
Wistin Exerts an Anti-Inflammatory Effect via Nuclear Factor-κB and p38 Signaling Pathways in Lipopolysaccharide-Stimulated RAW264.7 Cells


 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Effects of Wistin on Cell Viability in LPS-Induced RAW 264.7 Cells
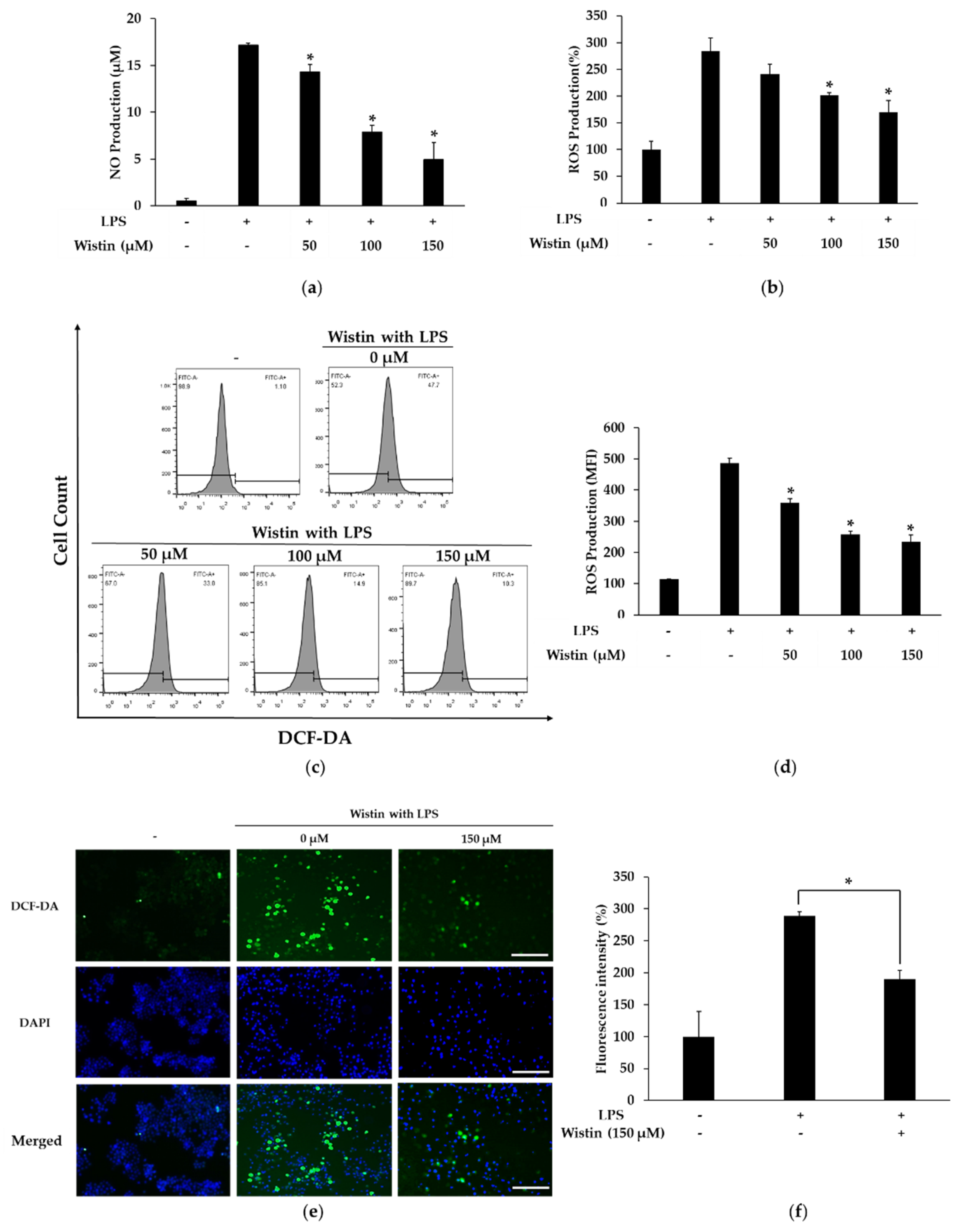
2.2. Effects of Wistin on the Production of Pro-Inflammatory Mediators in LPS-Induced RAW 264.7 Cells
2.3. Effects of Wistin on Pro-Inflammatory Enzymes and Cytokine Gene Expression in LPS-Induced RAW 264.7 Cells
2.4. Effects of Wistin on the Protein Expression Level of Pro-Inflammatory Enzymes in LPS-Induced RAW 264.7 Cells
2.5. Effects of Wistin on the Activation of AKT/NF-κB Pathway in LPS-Induced RAW 264.7 Cells
2.6. Effects of Wistin on the Activation of MAPK Pathway in LPS-Induced RAW 264.7 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Cell Viability
4.4. Measurement of NO Production
4.5. Measurement of ROS Production
4.6. QRT-PCR
4.7. Western Blot Analysis
4.8. Immunofluorescence
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, N.; Baby, D.; Rajguru, J.P.; Patil, P.B.; Thakkannavar, S.S.; Pujari, V.B. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Narumiya, S. Prostaglandin-cytokine crosstalk in chronic inflammation. Br. J. Pharmacol. 2019, 176, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Shi, G. Editorial: Regulation of Inflammation in Chronic Disease. Front. Immunol. 2019, 10, 737. [Google Scholar] [CrossRef]
- Panigrahy, D.; Gilligan, M.M.; Serhan, C.N.; Kashfi, K. Resolution of inflammation: An organizing principle in biology and medicine. Pharmacol. Ther. 2021, 227, 107879. [Google Scholar] [CrossRef]
- Kozuch, P.L.; Hanauer, S.B. Treatment of inflammatory bowel disease: A review of medical therapy. World J. Gastroenterol. 2008, 14, 354–377. [Google Scholar] [CrossRef]
- PLoSker, G.L.; Croom, K.F. Sulfasalazine: A review of its use in the management of rheumatoid arthritis. Drugs 2005, 65, 1825–1849. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Gonzalez-Rey, M.; Bebianno, M.J. Non-steroidal anti-inflammatory drug (NSAID) ibuprofen distresses antioxidant defense system in mussel Mytilus galloprovincialis gills. Aquat. Toxicol. 2011, 105, 264–269. [Google Scholar] [CrossRef]
- Parolini, M. Toxicity of the Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) acetylsalicylic acid, paracetamol, diclofenac, ibuprofen and naproxen towards freshwater invertebrates: A review. Sci. Total Environ. 2020, 740, 140043. [Google Scholar] [CrossRef]
- Rainsford, K.D. Ibuprofen: Pharmacology, efficacy and safety. Inflammopharmacology 2009, 17, 275–342. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Kang, R.; Coyne, C.B.; Zeh, H.J.; Lotze, M.T. PAMPs and DAMPs: Signal 0s that spur autophagy and immunity. Immunol. Rev. 2012, 249, 158–175. [Google Scholar] [CrossRef]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. TLR signaling. Semin. Immunol. 2007, 19, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Shan, C.; Ning, D. Walnut oil alleviates LPS-induced intestinal epithelial cells injury by inhibiting TLR4/MyD88/NF-kappaB pathway activation. J. Food Biochem. 2021, 45, e13955. [Google Scholar] [CrossRef]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy--from molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta 2005, 1754, 253–262. [Google Scholar] [CrossRef]
- Sharif, O.; Bolshakov, V.N.; Raines, S.; Newham, P.; Perkins, N.D. Transcriptional profiling of the LPS induced NF-kappaB response in macrophages. BMC Immunol. 2007, 8, 1. [Google Scholar] [CrossRef]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-kappaB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef]
- Bai, D.; Ueno, L.; Vogt, P.K. Akt-mediated regulation of NFkappaB and the essentialness of NFkappaB for the oncogenicity of PI3K and Akt. Int. J. Cancer 2009, 125, 2863–2870. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.S. Phytochemical Constituents of Medicinal Plants for the Treatment of Chronic Inflammation. Biomolecules 2021, 11, 672. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.L.; Brockmueller, A.; Kunnumakkara, A.B.; Shakibaei, M. Modulation of Inflammation by Plant-Derived Nutraceuticals in Tendinitis. Nutrients 2022, 14, 30. [Google Scholar] [CrossRef] [PubMed]
- Vonkeman, H.E.; van de Laar, M.A. Nonsteroidal anti-inflammatory drugs: Adverse effects and their prevention. Semin. Arthritis. Rheum. 2010, 39, 294–312. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Doan, H.Q.; Brunell, D.J.; Rady, P.; Tyring, S.K. Apoptotic gene expression in sinecatechins-treated external genital and perianal warts. Viral Immunol. 2014, 27, 556–558. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.V.; Fernandes, T.A.A.d.M.; de Azevedo, J.C.V.; Cobucci, R.N.O.; Cobucci, R.N.; de Carvalho, M.G.F.; Andrade, V.S.; de Araújo, J.M.G. Link between chronic inflammation and human papillomavirus-induced carcinogenesis (Review). Oncol. Lett. 2015, 9, 1015–1026. [Google Scholar] [CrossRef]
- Lovelace, E.S.; Wagoner, J.; MacDonald, J.; Bammler, T.; Bruckner, J.; Brownell, J.; Beyer, R.P.; Zink, E.M.; Kim, Y.M.; Kyle, J.E.; et al. Silymarin Suppresses Cellular Inflammation By Inducing Reparative Stress Signaling. J. Nat. Prod. 2015, 78, 1990–2000. [Google Scholar] [CrossRef]
- Fried, M.W.; Navarro, V.J.; Afdhal, N.; Belle, S.H.; Wahed, A.S.; Hawke, R.L.; Doo, E.; Meyers, C.M.; Reddy, K.R.; Silymarinin, N.; et al. Effect of silymarin (milk thistle) on liver disease in patients with chronic hepatitis C unsuccessfully treated with interferon therapy: A randomized controlled trial. JAMA 2012, 308, 274–282. [Google Scholar] [CrossRef]
- Mayer, K.E.; Myers, R.P.; Lee, S.S. Silymarin treatment of viral hepatitis: A systematic review. J. Viral Hepat. 2005, 12, 559–567. [Google Scholar] [CrossRef]
- Ryoo, S.B.; Oh, H.K.; Yu, S.A.; Moon, S.H.; Choe, E.K.; Oh, T.Y.; Park, K.J. The effects of eupatilin (stillen(R)) on motility of human lower gastrointestinal tracts. Korean J. Physiol. Pharmacol. 2014, 18, 383–390. [Google Scholar] [CrossRef]
- Saleh, H.A.; Yousef, M.H.; Abdelnaser, A. The Anti-Inflammatory Properties of Phytochemicals and Their Effects on Epigenetic Mechanisms Involved in TLR4/NF-kappaB-Mediated Inflammation. Front. Immunol. 2021, 12, 606069. [Google Scholar] [CrossRef]
- Goh, Y.X.; Jalil, J.; Lam, K.W.; Husain, K.; Premakumar, C.M. Genistein: A Review on its Anti-Inflammatory Properties. Front. Pharmacol. 2022, 13, 820969. [Google Scholar] [CrossRef]
- Yu, J.; Bi, X.; Yu, B.; Chen, D. Isoflavones: Anti-Inflammatory Benefit and Possible Caveats. Nutrients 2016, 8, 361. [Google Scholar] [CrossRef] [PubMed]
- Sanada, M.; Hayashi, R.; Imai, Y.; Nakamura, F.; Inoue, T.; Ohta, S.; Kawachi, H. 4′,6-dimethoxyisoflavone-7-O-beta-D-glucopyranoside (wistin) is a peroxisome proliferator-activated receptor gamma (PPARgamma) agonist that stimulates adipocyte differentiation. Anim. Sci. J. 2016, 87, 1347–1351. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Nakamura, F.; Taguchi, E.; Nakata, M.; Wada, F.; Takihi, M.; Inoue, T.; Ohta, S.; Kawachi, H. 4′,6-Dimethoxyisoflavone-7-O-beta-D-glucopyranoside (wistin) is a peroxisome proliferator-activated receptor alpha (PPARalpha) agonist in mouse hepatocytes. Mol. Cell Biochem. 2018, 446, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kazlauskaite, J.A.; Kopustinskiene, D.M. Pleiotropic Effects of Isoflavones in Inflammation and Chronic Degenerative Diseases. Int. J. Mol. Sci. 2021, 22, 5656. [Google Scholar] [CrossRef] [PubMed]
- Agita, A.; Alsagaff, M.T. Inflammation, Immunity, and Hypertension. Acta Med. Indones. 2017, 49, 158–165. [Google Scholar] [PubMed]
- Ma, Y.; Tang, T.; Sheng, L.; Wang, Z.; Tao, H.; Zhang, Q.; Zhang, Y.; Qi, Z. Aloin suppresses lipopolysaccharideinduced inflammation by inhibiting JAK1STAT1/3 activation and ROS production in RAW264.7 cells. Int. J. Mol. Med. 2018, 42, 1925–1934. [Google Scholar] [CrossRef]
- Park, J.Y.; Chung, T.W.; Jeong, Y.J.; Kwak, C.H.; Ha, S.H.; Kwon, K.M.; Abekura, F.; Cho, S.H.; Lee, Y.C.; Ha, K.T.; et al. Ascofuranone inhibits lipopolysaccharide-induced inflammatory response via NF-kappaB and AP-1, p-ERK, TNF-alpha, IL-6 and IL-1beta in RAW 264.7 macrophages. PLoS ONE 2017, 12, e0171322. [Google Scholar] [CrossRef]
- Murakami, A.; Ohigashi, H. Targeting NOX, INOS and COX-2 in inflammatory cells: Chemoprevention using food phytochemicals. Int. J. Cancer 2007, 121, 2357–2363. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-kappaB pathway for the therapy of diseases: Mechanism and clinical study. Signal. Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Thalhamer, T.; McGrath, M.A.; Harnett, M.M. MAPKs and their relevance to arthritis and inflammation. Rheumatology (Oxford) 2008, 47, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Lin, M.W.; Liang, C.J.; Wang, S.H. The Anti-Inflammatory Effects and Mechanisms of Eupafolin in Lipopolysaccharide-Induced Inflammatory Responses in RAW264.7 Macrophages. PLoS ONE 2016, 11, e0158662. [Google Scholar] [CrossRef] [PubMed]
- Adedapo, A.; Adewuyi, T.; Sofidiya, M. Phytochemistry, anti-inflammatory and analgesic activities of the aqueous leaf extract of Lagenaria breviflora (Cucurbitaceae) in laboratory animals. Rev. Biol. Trop. 2013, 61, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Song, E.H.; Chung, K.S.; Kang, Y.M.; Lee, J.H.; Lee, M.; An, H.J. Eupatilin suppresses the allergic inflammatory response in vitro and in vivo. Phytomedicine 2018, 42, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hartog, A.; Hougee, S.; Faber, J.; Sanders, A.; Zuurman, C.; Smit, H.F.; van der Kraan, P.M.; Hoijer, M.A.; Garssen, J. The multicomponent phytopharmaceutical SKI306X inhibits in vitro cartilage degradation and the production of inflammatory mediators. Phytomedicine 2008, 15, 313–320. [Google Scholar] [CrossRef]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar] [PubMed]
- Checa, J.; Aran, J.M. Reactive Oxygen Species: Drivers of Physiological and Pathological Processes. J. Inflamm. Res. 2020, 13, 1057–1073. [Google Scholar] [CrossRef]
- Moita, E.; Gil-Izquierdo, A.; Sousa, C.; Ferreres, F.; Silva, L.R.; Valentao, P.; Dominguez-Perles, R.; Baenas, N.; Andrade, P.B. Integrated analysis of COX-2 and iNOS derived inflammatory mediators in LPS-stimulated RAW macrophages pre-exposed to Echium plantagineum L. bee pollen extract. PLoS ONE 2013, 8, e59131. [Google Scholar] [CrossRef]
- Xiao, K.; Liu, C.; Tu, Z.; Xu, Q.; Chen, S.; Zhang, Y.; Wang, X.; Zhang, J.; Hu, C.A.; Liu, Y. Activation of the NF-kappaB and MAPK Signaling Pathways Contributes to the Inflammatory Responses, but Not Cell Injury, in IPEC-1 Cells Challenged with Hydrogen Peroxide. Oxid. Med. Cell Longev. 2020, 2020, 5803639. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-kappaB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Sun, M.; Liu, Z. Phytochemicals with protective effects against acute pancreatitis: A review of recent literature. Pharm. Biol. 2022, 60, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kauppinen, A.; Kaarniranta, K. Emerging role of NF-kappaB signaling in the induction of senescence-associated secretory phenotype (SASP). Cell Signal. 2012, 24, 835–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Primer | Primer Sequence |
|---|---|
| iNos | Forward 5′-GAACGGAGAACGTTGGATTTG-3′ |
| Reverse 5′-TCAGGTCACTTTGGTAGGATTT-3′ | |
| Cox-2 | Forward 5′-GAAGATTCCCTCCGGTGTTT-3′ |
| Reverse 5′-CCCTTCTCACTGGCTTATGTAG-3′ | |
| Tnf-α | Forward 5′-ACGTGGAACTGGCAGAAGAG-3′ |
| Reverse 5′-GGTCTGGGCCATAGAACTGA-3′ | |
| Il-6 | Forward 5′-TCTGAAGGACTCTGGCTTTG-3′ |
| Reverse 5′-GATGGATGCTACCAAACTGGA-3′ | |
| Il-1β | Forward 5′-AGGTCAAAGGTTTGGAAGCA-3′ |
| Reverse 5′-TGAAGCAGCTATGGCAACTG-3′ | |
| β-actin | Forward 5′-ATGGAGGGGAATACAGCCC-3′ |
| Reverse 5′-TTCTTTGCAGCTCCTTCGTT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, J.; Ryu, G.; Shin, S.-A.; Kim, H.; Ahn, M.-J.; Lee, J.H.; Lee, C.S. Wistin Exerts an Anti-Inflammatory Effect via Nuclear Factor-κB and p38 Signaling Pathways in Lipopolysaccharide-Stimulated RAW264.7 Cells. Molecules 2022, 27, 5719. https://doi.org/10.3390/molecules27175719
An J, Ryu G, Shin S-A, Kim H, Ahn M-J, Lee JH, Lee CS. Wistin Exerts an Anti-Inflammatory Effect via Nuclear Factor-κB and p38 Signaling Pathways in Lipopolysaccharide-Stimulated RAW264.7 Cells. Molecules. 2022; 27(17):5719. https://doi.org/10.3390/molecules27175719
Chicago/Turabian StyleAn, Jangeun, Gyoungah Ryu, Seong-Ah Shin, Huiji Kim, Mi-Jeong Ahn, Jun Hyuck Lee, and Chang Sup Lee. 2022. "Wistin Exerts an Anti-Inflammatory Effect via Nuclear Factor-κB and p38 Signaling Pathways in Lipopolysaccharide-Stimulated RAW264.7 Cells" Molecules 27, no. 17: 5719. https://doi.org/10.3390/molecules27175719
APA StyleAn, J., Ryu, G., Shin, S.-A., Kim, H., Ahn, M.-J., Lee, J. H., & Lee, C. S. (2022). Wistin Exerts an Anti-Inflammatory Effect via Nuclear Factor-κB and p38 Signaling Pathways in Lipopolysaccharide-Stimulated RAW264.7 Cells. Molecules, 27(17), 5719. https://doi.org/10.3390/molecules27175719