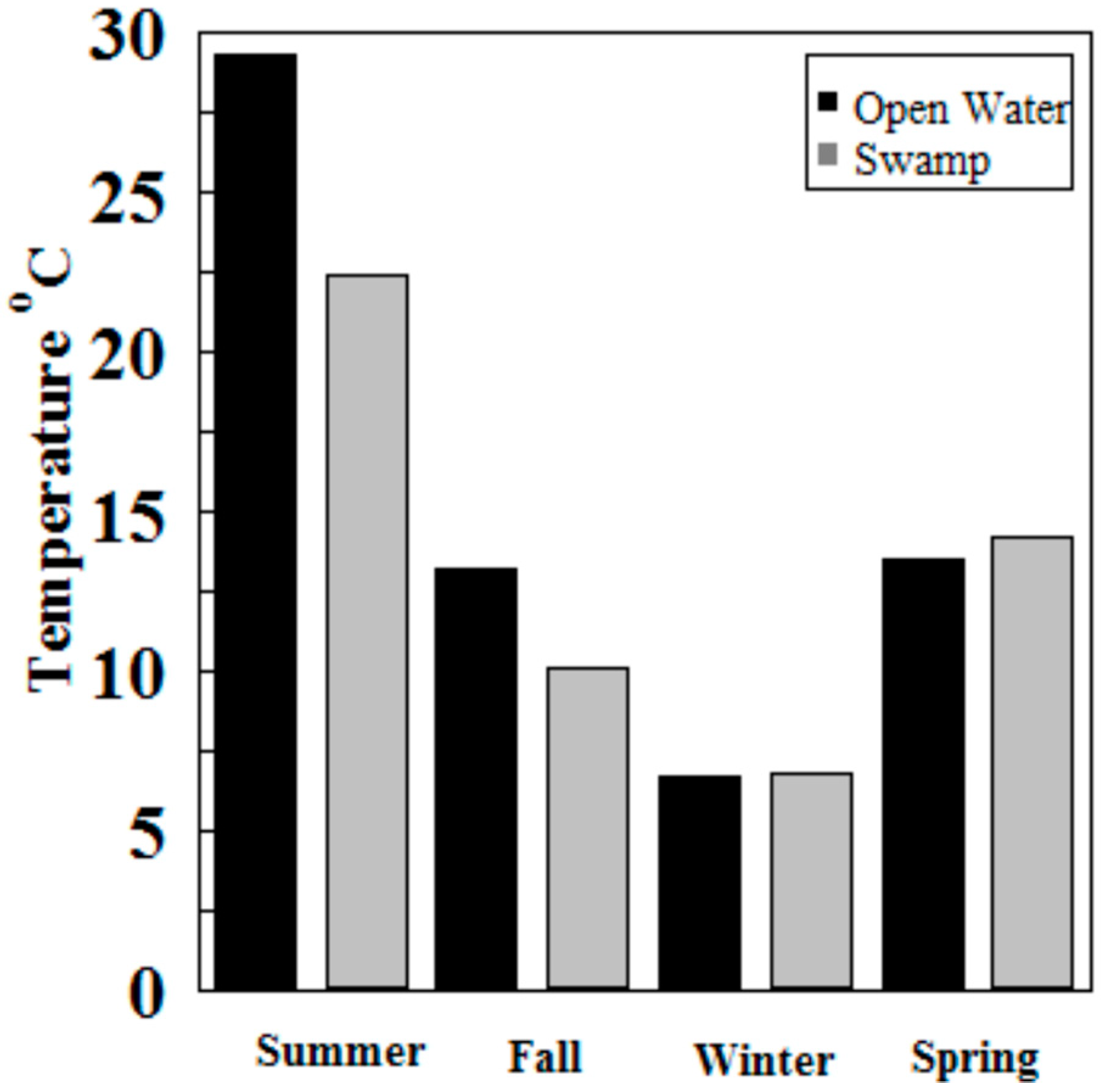
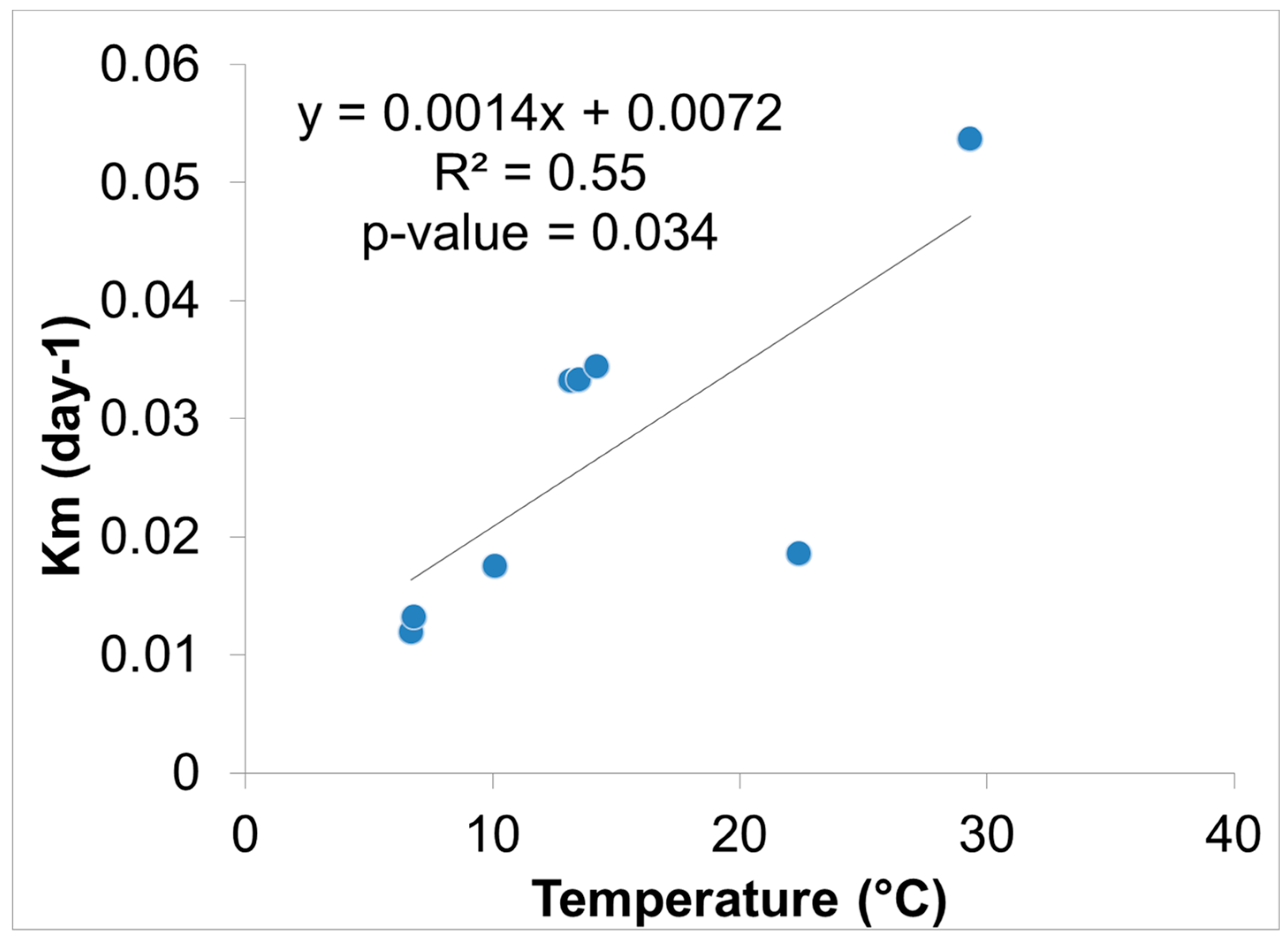
Hg methylation rates were lowest in the winter and highest in the summer for the open water site (Figure 6). This is likely related to higher microbial activity with higher temperatures [27]. Indeed, we found Hg methylation rates and temperature were correlated (r = 0.76, p = 0.034) despite the relatively low Km in the summer for the wetland (Figure 7 and Figure 8). As reported by others, temperature plays a major role for seasonal differences in Hg methylation rates [28,29]. In our study, the difference between methylation rates for the open water and wetland sites were likely exacerbated by the dense foliage and tree canopy of the forested wetland. The water temperature of the shaded wetland was substantially cooler than the unshaded open water site. In the winter, when foliage coverage is not a significant factor in the wetland, the methylation rates were found to be virtually identical. The same was found to be true in the early spring when wetland foliage had not yet peaked.
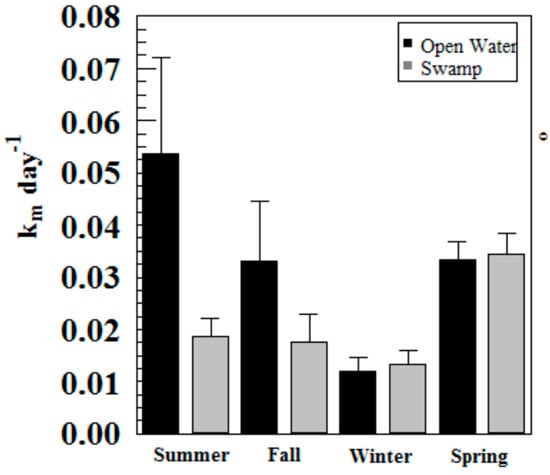
Figure 6.
Hg methylation rates in Sky Lake by season (n = 3 per sample).
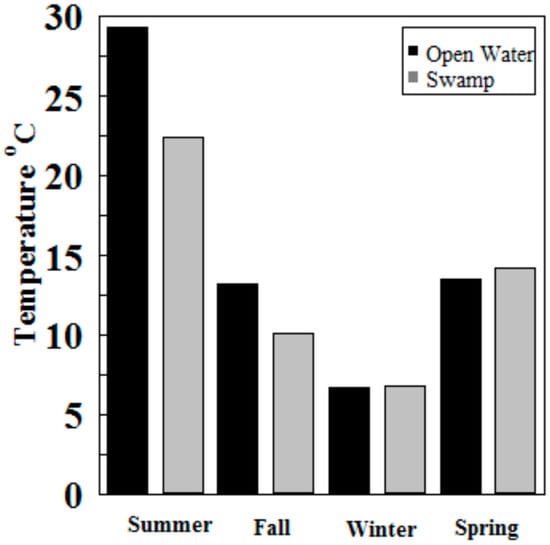
Figure 7.
Water temperatures at Sky Lake open water and wetland sites during sampling (n = 3 per sample).
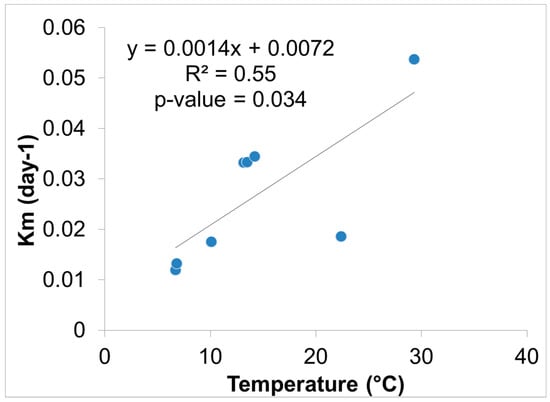
Figure 8.
Hg methylation rate vs. ambient water temperature near the sediment surface (n = 8).
Methylation rates were also generally higher in the open water sites compared to the wetland sites, except in winter and spring where they were nearly identical.
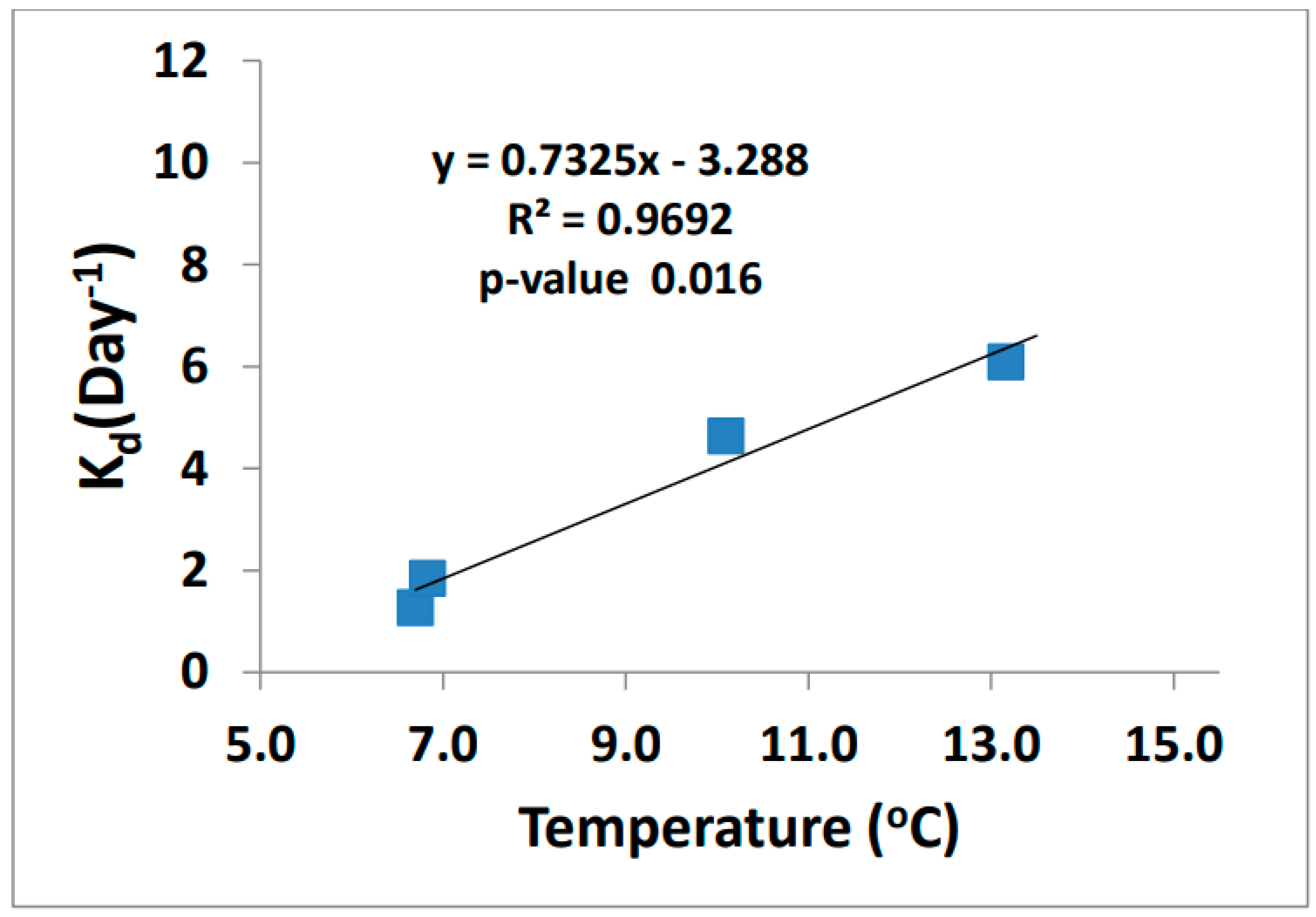
The demethylation rates increased as temperature increased (Figure 9). Demethylation can occur through both oxidative and reductive processes. The end products are what define the type of process that happens, either CO2 (oxidative demethylation) or CH4 (reductive demethylation). Marvin-Dipasquale et al. found that reductive demethylation occurs in highly contaminated sediments as a detoxification process and is performed by bacteria containing the mer-operon [30,31,32,33]. The microbes in the biotic reaction can cleave the methyl group from MeHg resulting in CH4 and Hg2+ products using the enzyme organomercurial lyase [34].
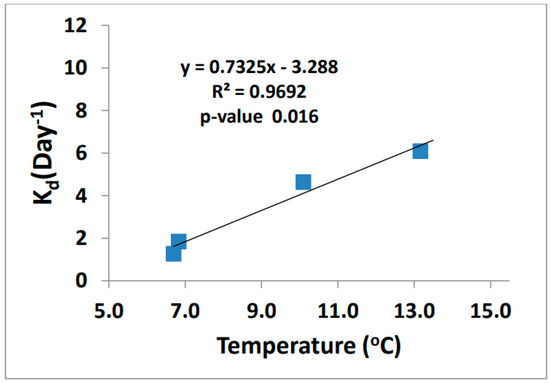
Figure 9.
MeHg demethylation rate vs. ambient temperature (n = 4).
A relationship between Hg methylation rates and ancillary data using a Pearson’s correlation matrix on the wetland and open water sites revealed the relationship between Km, Hg, MeHg, and ancillary data (Table 5). As noted, temperature was highest during the summer months (29.3 °C, open water and 22.4 °C, wetland water) and correlated positively with Km rate values (r = 0.76, p = 0.034). There was also a positive correlation between pH vs. MeHg (r = 0.80, p = 0.03), and a negative correlation between pH vs. Km rate values (r = −0.36, p = 0.65). Xun et al. similarly observed that a decrease in pH would result in specific rates of Hg methylation increasing [35]. The authors hypothesized that an increase in methylation at a lower pH could be due to higher uptake of Hg+2 by the methylating bacteria. The process of Hg uptake differs between species of microorganism, meaning that one species incorporates Hg primarily by passive diffusion, while another species by facilitated diffusion [35,36].
Table 5.
Pearson’s correlation matrix for water quality parameters measured in seasonal trends for Sky Lake, Mississippi, open water, and wetland sites. Correlations with p-values < 0.05 are italicized.
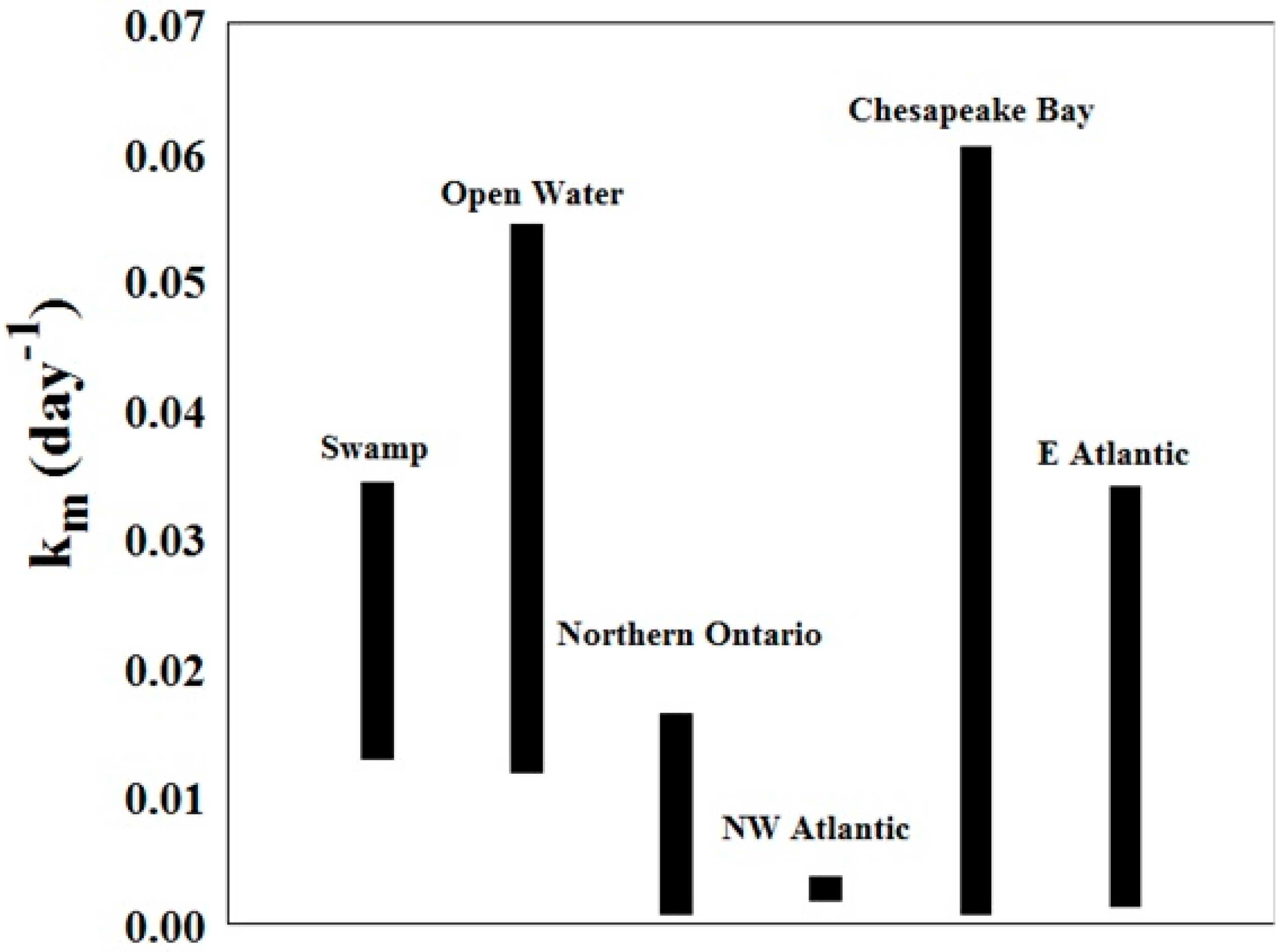
Comparison of Hg Methylation Rates with Literature Values
The values for Km (day−1) for this study generally overlap with the range found in previous studies at other locations [1,36,37]. They also tend to be on the higher side, with mean rates higher than those found in northern Ontario, in the northwest Atlantic Ocean, and in the east Atlantic Ocean (Figure 10). As explained previously, temperature plays an important role in the methylation process. The samples from this study were obtained in the Southeast US, where the climate is warmer than the other study sites.
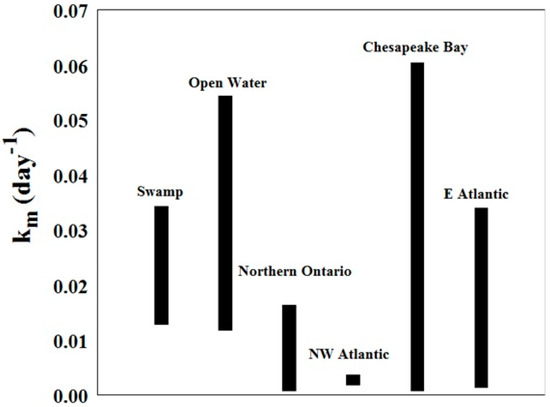
Figure 10.
Methylation rates from this study (bars labelled swamp and open water) compared to literature values at sites along the Atlantic Coast and in northern Ontario [1,36,37].
4. Conclusions
The processes that drive Hg methylation and MeHg demethylation in wetland sediments are of paramount importance as wetlands, due to their unique biochemical conditions, are known to have increased MeHg production compared to other aquatic environments. The study of these conditions and their influence on net MeHg production is important as MeHg can have profound effects on ecosystems. Here, we measured, for the first time, Hg methylation potentials in a unique and ancient cypress wetland with the aim of better understanding the complex factors affecting in situ MeHg production. As a result, a number of important data was collected, analyzed, and presented in this report. These include the amounts of MeHg in wetland sediments measured by GC-ICP-MS, which ranged from 0.36 to 1.43 ng g−1, and the amounts of total Hg in sediments by DMA-ICP-MS, which ranged from 12.5 to 189.0 ng g−1. In addition, Hg methylation and demethylation rates were assessed and found to vary seasonally with the highest rates in the summer (0.034 day−1) and the lowest in the winter (0.013 day−1). To help determine the factors driving Hg methylation in the system, correlations between methylation and demethylation rates with ancillary data were evaluated. Most critical was temperature, as the temperature increased the Hg methylation rate increased. For the open waters of Sky Lake, Hg methylation rates ranged from 0.012 to 0.054 day−1, slightly higher than the adjacent (shaded) wetland areas, which suggests that shallow waters in lakes should not be overlooked as potential sources of MeHg. Finally, as Hg methylation data are scarce in the literature, this work can serve as a baseline for future studies at this site, as well as to compare with other sites the Southeastern USA.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/molecules27154911/s1, Detailed Description of the method of extracting and isolating MeHg from sediments.
Author Contributions
Conceptualization, D.D.B. and J.V.C.; Methodology, D.D.B. and J.V.C.; Resources, D.D.B., J.V.C. and C.D.; data curation, D.D.B., J.V.C. and C.D.; writing-original draft preparation, D.D.B. and J.V.C.; writing-review and editing, D.D.B., J.V.C. and C.D.; visualization, D.D.B., J.V.C. and C.D.; supervision, D.D.B. and J.V.C.; project administration, D.D.B. and J.V.C.; funding acquisition, J.V.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available upon request.
Acknowledgments
We would like to express sincere thanks to the anonymous reviewers for their constructive comments, which significantly improved the quality of the paper.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Not available.
References
- Martín-Doimeadios, R.R.; Tessier, E.; Amouroux, D.; Guyoneaud, R.; Duran, R.; Caumette, P.; Donard, O.F.X. Mercury Methylation/Demethylation and Volatilization Pathways in Estuarine Sediment Slurries Using Species-Specific Enriched Stable Isotopes. Mar. Chem. 2004, 90, 107–123. [Google Scholar] [CrossRef]
- Kudo, A.; Fujikawa, Y.; Miyahara, S.; Zheng, J.; Takigami, H.; Sugahara, M.; Muramatsu, T. Lessons from Minamata Mercury Pollution, Japan—After a Continuous 22 Years of Observation. Water Sci. Technol. 1998, 38, 187–193. [Google Scholar] [CrossRef]
- Pirrone, N.; Cinnirella, S.; Feng, X.; Finkelman, R.B.; Friedli, H.R.; Leaner, J.; Mason, R.; Mukherjee, A.B.; Stracher, G.B.; Streets, D.G. Global Mercury Emissions to the Atmosphere from Anthropogenic and Natural Sources. Atmos. Chem. Phys. 2010, 10, 5951–5964. [Google Scholar] [CrossRef] [Green Version]
- Hammerschmidt, C.R.; Fitzgerald, W.F. Bioaccumulation and Trophic Transfer of Methylmercury in Long Island Sound. Arch. Environ. Contam. Toxicol. 2006, 51, 416–424. [Google Scholar] [CrossRef] [Green Version]
- Mason, R.P.; Lawson, N.M.; Lawrence, A.L.; Leaner, J.J.; Lee, J.G.; Sheu, G.-R. Mercury in the Chesapeake Bay. Mar. Chem. 1999, 65, 77–96. [Google Scholar] [CrossRef]
- Han, S.; Narasingarao, P.; Obraztsova, A.; Gieskes, J.; Hartmann, A.C.; Tebo, B.M.; Allen, E.E.; Deheyn, D.D. Mercury Speciation in Marine Sediments under Sulfate-Limited Conditions. Environ. Sci. Technol. 2010, 44, 3752–3757. [Google Scholar] [CrossRef]
- Bussan, D.D.; Ochs, C.A.; Jackson, C.R.; Anumol, T.; Snyder, S.A.; Cizdziel, J.V. Concentrations of Select Dissolved Trace Elements and Anthropogenic Organic Compounds in the Mississippi River and Major Tributaries during the Summer of 2012 and 2013. Environ. Monit. Assess. 2017, 189, 73. [Google Scholar] [CrossRef]
- Bussan, D.; Harris, A.; Douvris, C. Monitoring of Selected Trace Elements in Sediments of Heavily Industrialized Areas in Calcasieu Parish, Louisiana, United States by Inductively Coupled Plasma-Optical Emission Spectroscopy (ICP-OES). Microchem. J. 2019, 144, 51–55. [Google Scholar] [CrossRef]
- Snaychuk, L.; Vaughan, T.; Ullery, Z.; Douvris, C.; Bussan, D.D. Inductively Coupled Plasma Optical Emission Spectroscopy (ICP-OES) Analysis of Trace Metals in Cigarette Litter Collected at McNeese State University in Lake Charles, LA, United States. Methods Objects Chem. Anal. 2022, 17, 5–9. [Google Scholar] [CrossRef]
- St. Louis, V.L.; Rudd, J.W.; Kelly, C.A.; Beaty, K.G.; Bloom, N.S.; Flett, R.J. Importance of Wetlands as Sources of Methyl Mercury to Boreal Forest Ecosystems. Can. J. Fish. Aquat. Sci. 1994, 51, 1065–1076. [Google Scholar] [CrossRef]
- Mierle, G.; Ingram, R. The Role of Humic Substances in the Mobilization of Mercury from Watersheds. Water Air Soil Pollut. 1991, 56, 349–357. [Google Scholar] [CrossRef]
- Compeau, G.C.; Bartha, R. Sulfate-Reducing Bacteria: Principal Methylators of Mercury in Anoxic Estuarine Sediment. Appl. Environ. Microbiol. 1985, 50, 498–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloom, N.S.; Gill, G.A.; Cappellino, S.; Dobbs, C.; McShea, L.; Driscoll, C.; Mason, R.; Rudd, J. Speciation and Cycling of Mercury in Lavaca Bay, Texas, Sediments. Environ. Sci. Technol. 1999, 33, 7–13. [Google Scholar] [CrossRef]
- Bussan, D.D.; Sessums, R.F.; Cizdziel, J.V. Activated Carbon and Biochar Reduce Mercury Methylation Potentials in Aquatic Sediments. Bull. Environ. Contam. Toxicol. 2016, 96, 536–539. [Google Scholar] [CrossRef]
- Benoit, J.M.; Gilmour, C.C.; Heyes, A.; Mason, R.P.; Miller, C. Biogeochemistry of Environmentally Important Trace Elements; American Chemical Society: Washington, DC, USA, 2003; Chapter 19; pp. 262–297. [Google Scholar]
- Rodríguez Martín-Doimeadios, R.C.; Stoichev, T.; Krupp, E.; Amouroux, D.; Holeman, M.; Donard, O.F.X. Working Methods Paper: Micro-scale Preparation and Characterization of Isotopically Enriched Monomethylmercury. Appl. Organomet. Chem. 2002, 16, 610–615. [Google Scholar] [CrossRef]
- Fitzgerald, W.F.; Lamborg, C.H.; Hammerschmidt, C.R. Marine Biogeochemical Cycling of Mercury. Chem. Rev. 2007, 107, 641–662. [Google Scholar] [CrossRef] [PubMed]
- Hintelmann, H.; Evans, R.D. Application of Stable Isotopes in Environmental Tracer Studies–Measurement of Monomethylmercury (CH3Hg+) by Isotope Dilution ICP-MS and Detection of Species Transformation. Fresenius’ J. Anal. Chem. 1997, 358, 378–385. [Google Scholar] [CrossRef]
- US Environmental Protection Agency. Method 1630: Methyl Mercury in Water by Distillation, Aqueous Ethylation, Purge and Trap, and Cold Vapor Atomic Fluorescence Spectrometry; US Environmental Protection Agency: Washington DC, USA, 1998; pp. 1–55. Available online: https://www.epa.gov/sites/default/files/2015-08/documents/method_1630_1998.pdf (accessed on 31 July 2022).
- Lambertsson, L.; Lundberg, E.; Nilsson, M.; Frech, W. Applications of Enriched Stable Isotope Tracers in Combination with Isotope Dilution GC-ICP-MS to Study Mercury Species Transformation in Sea Sediments during in Situ Ethylation and Determination. J. Anal. At. Spectrom. 2001, 16, 1296–1301. [Google Scholar] [CrossRef]
- Haynes, W.M.; Lide, D.R.; Bruno, T.J. CRC Handbook of Chemistry and Physics; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Bussan, D.D.; Sessums, R.F.; Cizdziel, J.V. Direct Mercury Analysis in Environmental Solids by ICP-MS with on-Line Sample Ashing and Mercury Pre-Concentration Using a Direct Mercury Analyzer. J. Anal. At. Spectrom. 2015, 30, 1668–1672. [Google Scholar] [CrossRef]
- Hintelmann, H.; Keppel-Jones, K.; Evans, R.D. Constants of Mercury Methylation and Demethylation Rates in Sediments and Comparison of Tracer and Ambient Mercury Availability. Environ. Toxicol. Chem. Int. J. 2000, 19, 2204–2211. [Google Scholar] [CrossRef]
- Chen, J.; Chakravarty, P.; Davidson, G.R.; Wren, D.G.; Locke, M.A.; Zhou, Y.; Brown Jr, G.; Cizdziel, J.V. Simultaneous Determination of Mercury and Organic Carbon in Sediment and Soils Using a Direct Mercury Analyzer Based on Thermal Decomposition–Atomic Absorption Spectrophotometry. Anal. Chim. Acta 2015, 871, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Huggett, D.B.; Steevens, J.A.; Allgood, J.C.; Lutken, C.B.; Grace, C.A.; Benson, W.H. Mercury in Sediment and Fish from North Mississippi Lakes. Chemosphere 2001, 42, 923–929. [Google Scholar] [CrossRef]
- Hall, B.D.; Aiken, G.R.; Krabbenhoft, D.P.; Marvin-DiPasquale, M.; Swarzenski, C.M. Wetlands as Principal Zones of Methylmercury Production in Southern Louisiana and the Gulf of Mexico Region. Environ. Pollut. 2008, 154, 124–134. [Google Scholar] [CrossRef]
- Callister, S.M.; Winfrey, M.R. Microbial Methylation of Mercury in Upper Wisconsin River Sediments. Water Air Soil Pollut. 1986, 29, 453–465. [Google Scholar] [CrossRef]
- Megonigal, J.P.; Faulkner, S.P.; Patrick, W.H. The Microbial Activity Season in Southeastern Hydric Soils. Soil Sci. Soc. Am. J. 1996, 60, 1263–1266. [Google Scholar] [CrossRef] [Green Version]
- Canfield, D.E. Isotope Fractionation by Natural Populations of Sulfate-Reducing Bacteria. Geochim. Cosmochim. Acta 2001, 65, 1117–1124. [Google Scholar] [CrossRef]
- Marvin-DiPasquale, M.; Agee, J.; McGowan, C.; Oremland, R.S.; Thomas, M.; Krabbenhoft, D.; Gilmour, C.C. Methyl-Mercury Degradation Pathways: A Comparison among Three Mercury-Impacted Ecosystems. Environ. Sci. Technol. 2000, 34, 4908–4916. [Google Scholar] [CrossRef]
- Spangler, W.J.; Spigarelli, J.L.; Rose, J.M.; Miller, H.M. Methylmercury: Bacterial Degradation in Lake Sediments. Science 1973, 180, 192–193. [Google Scholar] [CrossRef]
- Robinson, J.B.; Tuovinen, O. Mechanisms of Microbial Resistance and Detoxification of Mercury and Organomercury Compounds: Physiological, Biochemical, and Genetic Analyses. Microbiol. Rev. 1984, 48, 95–124. [Google Scholar] [CrossRef]
- Barkay, T.; Turner, R.R.; VandenBrook, A.; Liebert, C. The Relationships of Hg (II) Volatilization from a Freshwater Pond to the Abundance Ofmer Genes in the Gene Pool of the Indigenous Microbial Community. Microb. Ecol. 1991, 21, 151–161. [Google Scholar] [CrossRef]
- Begley, T.P.; Walts, A.E.; Walsh, C.T. Bacterial Organomercurial Lyase: Overproduction, Isolation, and Characterization. Biochemistry 1986, 25, 7186–7192. [Google Scholar] [CrossRef] [PubMed]
- Xun, L.; Campbell, N.E.R.; Rudd, J.W. Measurements of Specific Rates of Net Methyl Mercury Production in the Water Column and Surface Sediments of Acidified and Circumneutral Lakes. Can. J. Fish. Aquat. Sci. 1987, 44, 750–757. [Google Scholar] [CrossRef]
- Schartup, A.T.; Mason, R.P.; Balcom, P.H.; Hollweg, T.A.; Chen, C.Y. Methylmercury Production in Estuarine Sediments: Role of Organic Matter. Environ. Sci. Technol. 2013, 47, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollweg, T.A.; Gilmour, C.C.; Mason, R.P. Methylmercury Production in Sediments of Chesapeake Bay and the Mid-Atlantic Continental Margin. Mar. Chem. 2009, 114, 86–101. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).