Abstract
Berberine (BBR), a Chinese herbal medicine used in intestinal infection, has been applied as a botanical pesticide in the prevention of fungal disease in recent years. However, its degradation in the environment remains poorly understood. Here, we investigated BBR’s degradation in soil water from different sources accompanied by its effect on bacterial diversity. Our results indicated that BBR was only degraded in soil water, while it was stable in tap water, river water and aquaculture water. Bacterial amplicon results of these samples suggested that the degradation of BBR was closely related to the enrichment of Methylotenera. To reveal this special relationship, we used bioinformatics tools to make alignments between the whole genome of Methylotenera and the pathway of BBR’s degradation. An ortholog of Tetrahydroisoquinoline N-methyltransferase from plant was discovered only in Methylotenera that catalyzed a crucial step in BBR’s degradation pathway. In summary, our work indicated that Methylotenera was an essential bacterial genus in the degradation of BBR in the environment because of its Tetrahydroisoquinoline N-methyltransferase. This study provided new insights into BBR’s degradation in the environment, laying foundations for its application as a botanical pesticide.
1. Introduction
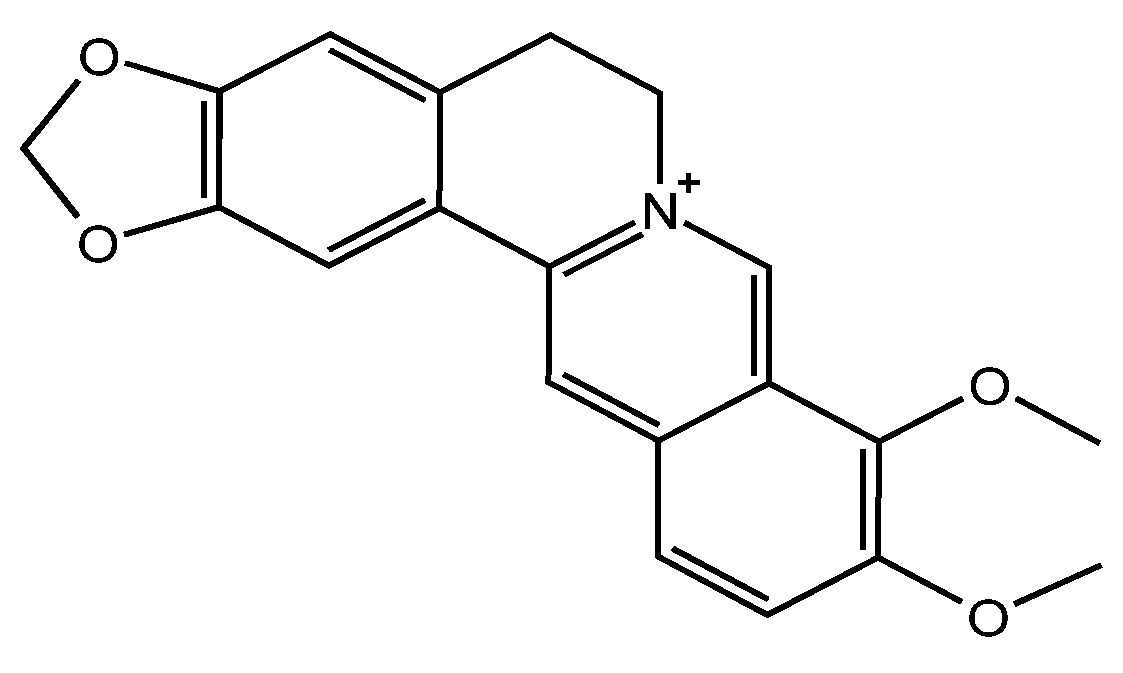
Berberine (BBR, Figure 1) is a plant-derived isoquinoline alkaloid widely distributed in plants of the Berberidaceae, Ranunculaceae and Papaveraceae families [1]. BBR has been used in anti-tumor disease, anti-oxidative stress, anti-Alzheimer’s disease, myocardial tissue protection and anti-hyperglycemic drugs [2,3,4]. Today, its application is extended as a botanical pesticide because of its exceptional antifungal effect [5,6]. The MIC of BBR against Monilinia fructicola is as low as 4.5 µg/mL according to our previous work, showing the potential to prevent fungal disease in peach [7]. With rapidly elevated demand of BBR in agricultural applications, increasing BBR residues remain in the soil and even filter into the underground water [8]. Therefore, the impacts of BBR on the environment need to be further elucidated.
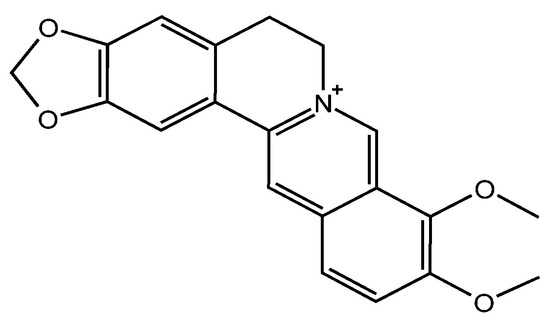
Figure 1.
Confirmed Chemical structure of BBR.
The degradation of BBR in soil was studied in our previous work [6]. We demonstrated that BBR can be completely degraded in soil within 9 days, and soil bacterial diversity was naturally restored as BBR degraded [6]. However, the retention time of BBR in soil was not as long as 9 days in nature, and some residues were filtered into the soil water or even into underground water [9]. When BBR enters water through runoff, leaching and wastewater discharge [8,10], the degradation of BBR in water and its impact on bacterial diversity in the water environment need further investigation. It has been confirmed that the microorganisms that can degrade BBR include Sphingopyxis [11], Burkholderia [12], Rhodococcus [13] and Bacillus [6]. Mixed cultures of Hydrogenophaga, Azoarcus, Sphingopyxis, Stenotrophomonas, Shinella, Alcaligenes and Nitrospir are identified as potential functional species for the biodegradation of BBR and its metabolites [14]. However, the degradation pathway of BBR in bacteria is still unclear. According to the secondary metabolite synthesis pathway in plants (Kyoto Encyclopedia of Genes and Genomes (KEGG): map00950, map01063), BBR’s degradation may rely on multiple steps, while these enzymes are deduced as not being contained by a single bacterium. Therefore, the digestion of BBR in microorganisms needs further study.
The diversity of microbial populations plays an important role in environmental remediation and pesticide degradation [15,16]. The improvement of the microbial community can significantly increase the degradation efficiency of pesticides in the environment [17]. In the case of BBR, there is a possibility that the enzymes essential for the degradation of BBR are distributed in different microorganisms, requiring them to participate in BBR’s degradation simultaneously. On the other hand, there is also a possibility that there is a rate-limiting enzyme that exists in only one or a few microorganisms. When key enzymes are present, the degradation process will be accelerated significantly, and this phenomenon can be also found in the degradation of lignin [18].
Based on these backgrounds, in this study, we aim to investigate BBR’s degradation in soil water from different sources and explore its relationship with bacterial diversity. We anticipate revealing the relationship between BBR’s degradation and the changes in the bacterial content in soil water, revealing the essential step of BBR’s degradation in the environment.
2. Materials and Methods
2.1. Materials
BBR (99%), methanol and phosphoric acid were purchased from Macklin (Shanghai, China). Dimethyl sulfoxide (DMSO) was purchased from Bailens Biotechnology Co., Ltd. (Tianjing, China). DNA primers (Miseq-F and Miseq-R) used for bacterial identification were synthesized by Shanghai Hanyu Biotechnology Co., Ltd. The genomes of each sample were extracted, and then amplicon sequencing was conducted using the two primers Miseq-F:TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGTACGGRAGGCAGCAG and Miseq-R: GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGAGGGTAT CTAATCCT. DNA sequencing was conducted in an Illumina Miseq platform. DNA polymerase and the other agents used for 16s rDNA amplification were products of Takara (Dalian, China).
2.2. Collection and Treatment of Water in Different Environments
Soil samples with a depth of 5 to 10 cm from different locations of China (Xinjiang, Beijing, Jiangxi, Yunnan, Heilongjiang and Guangxi) were collected and mixed evenly. Then, 50 g of the soil was soaked in 500 mL of sterilized water and filtered through a 22 µm filter to eliminate the insoluble particles. Subsequently, 500 mL of fish-cultured water, river water and tap water were respectively filtered and used in the following experiments. BBR was dissolved in DMSO as 10 mg/mL as a stock solution, and the final concentration of BBR in the experiment was set at 100 µg/mL.
2.3. HPLC Analysis of BBR
The standard BBR sample used in HPLC analysis was dissolved in sterile water to the final concentration of 100 µg/mL, and gradient concentrations of BBR samples were diluted and filtered through a 0.22 µm filter membrane before they were injected into the HPLC system. For each sample obtained from the soil water, samples were filtered before injection. The HPLC system (Shimazu, Kyoto, Japan) was equipped with a C18 column with a SPD-20A UV detector at 364 nm wavelength. The column temperature was 30 °C, and the injection volume was 20 μL. The mobile phase was composed of methanol (50%) and 0.05% phosphoric acid water (50%), and the flow rate was set at 0.8 mL/min. This is a commonly used method in our lab [6]. Data in the results were presented as means ± standard errors of mean (SD). Statistical significance was indicated by P values analyzed by one-way ANOVA.
2.4. Degradation Experiment of BBR in Water Samples
Each 20 mL sample of soil water, fish-cultured water, river water and tap water was collected, and the initial concentration of BBR was set at 100 µg/mL. All samples were placed in a constant temperature incubator at 30 °C for 14 days. Samples were taken every 24 h and centrifuged at 12,000 rpm for 5 min, and then filtered through a 0.22 µm membrane for HPLC detection.
2.5. 16s rDNA Sequencing of Soil Water Samples
The genomes of samples from Xinjiang, Beijing, Jiangxi, Yunnan, Heilongjiang and Guangxi were collected and then extracted for amplicon sequencing. Briefly, 1 mL of the soil water samples was obtained from the culture of BBR degradation. The total genome of the soil water samples was extracted using a whole genome extraction kit from Tiangen, China. Next, 2 μL of genome was used as the template for amplicon sequencing. The other agents were added as follows: 25 μL of Ex Taq, 6 μL of Miseq F, 6 μL of Miseq R, 11 μL of sterile water. PCR was carried out as follows: 94 °C for 10 min, 30 cycles (94 °C for 1 min, 55 °C for 40 s, 72 °C for 2 min), 72 °C for 1 min, and 16 °C hold. PCR amplification products were detected by 1% agarose gel electrophoresis, and then were sent to Shanghai Hanyu Biotechnology Co., Ltd. for 16s rDNA amplicon sequencing. The content of each genus was calculated by comparing the number of sequences identified belonging to certain genera and the total number of sequences in the same sample.
Soil water samples (Xinjiang, Beijing and Jiangxi) at 0 days, 3 days, 6 days, 10 days and 14 days were cultured in Luria-Bertani (LB) agar plates (NaCl 10 g/L; peptone 10 g/L; yeast extract 5 g/L; agar powder 15 g/L, if possible) at 30 °C for 24 h. Possible BBR-degrading strains with different colonial morphologies were collected and characterized. After purification three times, the genome of the strain was extracted, and 16s rDNA was amplified and sequenced by Sanger sequencing. All the strains were conserved in −80 °C mixed with 15% sterile glycerol.
2.6. Degradation of BBR by Isolated Bacterium
Soil waters were sterilized and then cooled for later use. Strains were grown overnight in LB with a 30 °C shaking incubator (200 rpm) prior to experiment. For the degradation experiment with single and multiple bacterial: 10 mL of the recovered single or mixed bacteria was washed three times and then added to 100 μL of sterilized water, followed by adding 50 μL of each to soil water or LB medium.
Next, the degradation experiment was conducted in the shake flask in the presence of 100 µg/mL BBR. The samples were shaken (200 rpm, 37 °C), and samples were taken at 0 day, 4 days, 8 days, 10 days and 14 days for HPLC analysis.
2.7. Genome Alignment and Homology Modeling
The biosynthesis pathway of BBR in plant (KEGG) was used as the template for detecting enzymes that participated in the degradation of BBR bacteria. Briefly, the enzymes in plants were collected, and the bacteria genomes were prepared for alignment. Then, the enzymes from BBR biosynthesis in plants were used as the template, and the alignment was conducted with the whole genome of each isolated bacterium. The genome alignment was carried out in Python, and the script used in this study was available from Github (https://github.com/liying0128/pathway_finder (accessed on 12 July 2022). Homology modeling was generated from YASARA Structure since we have the license. According to the protein sequences from plant and bacteria, models were generated and visualized after alignment and energy minimization.
3. Results
3.1. BBR Is Degraded Only in Soil Water
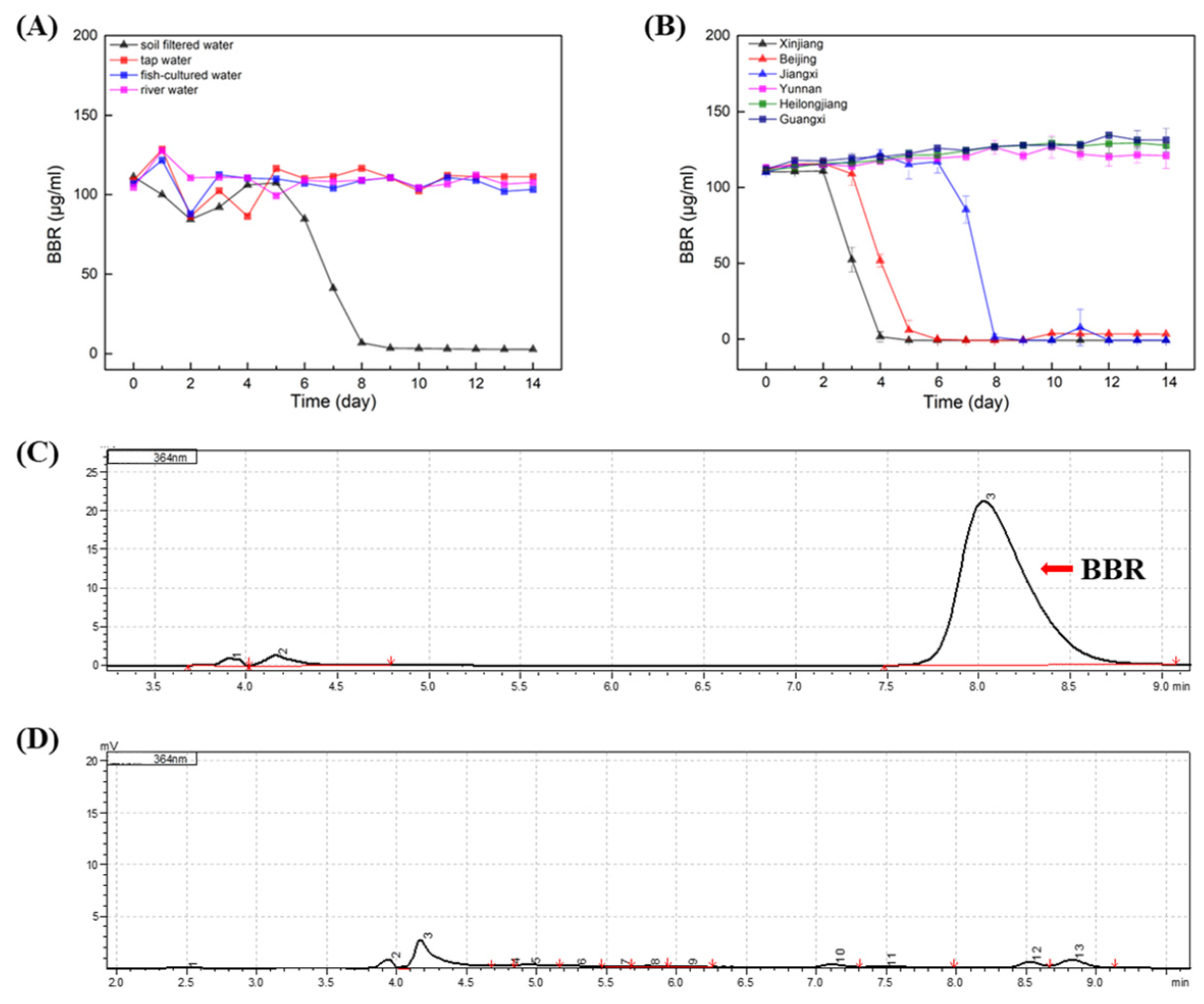
In order to evaluate the effect of BBR on the environment as agricultural applications, the degradation of BBR was carried out on samples of soil water, fish-cultured water, river water and tap water (Figure 2A). The results indicated that stabilities of BBR were distinct in different samples, and BBR’s degradation was only observed in samples of soil water. In soil water, the adaptation period of BBR was nearly 5 days before degradation, while BBR degrades rapidly, within 3 days after the initiation of degradation. It is speculated that the content of microorganisms in the soil water may be rich and that there are single or multiple bacteria that can favor the degradation of BBR.
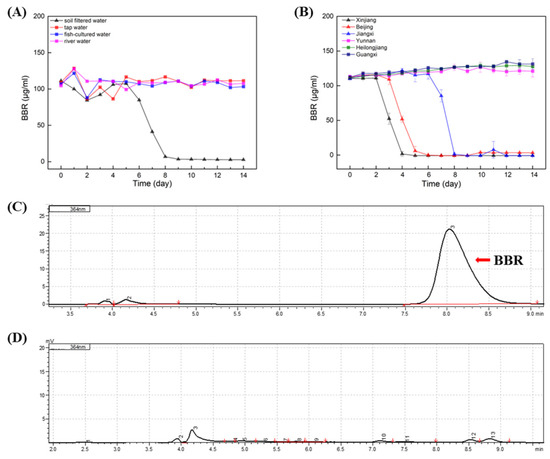
Figure 2.
BBR: Berberine; HPLC: High Performance Liquid Chromatography. BBR degradation in different waters. (A): Degradation of BBR in soil water, fish-cultured water, river water and tap water. (B): Degradation of BBR in soil water samples from different location of China. (C): HPLC analysis of BBR’s concentration in the initial of experiment. (D): HPLC analysis of BBR’s concentration in the end of experiment. Arrow indicated the peak of BBR.
To clarify whether the degradation of BBR by soil water is universal, BBR degradation experiments were carried out in soil water from different regions of China (Xinjiang, Beijing, Jiangxi, Yunnan, Heilongjiang and Guangxi; Figure 2B). Surprisingly, only in three soil water samples was BBR degraded. Notably, the adaption times of BBR degradation in these samples were different, while BBR degraded rapidly within two days after the degradation began. Therefore, it is concluded that the degradation of BBR in soil water is not universal, and it is consistent with Strickland’s conclusion that soil microbial communities in different regions are functionally different [19].
3.2. The Effect of BBR on Bacterial Diversity of Soil Water
Since the degradation of BBR in soil water was different among these samples, we conducted amplicon sequencing of the bacterial diversity on different days to investigate the variations in the contents of the bacteria during BBR’s degradation.
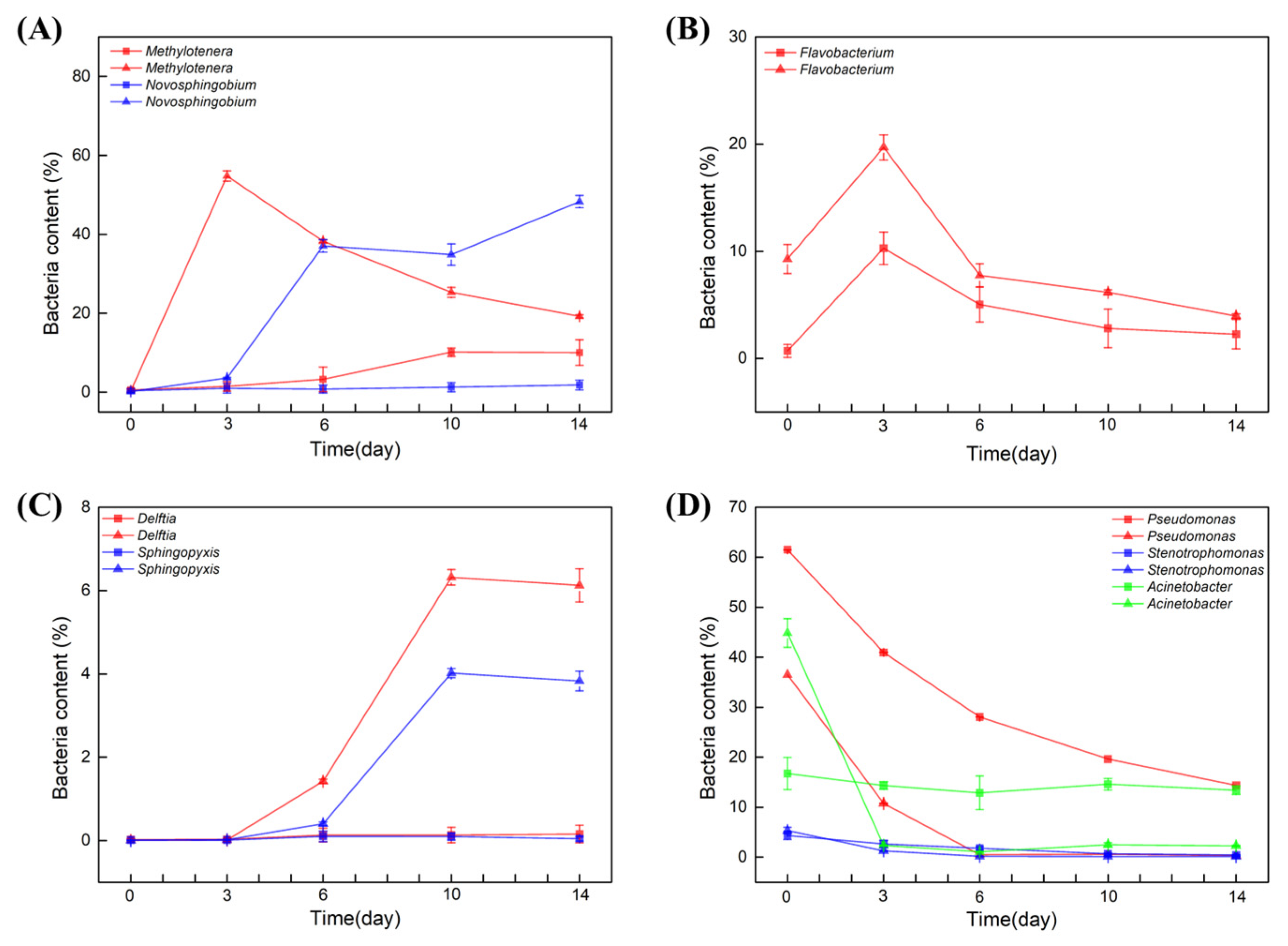
The most significant differences between the samples were the contents of Methylotenera and Novosphingobium in soil water. At day 0, the bacterial content of both Methylotenera and Novosphingobium was nearly 0%. Interestingly, in the soil water in which BBR was degraded, the two strains were both enriched significantly within 6 days. When Methylotenera reached the peak on day 3, the content of Novosphingobium began to increase (Figure 3A). However, in the sample in which BBR was not degraded, the content of these two strains did not change in the first 6 days. Compared with the different degradation rates of BBR, we suspect that the enrichments of Methylotenera and Novosphingobium had positive effects on the degradation of BBR.
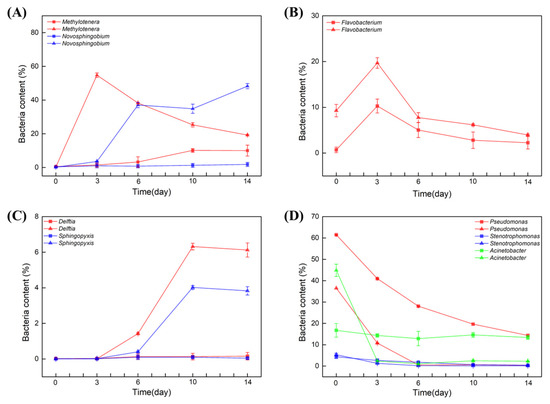
Figure 3.
The effect of BBR on bacterial diversity of soil water. Bacterial content within 14 days: Methylotenera and Novosphingobium (A), Flavobacterium (B), Delftia and sphingopyxis (C), Pseudomonas, Stenotrophomonas and Acinetobacter (D). Triangular pattern represents BBR degraded samples. Square pattern represents BBR undegraded samples.
In contrast, Flavobacterium genus in both groups began to grow on day 0 and reached a peak on day 3, but its content in the samples from Xinjiang, Beijing and Jiangxi was two times higher than that in the samples from Yunnan, Heilongjiang and Guangxi. However, its content decreased from day 3 to the end (Figure 3B). It is deduced that the presence of BBR stimulated the enrichment of Flavobacterium. In parallel, the bacterial contents of Delftia and sphingopyxis were nearly 0% in the initial of the experiments. In the samples from Xinjiang, Beijing and Jiangxi, the contents of these two strains began to increase after 3 daysof cultivation, and the increasing rate was even faster after 6 days, while its content in the samples from Yunnan, Heilongjiang and Guangxi did not change (Figure 3C). Therefore, these two strains might also have positive effects on the degradation of BBR.
The three genera of Pseudomonas, Stenotrophomonas and Acinetobacter were all detected in the initial of the two experiments. The bacterial contents of Pseudomonas and Stenotrophomonas were decreased in all of the samples. Different from these two strains, the content of Acinetobacter decreased rapidly to 0% in the samples from Xinjiang, Beijing and Jiangxi within 3 days, but its contents in the samples from Yunnan, Heilongjiang and Guangxi did not change (Figure 3D).
To sum up, Acinetobacter was abundant in initial soil water samples, while in content, it decreased sharply within 3 days in the samples from Xinjiang, Beijing and Jiangxi. In contrast, Novosphingobium, Delftia and sphingopyxis started to grow after day 3. Notably, the bacterial contents of Methylotenera increased sharply in the presence of BBR but decreased with the degradation of BBR.
3.3. Isolated Bacteria Were Unable to Degrade BBR
To explore whether single or multiple bacteria can degrade BBR in an artificial medium, bacteria were isolated from the soil water in which BBR was degraded, and these bacteria were named J1–8. DNA sequence alignment showed that these strains belonged to three different genera (Table 1)

Table 1.
Bacteria isolated from BBR-degrading soil water.
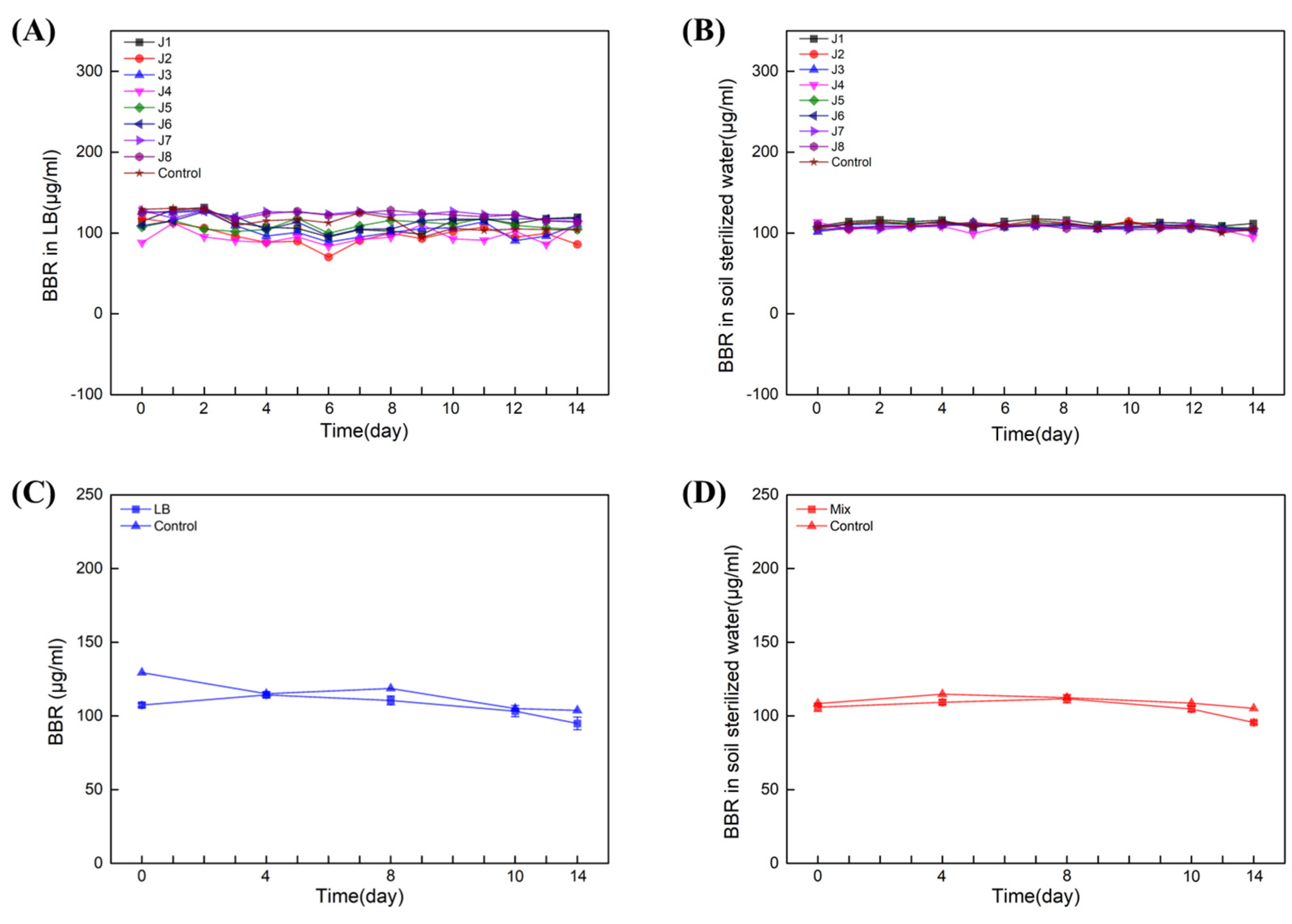
Next, single- and multiple-degradation experiments (J1–8) were carried out for 14 days in LB liquid medium and sterilized soil water (both contain 100 µL/mL BBR), respectively. The results showed that in LB liquid medium (Figure 4A) and soil-sterilized water (Figure 4B), neither of the single strains could degrade BBR. Next, multiple-degradation experiments (containing all of J1–8) were also carried out in LB liquid medium and soil-sterilized water with 100 μg/mL of BBR. Similar results were obtained and BBR was not degraded in presence of all these strains (Figure 4C,D). Together with the data from amplicon sequencing, we can conclude that the essential bacteria for BBR’s degradation might be unable to be cultivated in artificial medium.
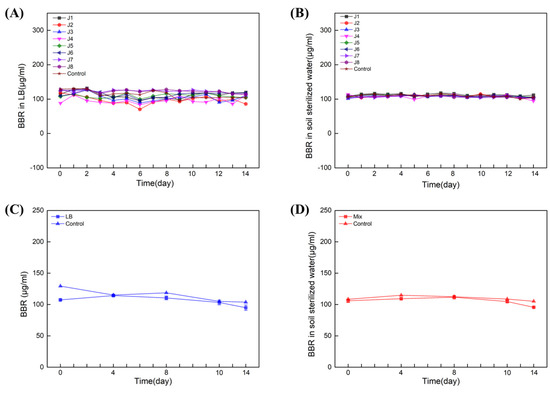
Figure 4.
Eight strains failed to degrade BBR. Degradation of BBR by J1−8 in LB: Experiments with single (A) and multiple (C) degradations. Degradation of BBR by J1−8 in soil sterilized water: Experiments with single (B) and multiple (D) degradations. Biological replicates were conducted for three times.
3.4. Whole-Genome Alignment Identifies an Essential N-methyltransferase from Methylotenera
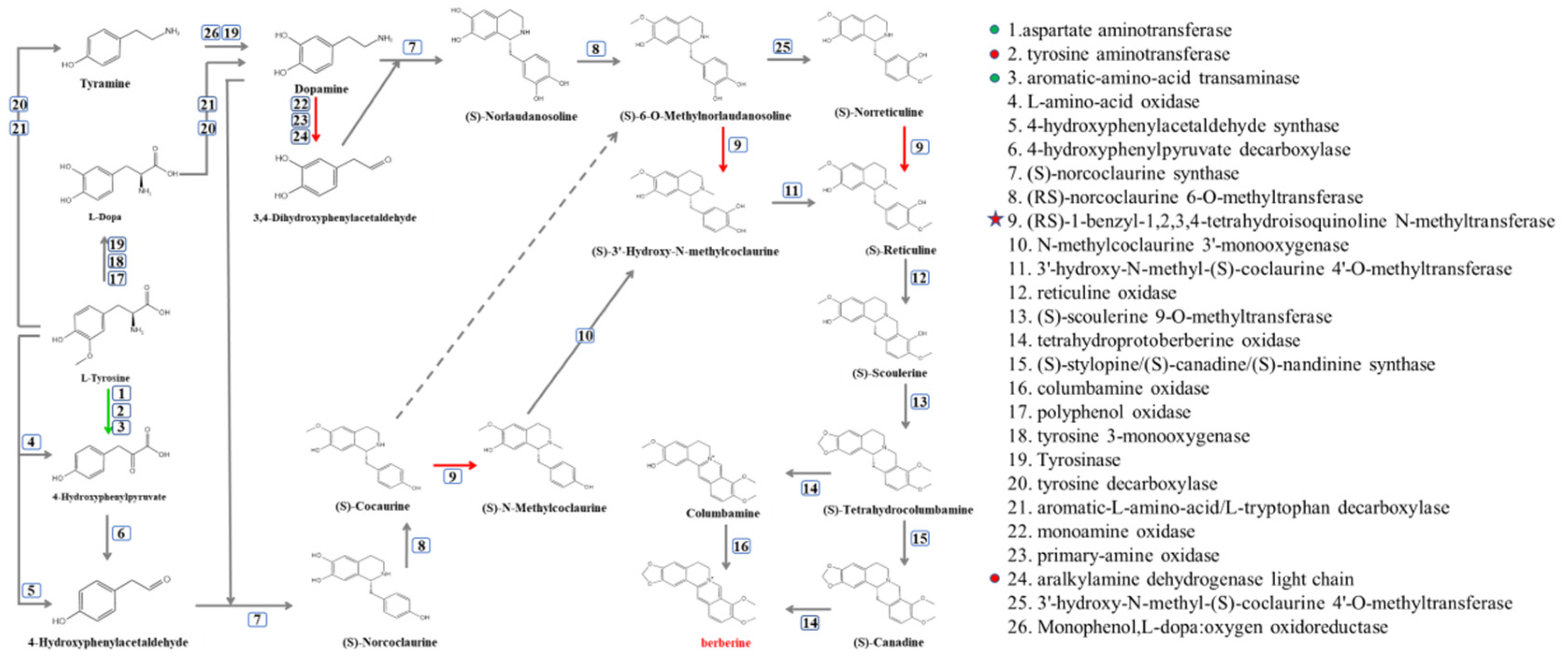
Although BBR was degraded in several soil water samples, the initiation ofthe degradation of BBR was different. However, the variations in the bacterial contents of Methylotenera, Novosphingobium, Delftia, sphingopyxis and Flavobacterium were similar in these soil water samples. Therefore, synthetic pathways of BBR and its related enzymes were obtained from KEGG (Figure 5), and the protein sequences of related enzymes were aligned with the genomes of Methylotenera, Novosphingobium, Delftia, sphingopyxis and Flavobacterium, the contents of which were increased in BBR’s degradation in soil water. The results indicated that orthologs of enzymes 1, 2, 3, 9 and 24 can be found in these bacteria (protein sequence similarity >30%).
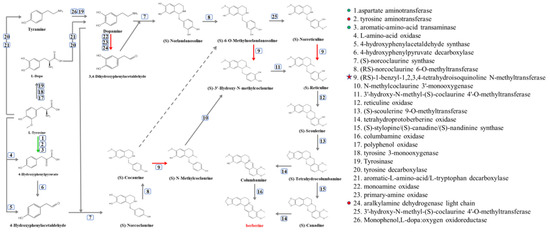
Figure 5.
Synthetic pathway of BBR. Green arrows represent the presence of the enzyme in bacteria with increased bacterial content and bacteria isolated from soil water. The red arrow indicates that the enzyme is only present in bacteria with increased bacterial content.
At the same time, these protein sequences were also aligned to the isolated bacteria from soil water (J1−8; these bacteria could not degrade BBR). The results showed that these bacteria contained orthologs or isoenzymes of enzyme 1, 2, 3 and 24.
Surprisingly, the ortholog of enzyme 9 can only be found in Methylotenera. Therefore, we can deduce that this enzyme is an essential enzyme in the degradation of BBR, which catalyzes three different reactions and existed solely in Methylotenera [20].
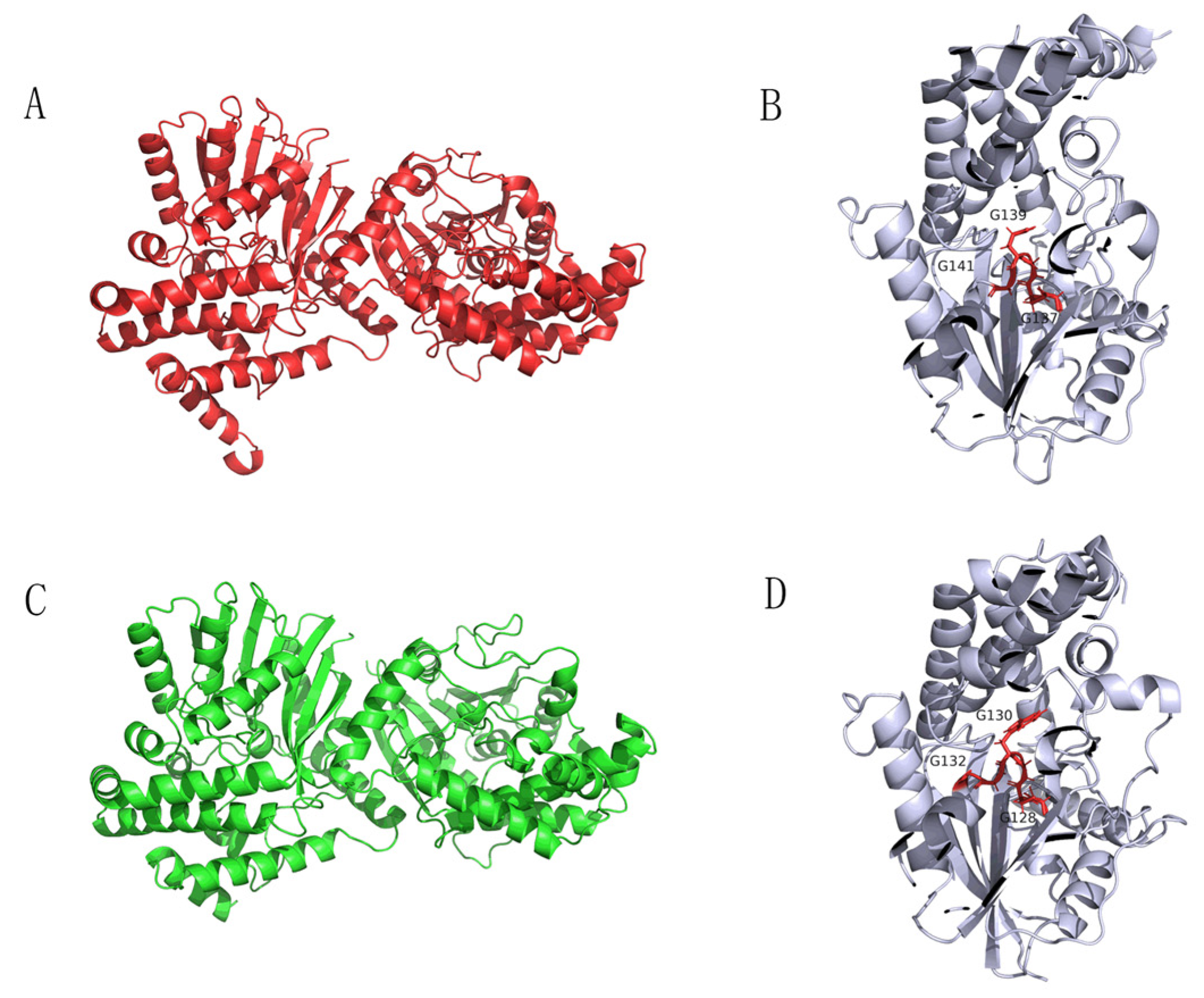
The sequence alignment of Tetrahydroisoquinoline N-methyltransferase (Ortholog of enzyme 9) from Methylotenera indicated a conserved region of GCGXG (Figure 6) [20]. The results are consistent with the study by Torres et al., who found that GXGXG is a highly conserved region [20]. More importantly, this conserved region is located at the active site (Figure 6) [20,21]. In addition, the protein structures and active site of the enzymes from plant and Methylotenera were similar based on the structures generated in silico (Total RMSD value of 5.03, Figure 7). Therefore, Tetrahydroisoquinoline N-methyltransferase from Methylotenera might have an essential role in the degradation of BBR.

Figure 6.
Conserved sequences identified by multiple gene sequence alignments. Alignment of Tetrahydroisoquinoline N-methyltransferase (from the Coptis japonica and from Methylotenera) with its similar multiple sequences. Sequences highlighted in red indicate conserved regions. The abbreviations are as follows: A: (RS)-1-benzyl-1,2,3,4-tetrahydroisoquinoline N-methyltransferase, ag:BAB71802 K13384; B: Crystal structure of Coclaurine N-Methyltransferase, PDB: 6gkv; C: Crystal structure of Coclaurine N-Methyltransferase, PDB: 6gkz; D: Pavine N-methyltransferase in complex with Tetrahydropapaverine and S-adenosylhomocysteine, PDB: 5kok; E: Pavine N-methyltransferase H206A mutant in complex with S-adenosylmethionine, PDB: 5kpc; F: Tetrahydroprotoberberine N-methyltransferase in complex with S-adenosylmethionine, PDB: 6p3n; G: cyclopropane-fatty-acyl-phospholipid synthase family protein, WP_013148163.1.
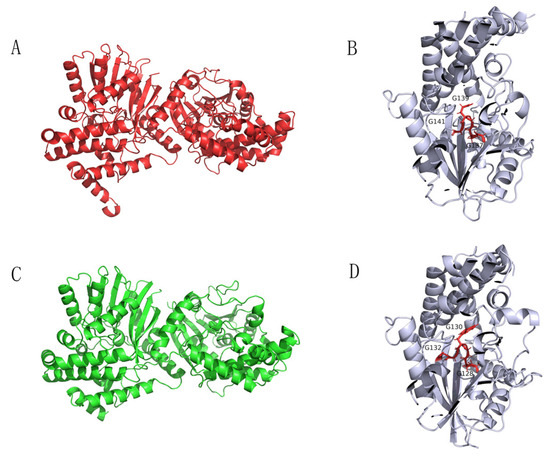
Figure 7.
Protein structure diagrams. The proteins were visualized in ribbon representation. Dimeric structure (A) and its active site (B) of Tetrahydroisoquinoline N-methyltransferase from Coptis japonica. Dimeric structure of Tetrahydroisoquinoline N-methyltransferase in Methylotenera (C) and its active site (D).
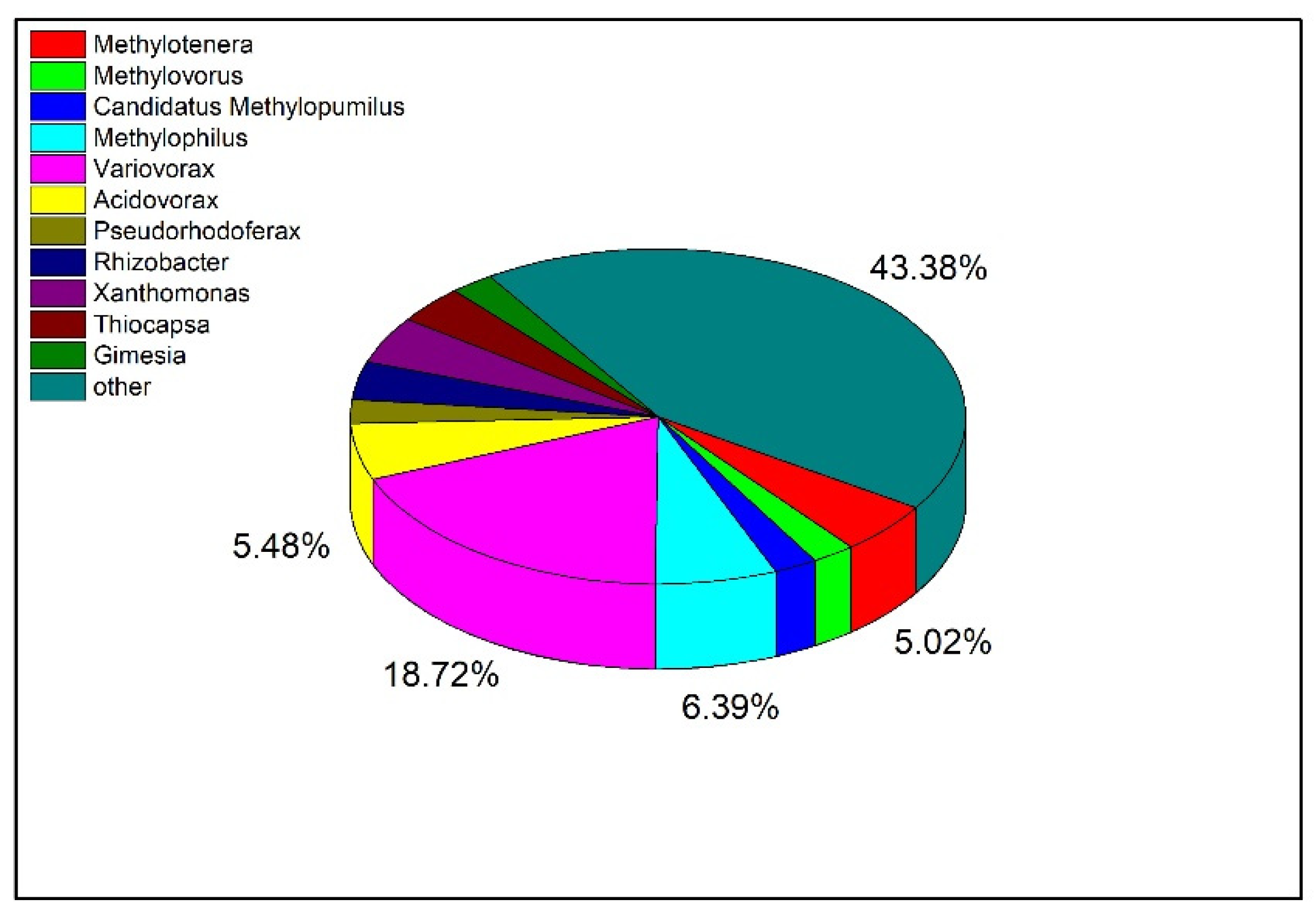
To explore the distribution of Tetrahydroisoquinoline N-methyltransferase in bacteria at the genus level, the protein sequence of the enzyme was blasted and visualized (Figure 8). The top 500 bacteria were selected for analysis according to the sequence similarity, and there were 219 bacteria with similar sequences among the bacteria at the genus level. Among them, there were 11 strains of Methylotenera, accounting for 5.02% of the total. In addition, the largest proportion of orthologs was also identified in Variovorax, which have been reported to regulate the homeostasis of plant hormones and restore contaminated soil and water [22,23].
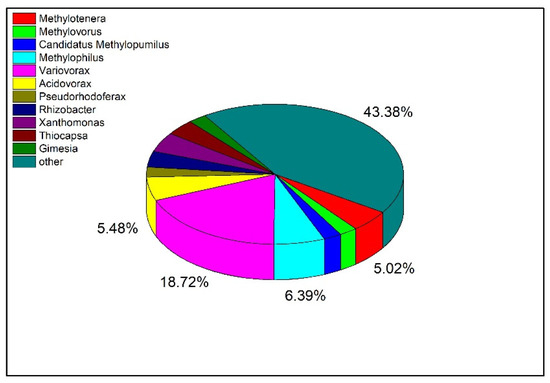
Figure 8.
Distribution of Tetrahydroisoquinoline N-methyltransferase in Different Bacteria.
4. Discussion
In this work, the degradations of BBR in water from different sources and in soil water from different regions are investigated. The 16s rDNA amplicon sequencing is carried out, and the relationship between BBR’s degradation and bacterial diversity is explored. In addition, the protein sequences of enzymes in the synthesis pathway of BBR are aligned with the whole genome of related bacteria. We demonstrated that Tetrahydroisoquinoline N-methyltransferase, which catalyzes three reactions in the BBR synthesis pathway, was discovered in Methylotenera, the content of which had positive relationships with BBR’s degradation. As a highly conserved protein, Tetrahydroisoquinoline N-methyltransferase is deduced to be an essential enzyme in the degradation of BBR by bacteria in soil water.
The degradations of BBR were distinct in soil water collected from different places, while the bacterial contents of Methylotenera, Novosphingobium, Delftia, Sphingopyxis and Flavobacterium were all increased in the samples in which BBR was degraded. Due to the long synthetic pathway of BBR in plants, one bacterium is unlikely to contain all the orthologs that can be used for the degradation of BBR. Therefore, the bacterial degradation of BBR is mainly accomplished by the bacterial community. However, BBR is degraded in only three soil water samples, and it is stable in the other ones within the 14-day cultivation. More importantly, once BBR is initiatedfor degradation, it will be consumed rapidly within two days. These results provide strong evidence that there exist one or more degradation-limiting steps in bacteria community. Even though several bacteria are isolated and grown in the artificial medium, BBR’s degradation is not observed when cultivated with these strains. Methylotenera might be a genus that is unable to grow in the artificial medium, but our data suggested that it is an important element in the degradation of BBR in soil water because of its Tetrahydroisoquinoline N-methyltransferase.
Another aim of our work is to answer whether the application of BBR is beneficial to the soil bacterial diversity or not. Among the strains with increased bacterial contents under the treatment of BBR, most of them are beneficial to the environment except Flavobacterium. Among them, Methylotenera exists belongs to Methylophilaceae. Methylophilaceae consists of two species, Methylotenera mobilis and Methylotenera versatilis, both of which were isolated from Lake Washington sediments [24]. The study reported that Methylotenera mobilis is an obligate methylamine utilizer. Methylotenera versatilis does not possess the hallmark methanol dehydrogenase of Methylophilaceae, but it still showed weak growth on solid agar medium supplemented with methanol [24,25]. It has been confirmed that Methylotenera can be used as a degradation agent for environmental pollutants such as petroleum, motor oil and methane [26,27,28]. In addition, Novosphingobium is an excellent phenanthrene degrader [29]. Delftia can biodegrade organic pollutants, such as phenolic compounds and chlorobenzene [30]. Sphingopyxis can efficiently degrade the aromatic compounds, such as microcystins, tetralin, styrene and triclosan [31]. These data demonstrate that BBR’s degradation enriched beneficial bacteria in the environment.
N-methyltransferases participate in the biosynthesis of the monoterpene indole alkaloids vindoline and ajmaline. Based on the widespread presence of a large number of N-methylated and N,N-dimethylated compounds, N-methyltransferases play an important role in Benzylisoquinoline alkaloids metabolism [32]. In particular, Tetrahydroprotoberberine N-methyltransferase exhibits specificity for certain intermediates in the tetrahydroprotoberberine branch pathway. Tetrahydroprotoberberine N-methyltransferase is the only AdoMet-dependent N-methyltransferase in plant alkaloid metabolism [33]. Here we identify an ortholog from bacteria and suggest its importance in biodegradation of BBR. As an enzyme that catalyzes this rate-limiting process, Tetrahydroprotoberberine N-methyltransferase might be a promising enzyme in soil bioremediation.
In summary, we demonstrated the essential factors in BBR’s degradation in soil water, providing evidence of its effects on the environment. Future work will be concentrated on analyzing this important enzyme to provide it as a soil bioremediation agent.
5. Conclusions
Extended applications of BBR pose potential risks to the environment. Herein, we investigated BBR’s degradation in water. We confirmed that BBR was only degraded in soil water, while it was stable in tap water, river water and aquaculture water. The degradation of BBR in soil water was closely related to the enrichment of Methylotenera. Interestingly, an ortholog of Tetrahydroisoquinoline N-methyltransferase from plant was discovered only in Methylotenera and catalyzed a crucial step in BBR’s degradation pathway.
Author Contributions
R.H. and X.G. designed the research. R.H. and Y.C. conducted the experiments and data analysis. R.H. and Y.L. wrote the manuscript and X.G. revised it. R.H. submitted the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Major project for the transformation of scientific and technological achievements in Hebei Province [19026517Z]; Beijing Municipal Commission of Education [KZ201911417049].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the compounds are available from the authors via email request.
References
- Chen, L.; Zhu, L.; Chen, J. Crystal structure-guided design of berberine-based novel chitinase inhibitors. J. Enzym. Inhib. Med. Chem. 2020, 35, 1937–1943. [Google Scholar] [CrossRef] [PubMed]
- Mujtaba, M.A.; Akhter, M.H.; Alam, M.S. An Updated Review on Therapeutic Potential and Recent Advances in Drug Delivery of Berberine: Current Status and Future Prospect. Curr. Pharm. Biotechnol. 2022, 23, 60–71. [Google Scholar] [CrossRef]
- Othman, M.S.; Safwat, G.; Aboulkhair, M.; Moneim, A.E.A. The potential effect of berberine in mercury-induced hepatorenal toxicity in albino rats. Food Chem. Toxicol. 2014, 69, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Shao, W.; Tian, A.S. Berberine alleviates myocardial ischemia–reperfusion injury by inhibiting inflammatory response and oxidative stress: The key function of miR-26b-5p-mediated PTGS2/MAPK signal transduction. Pharm. Biol. 2022, 60, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.J.-J. Herbicidal Spectrum, Absorption and Transportation, and Physiological Effect on Bidens pilosa of the Natural Alkaloid Berberine. Bidens Pilosa Nat. 2017, 65, 6100–6113. [Google Scholar]
- Ying, L.Y.-M. Evaluation of berberine as a natural fungicide: Biodegradation and antimicrobial mechanism. J. Asian Nat. Prod. Res. 2017, 20, 148–162. [Google Scholar]
- Fu, W.; Tian, G.; Pei, Q.; Ge, X.; Tian, P. Evaluation of berberine as a natural compound to inhibit peach brown rot pathogen Monilinia fructicola. Crop Prot. 2017, 91, 20–26. [Google Scholar] [CrossRef]
- Qin, W.; Song, Y.; Dai, Y.; Qiu, G.; Ren, M.; Zeng, P. Treatment of berberine hydrochloride pharmaceutical wastewater by O3/UV/H2O2 advanced oxidation process. Environ. Earth Sci. 2015, 73, 4939–4946. [Google Scholar] [CrossRef]
- Thakur, M.P.; Phillips, H.; Brose, U. Towards an integrative understanding of soil biodiversity. Biol. Rev. 2020, 95, 350–364. [Google Scholar] [CrossRef]
- Tudi, M.; Ruan, H.D.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D. Agriculture Development, Pesticide Application and Its Impact on the Environment. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.; Zeng, P.; Wang, H.; Song, Y.; Li, J. Isolation and Characterization of a Bacterial Strain Capable of Efficient Berberine Degradation. Environ. Res. Public Health 2019, 16, 646. [Google Scholar] [CrossRef]
- Yoshida, H.; Takeda, H.; Wakana, D. Characterization of Burkholderia sp. strain CJ1, a newly isolated berberine-degrading bacterium from rhizosphere of Coptis japonica. Biosci. Biotechnol. Biochem. 2020, 84, 1299–1302. [Google Scholar] [CrossRef]
- Takeda, H.; Ishikawa, K.; Wakana, D. 11-Hydroxylation of protoberberine by the novel berberine-utilizing aerobic bacterium Sphingobium sp. strain BD3100. J. Nat. Prod. 2015, 78, 2880–2886. [Google Scholar] [CrossRef]
- Qiu, G.; Song, Y.-H.; Zeng, P.; Duan, L.; Xiao, S. Characterization of bacterial communities in hybrid upflow anaerobic sludge blanket (UASB)–membrane bioreactor (MBR) process for berberine antibiotic wastewater treatment. Bioresour. Technol. 2013, 142, 52–62. [Google Scholar] [CrossRef]
- Delorenzo, M.E.R. Toxicity of pesticides to aquatic microorganisms: A review. Environ. Toxicol. Chem. 2001, 20, 84–98. [Google Scholar] [CrossRef]
- Boricha, H.; Fulekar, M.H.J.R.B.L. Identification of Owenweeksia honkongenesis as a novel organism for the remediation of pesticide-fenvalerate. Rom. Biotechnol. Lett. 2010, 15, 5104–5110. [Google Scholar]
- Han, L.; Fang, K.; Liu, Y.; Fang, J.; Wang, F.; Wang, X. Earthworms accelerated the degradation of the highly toxic acetochlor S-enantiomer by stimulating soil microbiota in repeatedly treated soils. J. Hazard. Mater. 2021, 420, 126669. [Google Scholar] [CrossRef]
- Purahong, K.P. Influence of Different Forest System Management Practices on Leaf Litter Decomposition Rates, Nutrient Dynamics and the Activity of Ligninolytic Enzymes: A Case Study from Central European Forests. PLoS ONE 2014, 9, e93700. [Google Scholar]
- Strickland, M.S.; Lauber, C.; Fierer, N.; Bradford, M. Testing the functional significance of microbial community composition. Ecology 2009, 90, 441–451. [Google Scholar] [CrossRef]
- Torres, M.A.; Hoffarth, E.; Eugenio, L.; Savtchouk, J.; Chen, X.; Morris, J.S.; Facchini, P.J.; Ng, K.K.-S. Structural and Functional Studies of Pavine N-Methyltransferase from Thalictrum flavum Reveal Novel Insights into Substrate Recognition and Catalytic Mechanism. J. Biol. Chem. 2016, 291, 23403–23415. [Google Scholar] [CrossRef]
- Lang, D.E.; Morris, J.S.; Rowley, M.; Torres, M.A.; Maksimovich, V.A.; Facchini, P.J.; Ng, K.K. Structure–function studies of tetrahydroprotoberberine N -methyltransferase reveal the molecular basis of stereoselective substrate recognition. J. Biol. Chem. 2019, 294, 11482–14498. [Google Scholar] [CrossRef] [PubMed]
- Rgta, C.; Aab, B.; Icd, A.J.N.B. Microbial inoculum development for ameliorating crop drought stress: A case study of Variovorax paradoxus 5C-2. New Biotechnol. 2020, 56, 103–113. [Google Scholar]
- Sorensen, S.R.; Albers, C.N.; Aamand, J. Rapid Mineralization of the Phenylurea Herbicide Diuron by Variovorax sp. Strain SRS16 in Pure Culture and within a Two-Member Consortium. Appl. Environ. Microbiol. 2008, 74, 2332–2340. [Google Scholar] [CrossRef] [PubMed]
- Kalyuzhnaya, M.; Bowerman, S.; Lara, J.C.; Lidstrom, M.E.; Chistoserdova, L. an obligately methylamine-utilizing bacterium within the family Methylophilaceae. Int J Syst Evol Microbiol. Int. J. Syst. Evol. Microbiol. 2007, 56, 2819–2823. [Google Scholar] [CrossRef]
- Beck, D.A.C.; Hendrickson, E.L.; Vorobev, A.; Wang, T.; Lim, S.; Kalyuzhnaya, M.G.; Lidstrom, M.E.; Hackett, M.; Chistoserdova, L. An Integrated Proteomics/Transcriptomics Approach Points to Oxygen as the Main Electron Sink for Methanol Metabolism in Methylotenera mobilis. J. Bacteriol. 2011, 193, 4758–4765. [Google Scholar] [CrossRef] [Green Version]
- Idomeh, J.E.; Shittu, O.B.; Oyedepo, J.A.; Bada, B.S.; Balogun, S.A.; Idomeh, F.A.; Ezenweani, R.S. Petroleum Hydrocarbon Impacted Aquatic Ecosystem Reveals Methylotenera as the Dominant Genera in the Niger Delta Region of Nigeria. Geomicrobiol. J. 2021, 38, 879–894. [Google Scholar] [CrossRef]
- Wang, M.; Deng, B.; Fu, X.; Sun, H.; Xu, Z. Characterizations of microbial diversity and machine oil degrading microbes in machine oil contaminated soil. Environ. Pollut. 2019, 255, 113190. [Google Scholar] [CrossRef]
- Grinsven, S.V.; Damsté, J.S.S.; Harrison, J. Nitrate promotes the transfer of methane-derived carbon from the methanotroph Methylobacter sp. to the methylotroph Methylotenera sp. in eutrophic lake water. Limonlogy Oceanogr. 2020, 66, 878–891. [Google Scholar] [CrossRef]
- Segura, A.; Udaondo, Z.; Microbiology, L.M.J.E. PahT regulates carbon fluxes in Novosphingobium sp. HR1a and influences its survival in soil and rhizospheres. Environ. Microbiol. 2021, 23, 2969–2991. [Google Scholar] [CrossRef]
- Cong, C.; Zhou, W.; Xu, D. Genomic Analysis of Delftia tsuruhatensis Strain TR1180 Isolated From A Patient From China With In4-Like Integron-Associated Antimicrobial Resistance. Front. Cell Infect. Microbiol. 2021, 11, 663933. [Google Scholar]
- Yang, F.; Feng, H.; Massey, I.Y.; Huang, F.; Guo, J.; Zhang, X. Genome-Wide Analysis Reveals Genetic Potential for Aromatic Compounds Biodegradation of Sphingopyxis. BioMed Res. Int. 2020, 2020, 5849123. [Google Scholar] [CrossRef] [PubMed]
- Liscombe, D.K.; Ziegler, J.; Schmidt, J. Targeted metabolite and transcript profiling for elucidating enzyme function: Isolation of novel N-methyltransferases from three benzylisoquinoline alkaloid-producing species. Plant J. 2009, 60, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Liscombe, D.K.; Facchini, P.J. Molecular cloning and characterization of tetrahydroprotoberberine cis-N-methyltransferase, an enzyme involved in alkaloid biosynthesis in opium poppy. J. Biol. Chem. 2007, 282, 14741. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).