
Bioinformatics-Guided Identification of Ethyl Acetate Extract of Citri Reticulatae Pericarpium as a Functional Food Ingredient with Anti-Inflammatory Potential

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
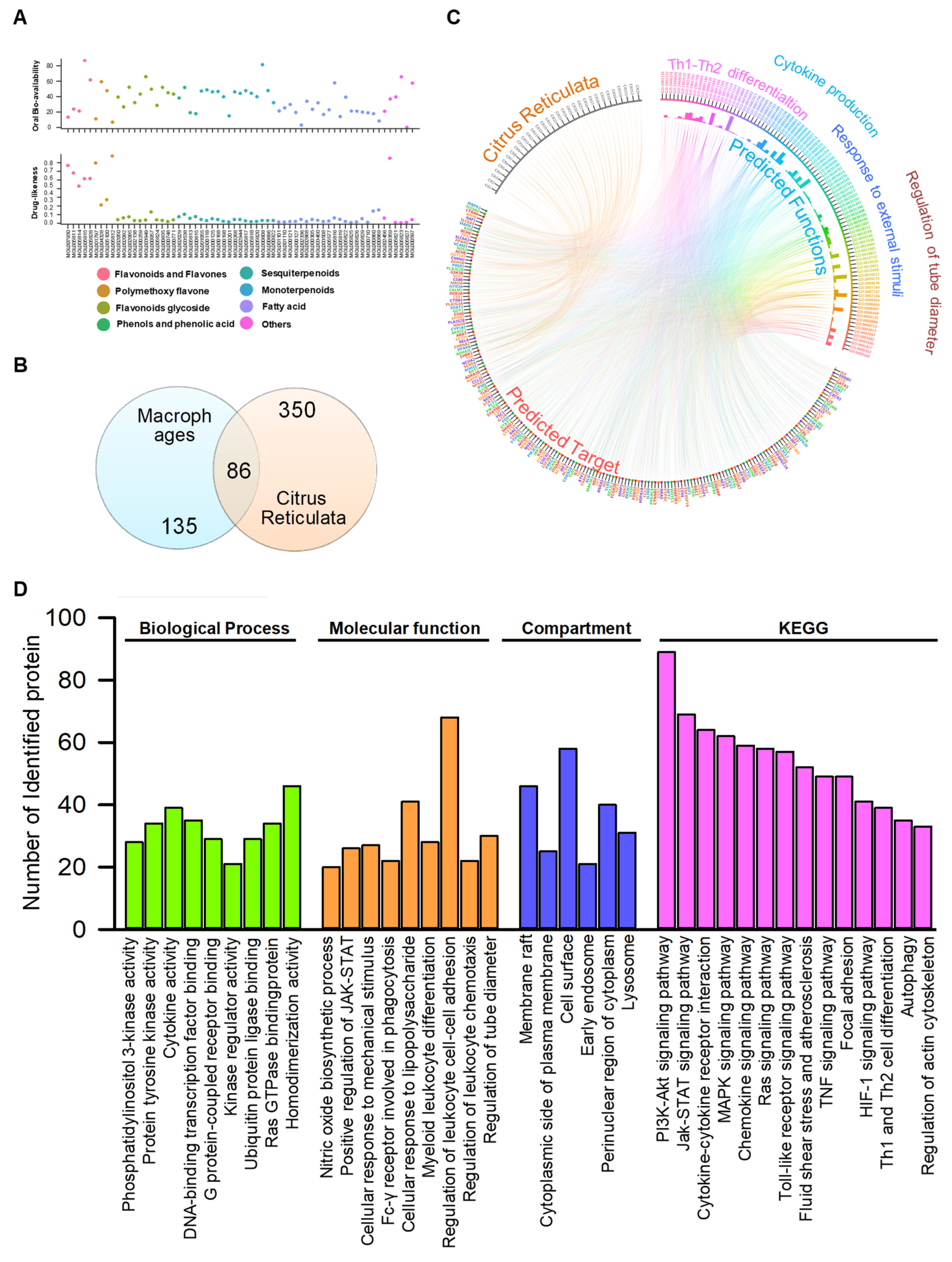
2.1. Network Pharmacological Analysis Predicted TLR4, PI3K/Akt, JAK/STATs, MAPK, and mTOR Signaling Pathways Were Closely Related to the Anti-Inflammatory Function of Citri Reticulatae Pericarpium Extract
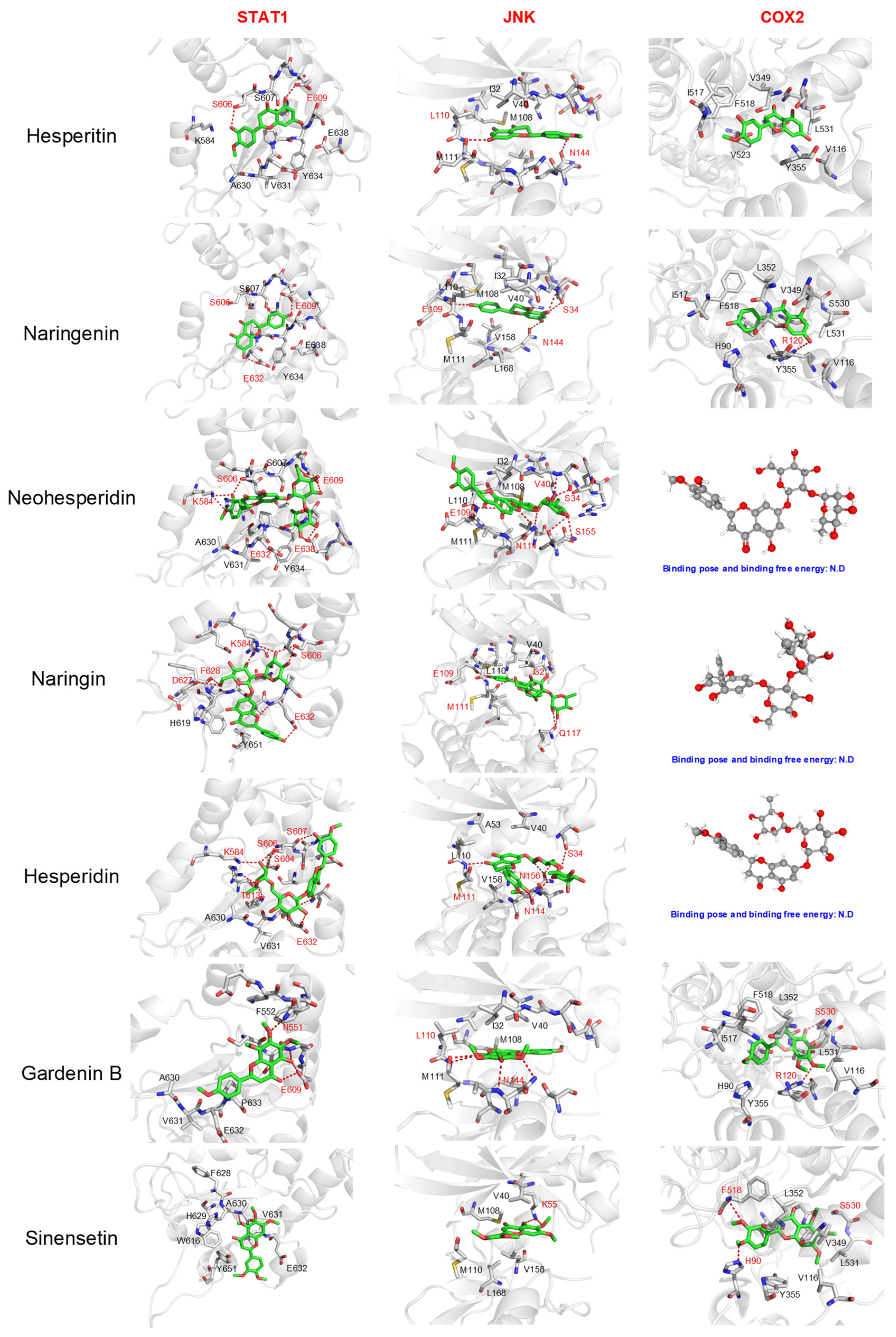
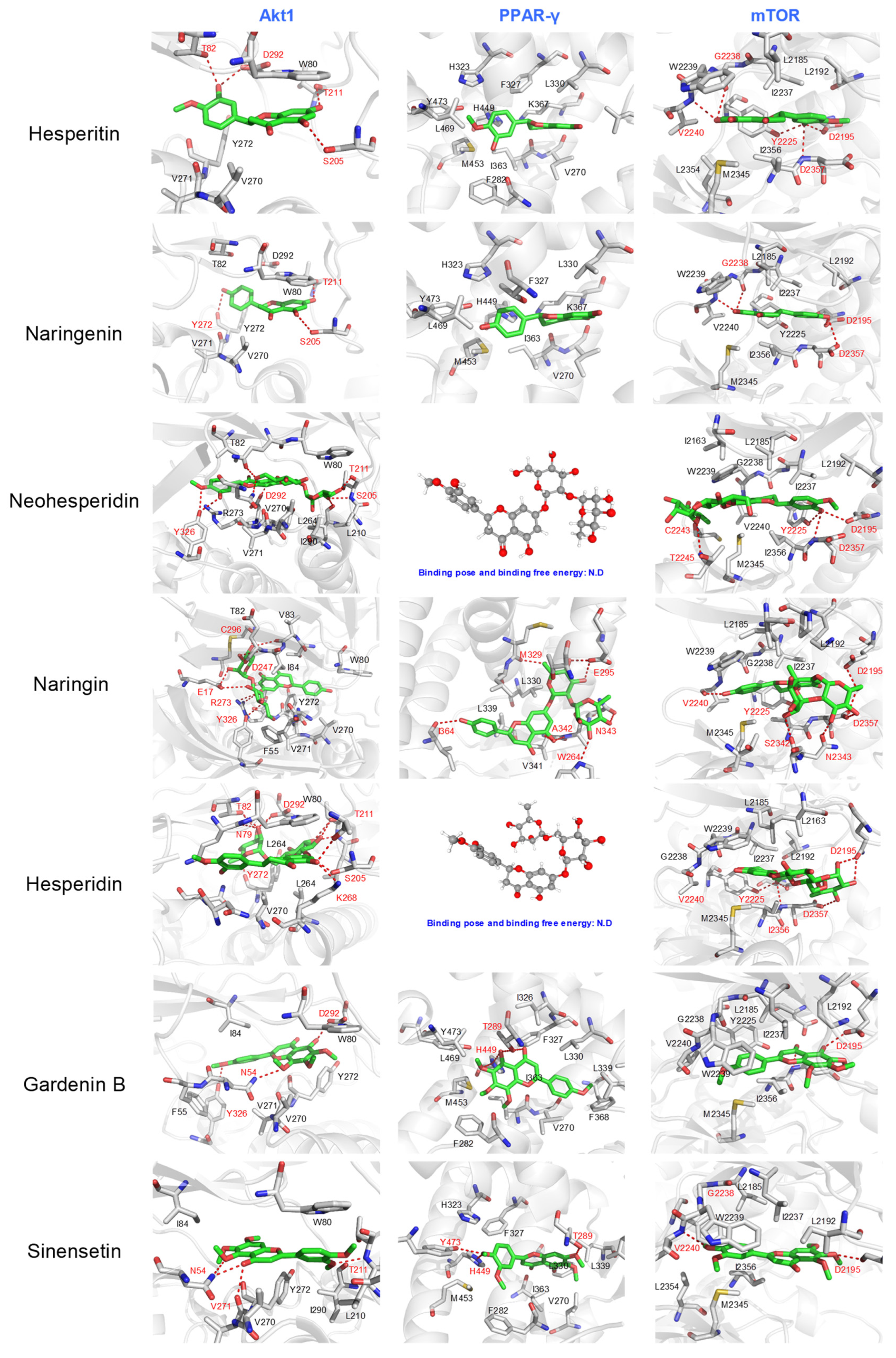
2.2. The Proposed Bioactive Candidate Identification Model Separated the Identified Compounds into Four Clusters According to Their Docking Score Profiles
2.3. In Vitro Anti-Inflammatory Evaluation of Reference Compounds Confirmed That the Cluster Comprising Flavonoids, Phenols, and Phenolic Acid Might Possess Potential Anti-Inflammatory Functions
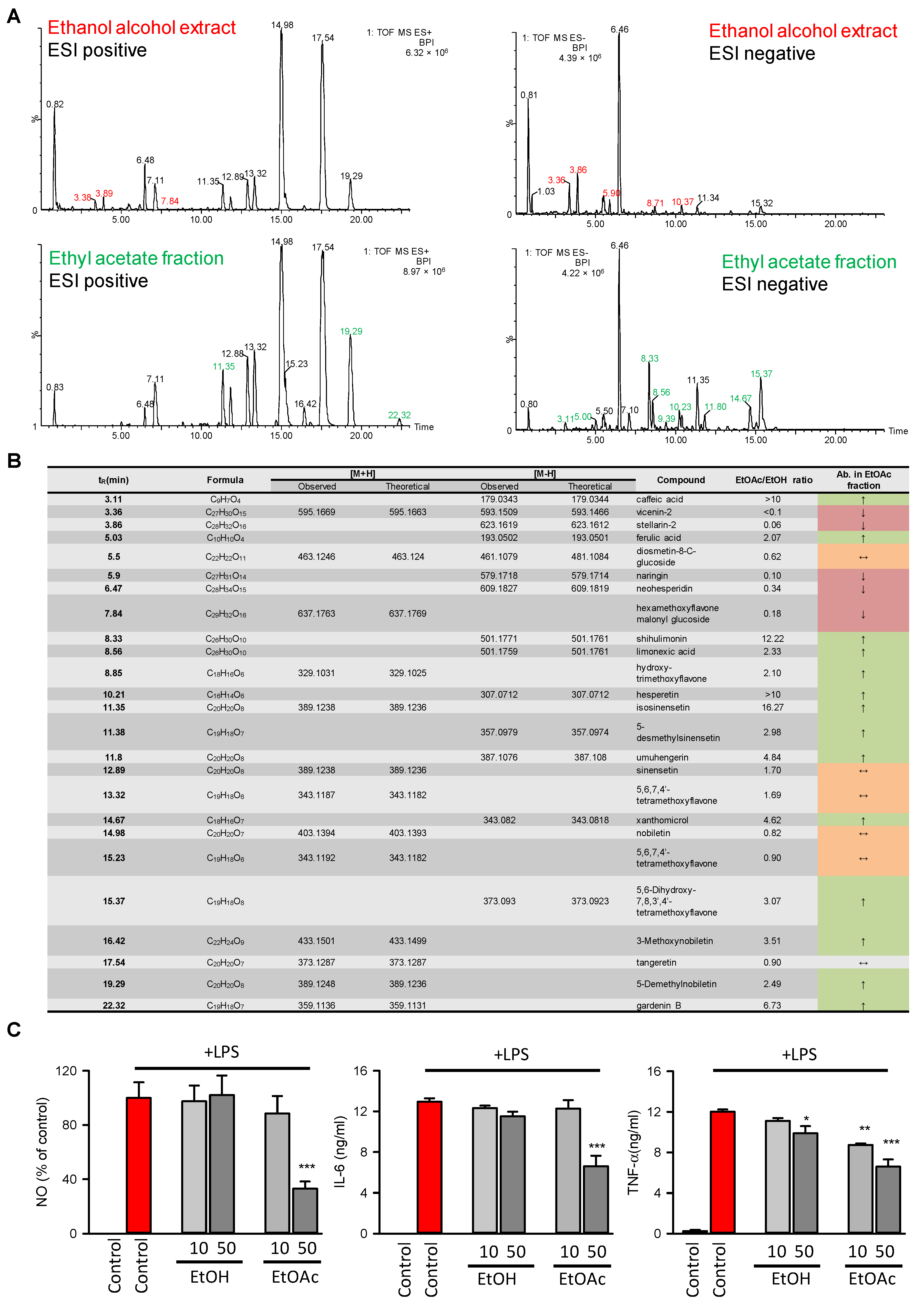
2.4. Chemical Profiling and In Vitro Anti-Inflammatory Test Demonstrated That Flavonoids and Phenols Were Enriched in the EtOAc Extract, Which Might Contribute to its Increased Anti-Inflammatory Potential
3. Materials and Methods
3.1. Chemical Reagents and Extracts
3.2. Network Pharmacology Analysis
3.3. Molecular Docking and Analysis
3.4. Cell Culture and Treatment
3.5. Analysis of Macrophage Cytokines and NO Production
3.6. HPLC/MS/MS-Based Chemical Characterization of Crude Alcoholic Extract of Citri Reticulatae Pericarpium and Its EtOAc Extract
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A guiding map for inflammation. Nat. Immunol. 2017, 18, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Fakhoury, M. Role of Immunity and Inflammation in the Pathophysiology of Neurodegenerative Diseases. Neuro-Degener. Dis. 2015, 15, 63–69. [Google Scholar] [CrossRef]
- Schett, G.; Neurath, M.F. Resolution of chronic inflammatory disease: Universal and tissue-specific concepts. Nat. Commun. 2018, 9, 3261. [Google Scholar] [CrossRef] [PubMed]
- Allison, A.C.; Ferluga, J.; Prydz, H.; Schorlemmer, H.U. The role of macrophage activation in chronic inflammation. Agents Actions 1978, 8, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Parisi, L.; Gini, E.; Baci, D.; Tremolati, M.; Fanuli, M.; Bassani, B.; Farronato, G.; Bruno, A.; Mortara, L. Macrophage Polarization in Chronic Inflammatory Diseases: Killers or Builders? J. Immunol. Res. 2018, 2018, 8917804. [Google Scholar] [CrossRef] [PubMed]
- Koh, T.J.; DiPietro, L.A. Inflammation and wound healing: The role of the macrophage. Expert Rev. Mol. Med. 2011, 13, e23. [Google Scholar] [CrossRef]
- Dinarello, C.A. Anti-inflammatory Agents: Present and Future. Cell 2010, 140, 935–950. [Google Scholar] [CrossRef]
- Marsico, F.; Paolillo, S.; Filardi, P.P. NSAIDs and cardiovascular risk. J. Cardiovasc. Med. 2017, 18 (Suppl. S1), e40–e43. [Google Scholar] [CrossRef]
- Serafini, M.; Peluso, I. Functional Foods for Health: The Interrelated Antioxidant and Anti-Inflammatory Role of Fruits, Vegetables, Herbs, Spices and Cocoa in Humans. Curr. Pharm. Des. 2016, 22, 6701–6715. [Google Scholar] [CrossRef]
- Mijan, M.A.; Lim, B.O. Diets, functional foods, and nutraceuticals as alternative therapies for inflammatory bowel disease: Present status and future trends. World J. Gastroenterol. 2018, 24, 2673–2685. [Google Scholar] [CrossRef]
- Yu, X.; Sun, S.; Guo, Y.Y.; Liu, Y.; Yang, D.Y.; Li, G.Y.; Lu, S.W. Citri Reticulatae Pericarpium (Chenpi): Botany, ethnopharmacology, phytochemistry, and pharmacology of a frequently used traditional Chinese medicine. J. Ethnopharmacol. 2018, 220, 265–282. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Xie, X.; Cai, J.; Zhao, Y.; Miao, W.; Chen, X.; Xiao, Y.; Li, N.; Wu, J.L. Dynamic changes of phenolic acids and antioxidant activity of Citri Reticulatae Pericarpium during aging processes. Food Chem. 2022, 373, 131399. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Chen, Y.L.; Zhang, M.W.; Xu, Y.J.; Pan, S.Y. Chemical Composition, Antioxidant and Antimicrobial Activity of Pericarpium Citri Reticulatae Essential Oil. Molecules 2011, 16, 4082–4096. [Google Scholar] [CrossRef]
- Zheng, G.D.; Zhou, P.; Yang, H.; Li, Y.S.; Li, P.; Liu, E.H. Rapid resolution liquid chromatography-electrospray ionisation tandem mass spectrometry method for identification of chemical constituents in Citri Reticulatae Pericarpium. Food Chem. 2013, 136, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Gosslau, A.; Chen, K.Y.; Ho, C.-T.; Li, S. Anti-inflammatory effects of characterized orange peel extracts enriched with bioactive polymethoxyflavones. Food Sci. Hum. Wellness 2014, 3, 26–35. [Google Scholar] [CrossRef]
- Banjoo, D.R.; Nelson, P.K. Improved ultrasonic extraction procedure for the determination of polycyclic aromatic hydrocarbons in sediments. J. Chromatogr. A 2005, 1066, 9–18. [Google Scholar] [CrossRef]
- Shi, X.; Niu, L.; Zhao, L.; Wang, B.; Jin, Y.; Li, X. The antiallergic activity of flavonoids extracted from Citri Reticulatae Pericarpium. J. Food Processing Preserv. 2018, 42, e13588. [Google Scholar] [CrossRef]
- Shi, Q.; Liu, Z.; Yang, Y.; Geng, P.; Zhu, Y.Y.; Zhang, Q.; Bai, F.; Bai, G. Identification of anti-asthmatic compounds in Pericarpium citri reticulatae and evaluation of their synergistic effects. Acta Pharmacol. Sin. 2009, 30, 567–575. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Li, J.-M.; Chen, B.-Z.; Zhang, X.-M.; Wu, R.-H.; Wu, P.-P.; Li, C.; Chen, W.-H. Recent Advance in the biological activity of synephrine in Citri Reticulatae Pericarpium. Eur. J. Med. Chem. Rep. 2022, 5, 100061. [Google Scholar] [CrossRef]
- Wu, J.; Ye, X.; Yang, S.; Yu, H.; Zhong, L.; Gong, Q. Systems Pharmacology Study of the Anti-Liver Injury Mechanism of Citri Reticulatae Pericarpium. Front. Pharmacol. 2021, 12, 618846. [Google Scholar] [CrossRef]
- Zhang, W.; Huai, Y.; Miao, Z.; Qian, A.; Wang, Y. Systems Pharmacology for Investigation of the Mechanisms of Action of Traditional Chinese Medicine in Drug Discovery. Front. Pharmacol. 2019, 10, 743. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.Y.; Zhang, H.X.; Mezei, M.; Cui, M. Molecular docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided. Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Athar, M.; Sona, A.N.; Bekono, B.D.; Ntie-Kang, F. Fundamental physical and chemical concepts behind “drug-likeness” and “natural product-likeness”. Phys. Sci. Rev. 2019, 4, 20180101. [Google Scholar] [CrossRef]
- Han, M.S.; Jung, D.Y.; Morel, C.; Lakhani, S.A.; Kim, J.K.; Flavell, R.A.; Davis, R.J. JNK expression by macrophages promotes obesity-induced insulin resistance and inflammation. Science 2013, 339, 218–222. [Google Scholar] [CrossRef]
- Zhou, D.; Huang, C.; Lin, Z.; Zhan, S.; Kong, L.; Fang, C.; Li, J. Macrophage polarization and function with emphasis on the evolving roles of coordinated regulation of cellular signaling pathways. Cell Signal 2014, 26, 192–197. [Google Scholar] [CrossRef]
- Irey, E.A.; Lassiter, C.M.; Brady, N.J.; Chuntova, P.; Wang, Y.; Knutson, T.P.; Henzler, C.; Chaffee, T.S.; Vogel, R.I.; Nelson, A.C.; et al. JAK/STAT inhibition in macrophages promotes therapeutic resistance by inducing expression of protumorigenic factors. Proc. Natl. Acad. Sci. USA 2019, 116, 12442–12451. [Google Scholar] [CrossRef]
- Linton, M.F.; Moslehi, J.J.; Babaev, V.R. Akt Signaling in Macrophage Polarization, Survival, and Atherosclerosis. Int. J. Mol. Sci. 2019, 20, 2703. [Google Scholar] [CrossRef]
- Li, S.; Zhang, B. Traditional Chinese medicine network pharmacology: Theory, methodology and application. Chin J. Nat. Med. 2013, 11, 110–120. [Google Scholar] [CrossRef]
- Baier, A.; Szyszka, R. Compounds from Natural Sources as Protein Kinase Inhibitors. Biomolecules 2020, 10, 1546. [Google Scholar] [CrossRef]
- Benavente-Garcia, O.; Castillo, J. Update on uses and properties of citrus flavonoids: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agr. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.C.; Kuo, C.T. Hesperidin, nobiletin, and tangeretin are collectively responsible for the anti-neuroinflammatory capacity of tangerine peel (Citri reticulatae pericarpium). Food Chem. Toxicol. 2014, 71, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Tominari, T.; Hirata, M.; Matsumoto, C.; Inada, M.; Miyaura, C. Polymethoxy flavonoids, nobiletin and tangeretin, prevent lipopolysaccharide-induced inflammatory bone loss in an experimental model for periodontitis. J. Pharmacol. Sci. 2012, 119, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Ogawa, M.; Amakura, Y.; Yoshimura, M.; Okuyama, S.; Furukawa, Y. 3,5,6,7,8,3′,4′-Heptamethoxyflavone reduces interleukin-4 production in the spleen cells of mice. Biomed. Res. 2016, 37, 95–99. [Google Scholar] [CrossRef]
- Ren, H.; Hao, J.; Liu, T.; Zhang, D.; Lv, H.; Song, E.; Zhu, C. Hesperetin Suppresses Inflammatory Responses in Lipopolysaccharide-Induced RAW 264.7 Cells via the Inhibition of NF-kappaB and Activation of Nrf2/HO-1 Pathways. Inflammation 2016, 39, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Bodet, C.; La, V.D.; Epifano, F.; Grenier, D. Naringenin has anti-inflammatory properties in macrophage and ex vivo human whole-blood models. J. Periodontal Res. 2008, 43, 400–407. [Google Scholar] [CrossRef]
- Tutunchi, H.; Naeini, F.; Ostadrahimi, A.; Hosseinzadeh-Attar, M.J. Naringenin, a flavanone with antiviral and anti-inflammatory effects: A promising treatment strategy against COVID-19. Phytother. Res. PTR 2020, 34, 3137–3147. [Google Scholar] [CrossRef]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and Anti-Inflammatory Properties of the Citrus Flavonoids Hesperidin and Hesperetin: An Updated Review of their Molecular Mechanisms and Experimental Models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef]
- Hirata, A.; Murakami, Y.; Shoji, M.; Kadoma, Y.; Fujisawa, S. Kinetics of radical-scavenging activity of hesperetin and hesperidin and their inhibitory activity on COX-2 expression. Anticancer. Res. 2005, 25, 3367–3374. [Google Scholar]
- Liang, S.J.; Wen, Z.J.; Tang, T.X.; Liu, Y.F.; Dang, F.L.; Xie, T.X.; Wu, H. Study on flavonoid and bioactivity features of the pericarp of Citri Reticulatae ‘chachi’ during storage. Arab. J. Chem. 2022, 15, 103653. [Google Scholar] [CrossRef]
- Hao da, C.; Xiao, P.G. Network pharmacology: A Rosetta Stone for traditional Chinese medicine. Drug Dev. Res. 2014, 75, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Seto, S.W.; Chang, D.; Kiat, H.; Razmovski-Naumovski, V.; Chan, K.; Bensoussan, A. Synergistic Effects of Chinese Herbal Medicine: A Comprehensive Review of Methodology and Current Research. Front. Pharmacol. 2016, 7, 201. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.; Natoli, G. Transcriptional regulation of macrophage polarization: Enabling diversity with identity. Nat. Rev. Immunol. 2011, 11, 750–761. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Schmidt, S.V.; Sander, J.; Draffehn, A.; Krebs, W.; Quester, I.; De Nardo, D.; Gohel, T.D.; Emde, M.; Schmidleithner, L.; et al. Transcriptome-based network analysis reveals a spectrum model of human macrophage activation. Immunity 2014, 40, 274–288. [Google Scholar] [CrossRef]
- Jin, L.; Schmiech, M.; El Gaafary, M.; Zhang, X.; Syrovets, T.; Simmet, T. A comparative study on root and bark extracts of Eleutherococcus senticosus and their effects on human macrophages. Phytomedicine 2020, 68, 153181. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Y.; Wu, J.; Guo, Y.; Xiao, H.; Zhang, Y.; Ma, K. Resveratrol Targets AKT1 to Inhibit Inflammasome Activation in Cardiomyocytes Under Acute Sympathetic Stress. Front. Pharmacol. 2022, 13, 818127. [Google Scholar] [CrossRef]
- Rehan, M. Anticancer compound XL765 as PI3K/mTOR dual inhibitor: A structural insight into the inhibitory mechanism using computational approaches. PLoS ONE 2019, 14, e0219180. [Google Scholar] [CrossRef]
- Wang, W.; Xiong, X.; Li, X.; Zhang, Q.; Yang, W.; Du, L. In Silico Investigation of the Anti-Tumor Mechanisms of Epigallocatechin-3-Gallate. Molecules 2019, 24, 1445. [Google Scholar] [CrossRef]
- Garvie, C.W.; Wu, X.; Papanastasiou, M.; Lee, S.; Fuller, J.; Schnitzler, G.R.; Horner, S.W.; Baker, A.; Zhang, T.; Mullahoo, J.P.; et al. Structure of PDE3A-SLFN12 complex reveals requirements for activation of SLFN12 RNase. Nat. Commun. 2021, 12, 4375. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.J.; Leung, K.H.; Chan, D.S.; Wang, Y.T.; Ma, D.L.; Leung, C.H. Identification of a natural product-like STAT3 dimerization inhibitor by structure-based virtual screening. Cell Death Dis. 2014, 5, e1293. [Google Scholar] [CrossRef] [PubMed]
- Jha, V.; Macchia, M.; Tuccinardi, T.; Poli, G. Three-Dimensional Interactions Analysis of the Anticancer Target c-Src Kinase with Its Inhibitors. Cancers 2020, 12, 2327. [Google Scholar] [CrossRef] [PubMed]
- Maltarollo, V.G.; Togashi, M.; Nascimento, A.S.; Honorio, K.M. Structure-based virtual screening and discovery of New PPARdelta/gamma dual agonist and PPARdelta and gamma agonists. PLoS ONE 2015, 10, e0118790. [Google Scholar] [CrossRef]
- Espinoza-Culupu, A.; Vazquez-Ramirez, R.; Farfan-Lopez, M.; Mendes, E.; Notomi Sato, M.; da Silva Junior, P.I.; Borges, M.M. Acylpolyamine Mygalin as a TLR4 Antagonist Based on Molecular Docking and In Vitro Analyses. Biomolecules 2020, 10, 1624. [Google Scholar] [CrossRef]
- Yang, L.; Sun, X.; Ye, Y.; Lu, Y.; Zuo, J.; Liu, W.; Elcock, A.; Zhu, S. p38alpha Mitogen-Activated Protein Kinase Is a Druggable Target in Pancreatic Adenocarcinoma. Front. Oncol. 2019, 9, 1294. [Google Scholar] [CrossRef]
- Sanachai, K.; Mahalapbutr, P.; Choowongkomon, K.; Poo-Arporn, R.P.; Wolschann, P.; Rungrotmongkol, T. Insights into the Binding Recognition and Susceptibility of Tofacitinib toward Janus Kinases. ACS Omega 2020, 5, 369–377. [Google Scholar] [CrossRef]
- Hermawan, A.; Putri, H.; Hanif, N.; Ikawati, M. Integrative Bioinformatics Study of Tangeretin Potential Targets for Preventing Metastatic Breast Cancer. Evid. Based Complement. Altern. Med. 2021, 2021, 2234554. [Google Scholar] [CrossRef]
- Zheng, C.; Wu, X.; Zeng, R.; Lin, L.; Xu, L.; Li, E.; Dong, G. Computational Prediction of Hot Spots and Binding Site of Inhibitor NSC23766 on Rac1 Binding With Tiam1. Front. Chem. 2020, 8, 625437. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; Huang, Z.; Wang, H.; Luo, H.; Wang, X.; Zhou, N.; Wu, C.; Bao, J. Computer-aided identification of potential TYK2 inhibitors from drug database. J. Mol. Struct. 2016, 1122, 309–317. [Google Scholar] [CrossRef]
- Lee, K.H.; Yen, W.C.; Lin, W.H.; Wang, P.C.; Lai, Y.L.; Su, Y.C.; Chang, C.Y.; Wu, C.S.; Huang, Y.C.; Yang, C.M.; et al. Discovery of BPR1R024, an Orally Active and Selective CSF1R Inhibitor that Exhibits Antitumor and Immunomodulatory Activity in a Murine Colon Tumor Model. J. Med. Chem. 2021, 64, 14477–14497. [Google Scholar] [CrossRef] [PubMed]
- Ravelli, R.B.; Gigant, B.; Curmi, P.A.; Jourdain, I.; Lachkar, S.; Sobel, A.; Knossow, M. Insight into tubulin regulation from a complex with colchicine and a stathmin-like domain. Nature 2004, 428, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Kurumbail, R.G.; Stevens, A.M.; Gierse, J.K.; McDonald, J.J.; Stegeman, R.A.; Pak, J.Y.; Gildehaus, D.; Miyashiro, J.M.; Penning, T.D.; Seibert, K.; et al. Structural basis for selective inhibition of cyclooxygenase-2 by anti-inflammatory agents. Nature 1996, 384, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Wang, X.; Li, W.; Ma, X.; Chu, Y.; Li, S.; Guo, J.; Jia, Y.; Zhou, S.; Zhu, Y.; Liu, C. Simultaneous determination of caffeic acid and its major pharmacologically active metabolites in rat plasma by LC-MS/MS and its application in pharmacokinetic study. Biomed Chromatogr. 2015, 29, 552–559. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, X.; Qiu, J.; Zhang, L.; Zhang, Y.; Qiu, X.; Huang, Z.; Xu, W. Comprehensive Comparison on the Chemical Profile of Guang Chen Pi at Different Ripeness Stages Using Untargeted and Pseudotargeted Metabolomics. J. Agr. Food Chem. 2020, 68, 8483–8495. [Google Scholar] [CrossRef]
- Xu, L.L.; Xu, J.J.; Zhong, K.R.; Shang, Z.P.; Wang, F.; Wang, R.F.; Zhang, L.; Zhang, J.Y.; Liu, B. Analysis of Non-Volatile Chemical Constituents of Menthae Haplocalycis Herba by Ultra-High Performance Liquid Chromatography-High Resolution Mass Spectrometry. Molecules 2017, 22, 1756. [Google Scholar] [CrossRef]
- Tang, J.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF/MS Characterization of Phenolic Compounds from Medicinal Plants (Hops and Juniper Berries) and Their Antioxidant Activity. Foods 2019, 9, 7. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, E.; Jin, L.; Qian, C.; Feng, C.; Zhao, Z.; Tian, H.; Yang, D. Bioinformatics-Guided Identification of Ethyl Acetate Extract of Citri Reticulatae Pericarpium as a Functional Food Ingredient with Anti-Inflammatory Potential. Molecules 2022, 27, 5435. https://doi.org/10.3390/molecules27175435
Ma E, Jin L, Qian C, Feng C, Zhao Z, Tian H, Yang D. Bioinformatics-Guided Identification of Ethyl Acetate Extract of Citri Reticulatae Pericarpium as a Functional Food Ingredient with Anti-Inflammatory Potential. Molecules. 2022; 27(17):5435. https://doi.org/10.3390/molecules27175435
Chicago/Turabian StyleMa, Enyao, Lu Jin, Chunguo Qian, Chong Feng, Zhimin Zhao, Hongru Tian, and Depo Yang. 2022. "Bioinformatics-Guided Identification of Ethyl Acetate Extract of Citri Reticulatae Pericarpium as a Functional Food Ingredient with Anti-Inflammatory Potential" Molecules 27, no. 17: 5435. https://doi.org/10.3390/molecules27175435
APA StyleMa, E., Jin, L., Qian, C., Feng, C., Zhao, Z., Tian, H., & Yang, D. (2022). Bioinformatics-Guided Identification of Ethyl Acetate Extract of Citri Reticulatae Pericarpium as a Functional Food Ingredient with Anti-Inflammatory Potential. Molecules, 27(17), 5435. https://doi.org/10.3390/molecules27175435