
Sweroside Ameliorated Memory Deficits in Scopolamine-Induced Zebrafish (Danio rerio) Model: Involvement of Cholinergic System and Brain Oxidative Stress

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
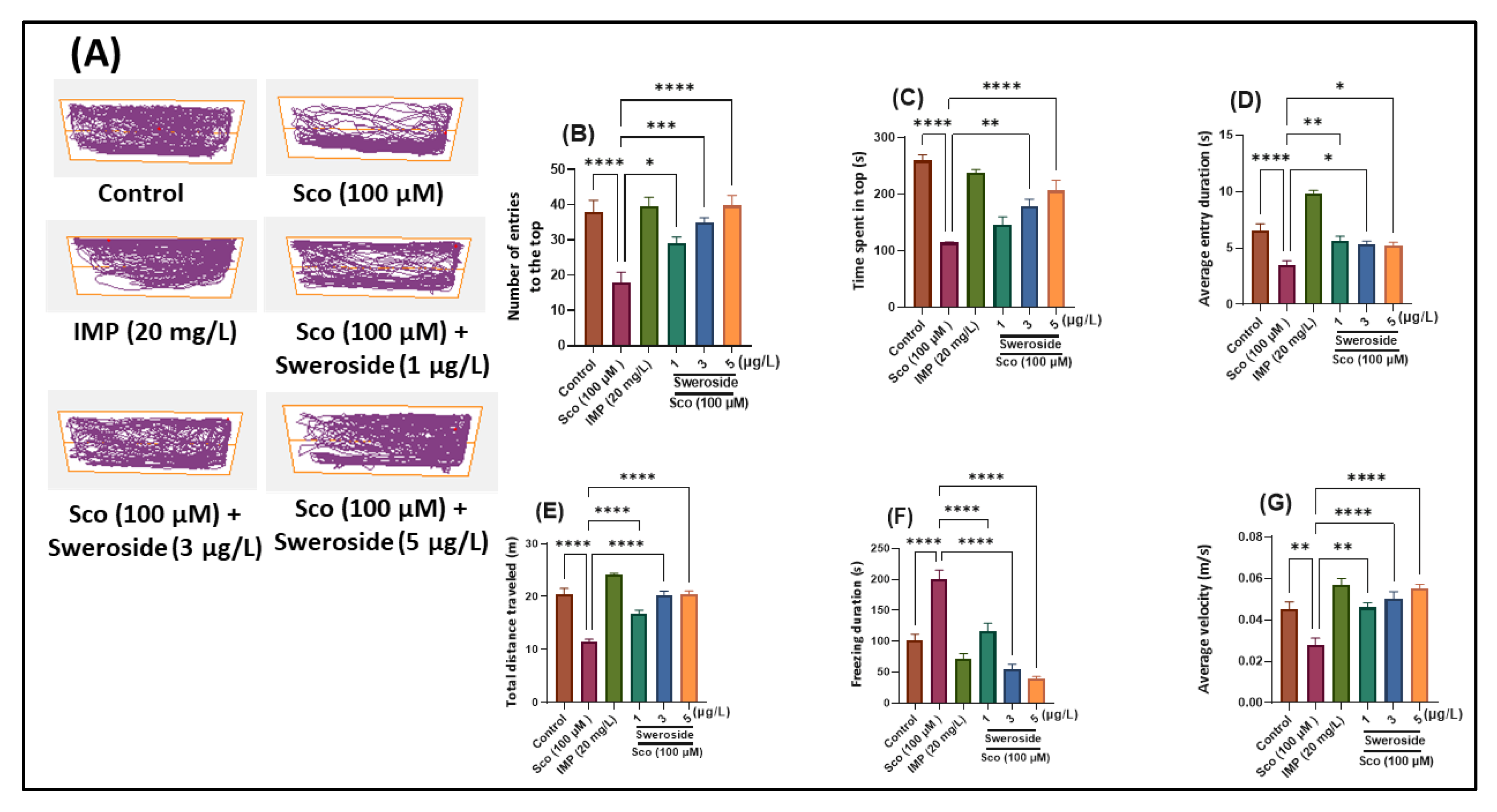
2.1. Effects on Anxiety-Like Behavior in the NTT Test
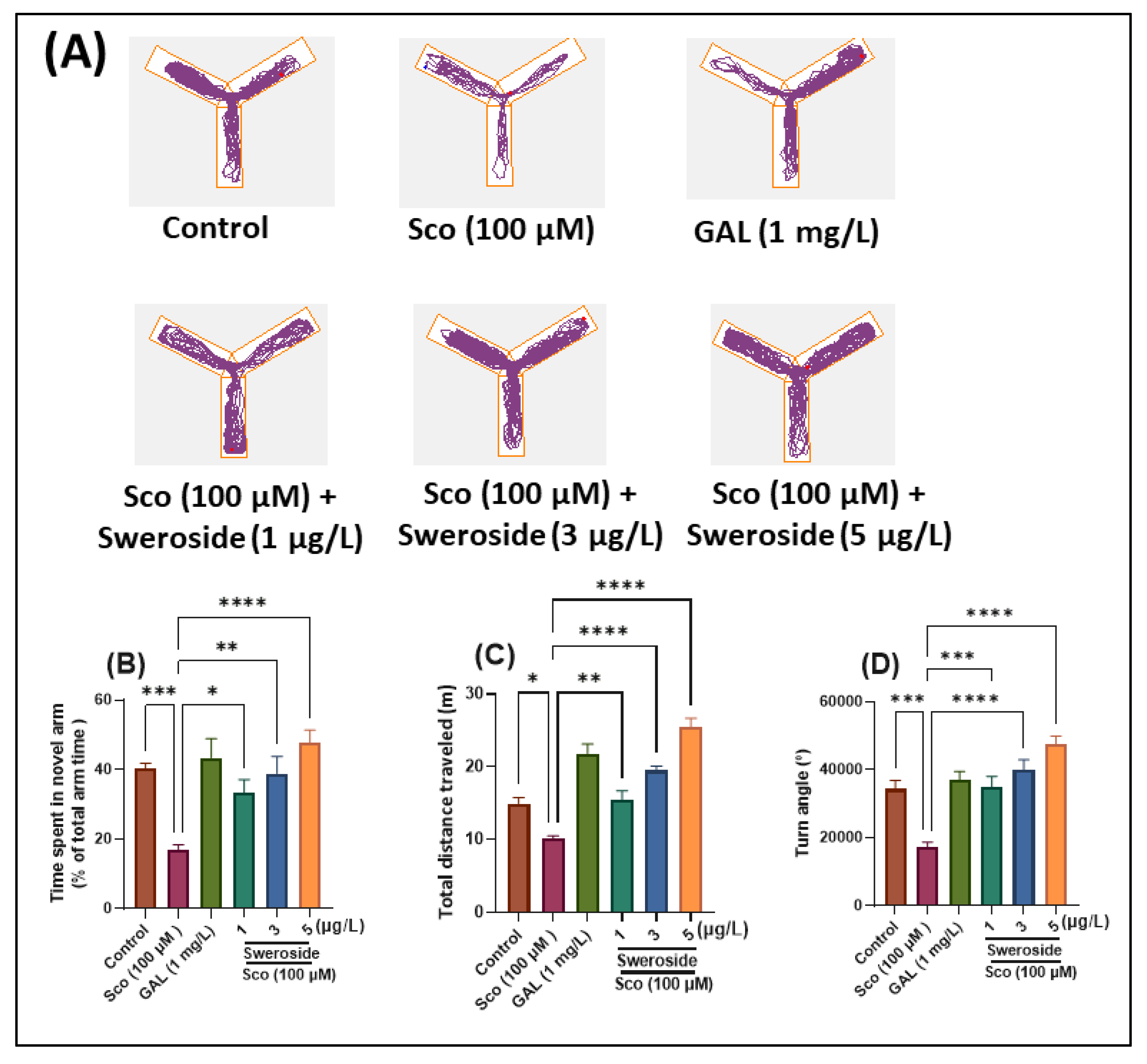
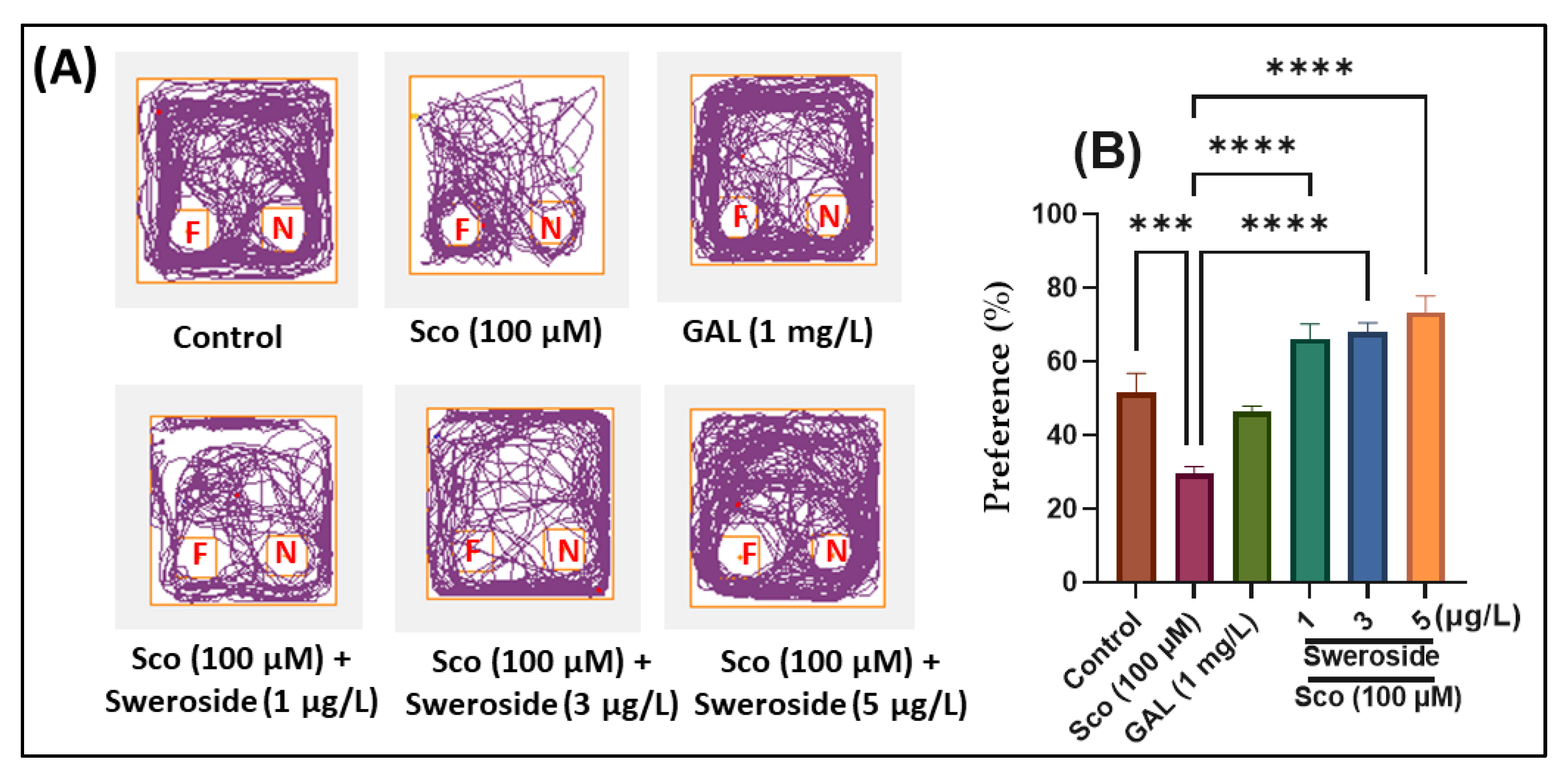
2.2. Effects on Response to Novelty and Recognition Memory in the Y-maze and NOR Tests
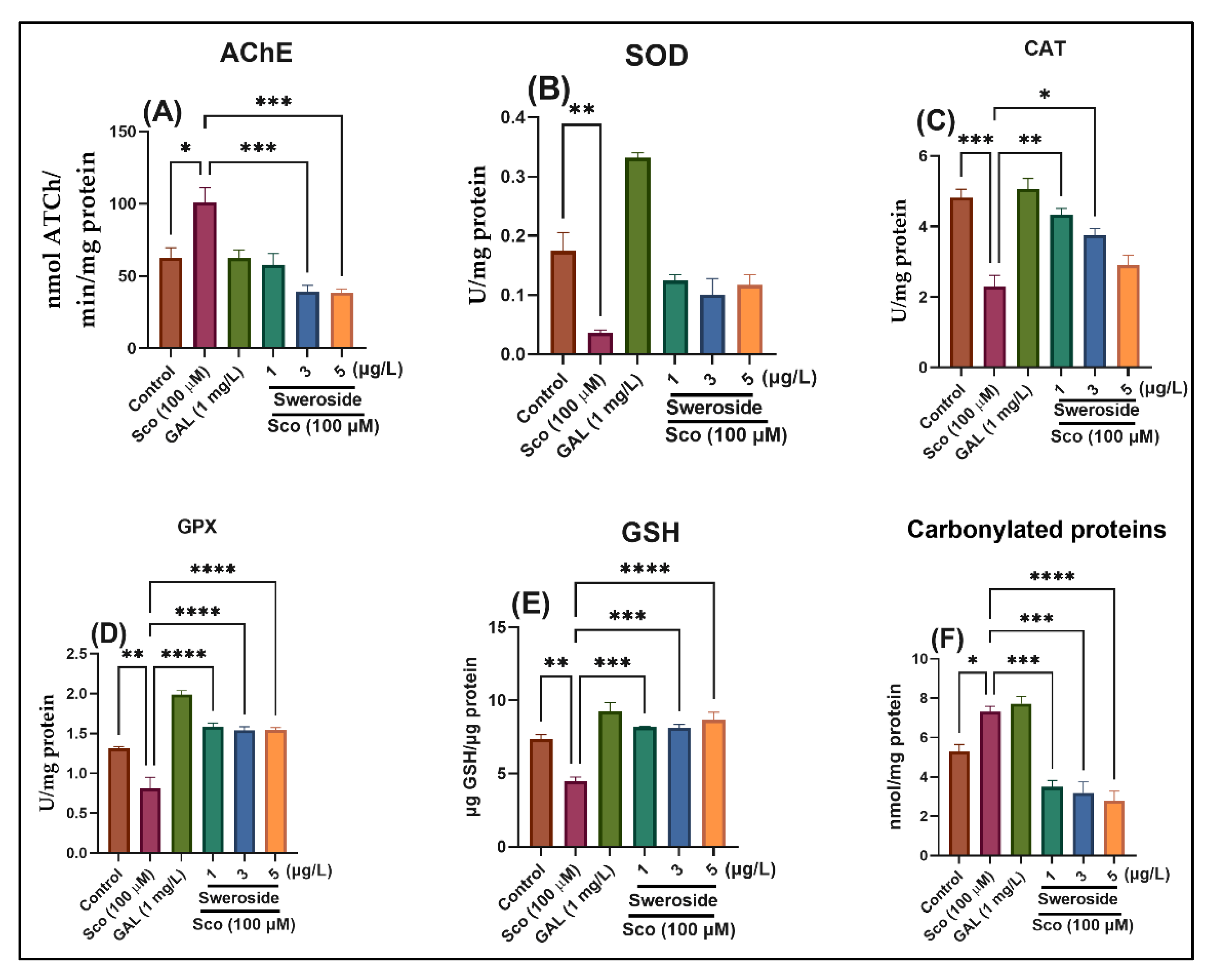
2.3. Effects on AChE Activity
2.4. Effects on SOD-, CAT-, and GPX-Specific Activities
2.5. Effects on GSH and Carbonylated Protein Levels
3. Materials and Methods
3.1. Plant Material and Extraction Method
3.2. Isolation of Sweroside
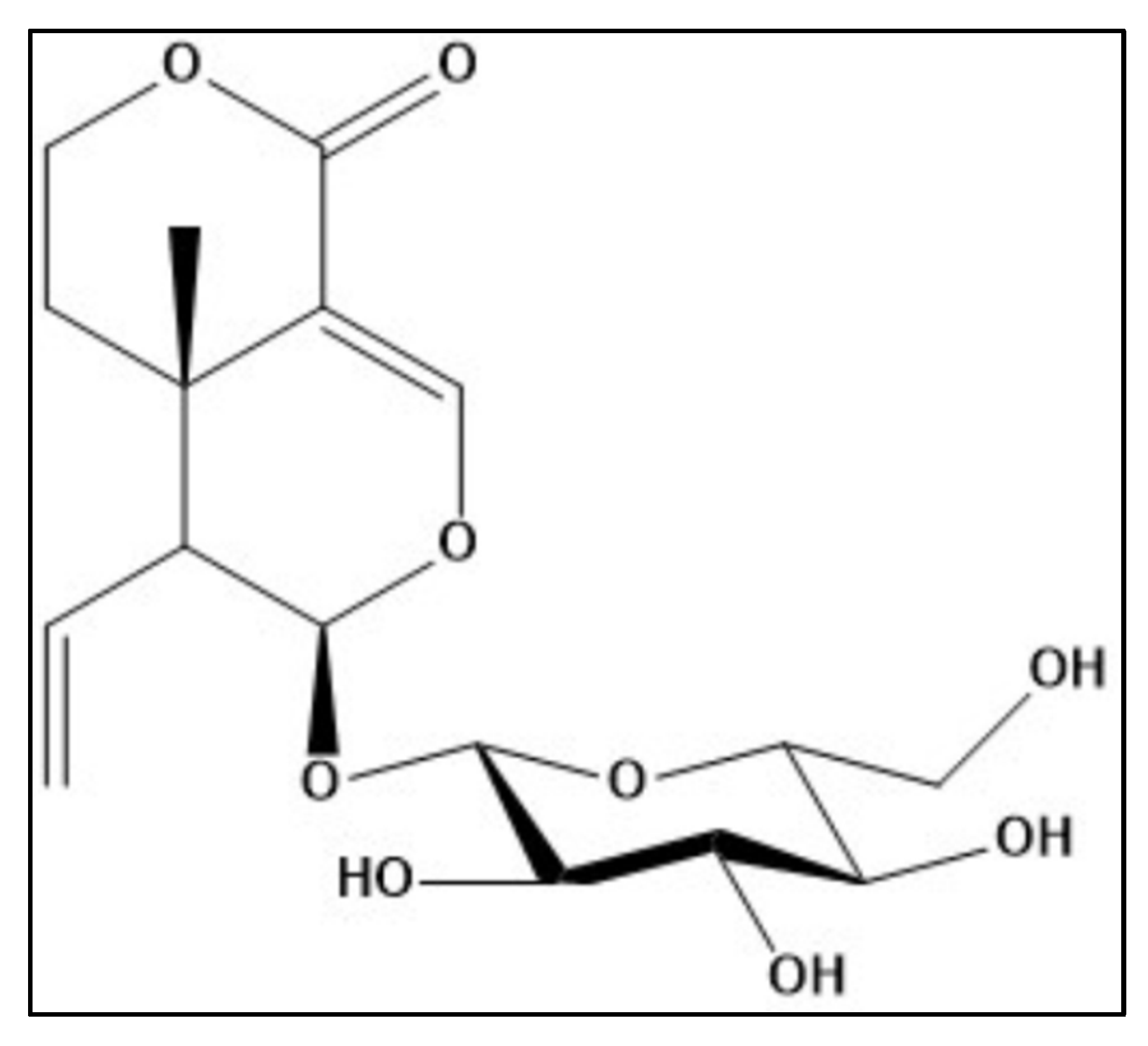
3.3. Structure Elucidation of Sweroside
3.4. Animals
3.5. Novel Tank Diving Test (NTT)
3.6. Y-maze Test
3.7. Novel Object Recognition Test (NOR)
3.8. Acetylcholinesterase (AChE), Antioxidant Enzyme, Glutathione (GSH), and Protein Carbonyl Measurment
3.9. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Oh, S.-Y.; Jang, M.J.; Choi, Y.-H.; Hwang, H.; Rhim, H.; Lee, B.; Choi, C.W.; Kim, M.S. Central administration of afzelin extracted from Ribes fasciculatum improves cognitive and memory function in a mouse model of dementia. Sci. Rep. 2021, 11, 9182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.J.; Luo, D.; Li, L.; Tan, R.R.; Xu, Q.Q.; Qin, J.; Zhu, L.; Luo, N.C.; Xu, T.T.; Zhang, R.; et al. Ethyl Acetate Extract Components of Bushen-Yizhi Formula Provides Neuroprotection against Scopolamine-induced Cognitive Impairment. Sci. Rep. 2017, 7, 9824. [Google Scholar] [CrossRef] [PubMed]
- Tönnies, E.; Trushina, E. Oxidative Stress, Synaptic Dysfunction, and Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 57, 1105–1121. [Google Scholar] [CrossRef] [PubMed]
- Kotani, S.; Yamauchi, T.; Teramoto, T.; Ogura, H. Donepezil, an acetylcholinesterase inhibitor, enhances adult hippocampal neurogenesis. Chem.-Biol. Interact. 2008, 175, 227–230. [Google Scholar] [CrossRef]
- Singab, A.N.B.; Mostafa, N.M.; Eldahshan, O.A.; Ashour, M.L.; Wink, M. Profile of volatile components of hydrodistilled and extracted leaves of Jacaranda acutifolia and their antimicrobial activity against foodborne pathogens. Nat. Prod. Commun. 2014, 9, 1007–1010. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, N.M.; Ashour, M.L.; Eldahshan, O.A.; Singab, A.N.B. Cytotoxic activity and molecular docking of a novel biflavonoid isolated from Jacaranda acutifolia (Bignoniaceae). Nat. Prod. Res. 2016, 30, 2093–2100. [Google Scholar] [CrossRef]
- Shahat, E.A.; Bakr, R.O.; Eldahshan, O.A.; Ayoub, N.A. Chemical Composition and Biological Activities of the Essential Oil from Leaves and Flowers of Pulicaria incisa sub. candolleana (Family Asteraceae). Chem. Biodivers. 2017, 14, e1600156. [Google Scholar] [CrossRef]
- Sheng, N.; Yuan, L.; Zhi, X.; Cui, C.; Zhang, Z.; Jia, P.; Zhang, X.; Zhang, L.; Wang, X. Application of a liquid chromatography-tandem mass spectrometry method to the pharmacokinetics, tissue distribution and excretion studies of sweroside in rats. J. Chromatogr. B. 2014, 969, 1–11. [Google Scholar] [CrossRef]
- Wang, R.; Dong, Z.; Lan, X.; Liao, Z.; Chen, M. Sweroside Alleviated LPS-Induced Inflammation via SIRT1 Mediating NF-κB and FOXO1 Signaling Pathways in RAW264.7 Cells. Molecules 2019, 24, 872. [Google Scholar] [CrossRef]
- Han, X.L.; Li, J.D.; Wang, W.L.; Yang, C.; Li, Z.Y. Sweroside eradicated leukemia cells and attenuated pathogenic processes in mice by inducing apoptosis. Biomed. Pharmacother. 2017, 95, 477–486. [Google Scholar] [CrossRef]
- Yang, Q.; Shu, F.; Gong, J.; Ding, P.; Cheng, R.; Li, J.; Tong, R.; Ding, L.; Sun, H.; Huang, W.; et al. Sweroside ameliorates NAFLD in high-fat diet induced obese mice through the regulation of lipid metabolism and inflammatory response. J. Ethnopharmacol. 2020, 255, 112556. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.L.; Yang, F.; Gong, J.T.; Tang, X.W.; Wang, G.Y.; Wang, Z.T.; Yang, L. Sweroside ameliorates α-naphthylisothiocyanate-induced cholestatic liver injury in mice by regulating bile acids and suppressing pro-inflammatory responses. Acta Pharmacol. Sin. 2016, 37, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.J.; Li, J.; Mei, Z.Y.; Chen, G. Gentiopicroside and sweroside from Veratrilla baillonii Franch. induce phosphorylation of Akt and suppress Pck1 expression in hepatoma cells. Biochem. Cell Biol. 2016, 94, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, L.; Zhang, A.; Zhang, N.; Lv, H.; Sun, W.; Wang, X. Protective effects of sweroside on human MG-63 cells and rat osteoblasts. Fitoterapia 2013, 84, 174–179. [Google Scholar] [CrossRef]
- Ye, F.; Tian, S.; Hu, H.; Yu, Z. Electroacupuncture reduces scopolamine-induced amnesia via mediating the miR-210/SIN3A and miR-183/SIN3A signaling pathway. Mol. Med. 2020, 26, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Budzynska, B.; Boguszewska-Czubara, A.; Kruk-Slomka, M.; Skalicka-Wozniak, K.; Michalak, A.; Musik, I.; Biala, G. Effects of imperatorin on scopolamine-induced cognitive impairment and oxidative stress in mice. Psychopharmacology 2015, 232, 931–942. [Google Scholar] [CrossRef]
- Mahendran, G.; Thamotharan, G.; Sengottuvelu, S.; Bai, V.N. Evaluation of anticonvulsant, sedative, anxiolytic, and phytochemical profile of the methanol extract from the aerial parts of Swertia corymbosa (Griseb.) wight ex C.B. Clarke. Biomed Res. Int. 2014, 2014, 542385. [Google Scholar] [CrossRef]
- Wang, Z.X.; Lian, W.W.; He, J.; He, X.L.; Wang, Y.M.; Pan, C.H.; Li, M.; Zhang, W.K.; Liu, L.Q.; Xu, J.K. Cornuside ameliorates cognitive impairments in scopolamine induced AD mice: Involvement of neurotransmitter and oxidative stress. J. Ethnopharmacol. 2022, 293, 115252. [Google Scholar] [CrossRef]
- Eskandarzadeh, M.; Kordestani-Moghadam, P.; Pourmand, S.; Khalili Fard, J.; Almassian, B.; Gharaghani, S. Inhibition of GSK_3β by Iridoid Glycosides of Snowberry ( Symphoricarpos albus) Effective in the Treatment of Alzheimer’s Disease Using Computational Drug Design Methods. Front. Chem. 2021, 9, 709932. [Google Scholar] [CrossRef]
- Agrawal, R.; Tyagi, E.; Saxena, G.; Nath, C. Cholinergic influence on memory stages: A study on scopolamine amnesic mice. Indian J. Pharmacol. 2009, 41, 192. [Google Scholar] [CrossRef]
- Karthivashan, G.; Park, S.-Y.Y.; Kweon, M.-H.H.; Kim, J.; Haque, M.E.; Cho, D.-Y.Y.; Kim, I.-S.S.; Cho, E.-A.A.; Ganesan, P.; Choi, D.-K.K. Ameliorative potential of desalted Salicornia europaea L. extract in multifaceted Alzheimer’s-like scopolamine-induced amnesic mice model. Sci. Rep. 2018, 8, 8826. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Lin, J.; Zhang, H.; Liu, Y.; Kan, M.; Xiu, Z.; Chen, X.; Lan, X.; Li, X.; Shi, X.; et al. Ginsenoside Compound K Regulates Amyloid β via the Nrf2/Keap1 Signaling Pathway in Mice with Scopolamine Hydrobromide-Induced Memory Impairments. J. Mol. Neurosci. 2019, 67, 62–71. [Google Scholar] [CrossRef]
- Xu, Q.Q.; Xu, Y.J.; Yang, C.; Tang, Y.; Li, L.; Cai, H.-B.; Hou, B.N.; Chen, H.F.; Wang, Q.; Shi, X.G.; et al. Sodium Tanshinone IIA Sulfonate Attenuates Scopolamine-Induced Cognitive Dysfunctions via Improving Cholinergic System. Biomed Res. Int. 2016, 2016, 9852536. [Google Scholar] [CrossRef]
- Song, S.H.; Choi, S.M.; Kim, J.E.; Sung, J.E.; Lee, H.A.; Choi, Y.H.; Bae, C.J.; Choi, Y.W.; Hwang, D.Y. α-Isocubebenol alleviates scopolamine-induced cognitive impairment by repressing acetylcholinesterase activity. Neurosci. Lett. 2017, 638, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Araujo, J.A.; Studzinski, C.M.; Milgram, N.W. Further evidence for the cholinergic hypothesis of aging and dementia from the canine model of aging. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Blokland, A. Scopolamine-induced deficits in cognitive performance: A review of animal studies. Scopolamine Rev. 2005, 1, 1–76. [Google Scholar]
- Paola, D.; Di Paola, D.; Capparucci, F.; Lanteri, G.; Crupi, R.; Marino, Y.; Antonio Franco, G.; Cuzzocrea, S.; Spanò, N.; Gugliandolo, E.; et al. Environmental Toxicity Assessment of Sodium Fluoride and Platinum-Derived Drugs Co-Exposure on Aquatic Organisms. Toxics 2022, 10, 272. [Google Scholar] [CrossRef]
- Cheng, Q.; Feng, X.; Meng, Q.; Li, Y.; Chen, S.; Wang, G.; Nie, K. [6]-Gingerol Ameliorates Cisplatin-Induced Pica by Regulating the TPH/MAO-A/SERT/5-HT/5-HT 3 Receptor System in Rats. Drug Des. Devel. Ther. 2020, 14, 4085–4099. [Google Scholar] [CrossRef]
- Fukui, K.; Onodera, K.; Shinkai, T.; Suzuki, S.; Urano, S. Impairment of learning and memory in rats caused by oxidative stress and aging, and changes in antioxidative defense systems. Ann. N. Y. Acad. Sci. 2001, 928, 168–175. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Giustarini, D.; Colombo, R.; Rossi, R.; Milzani, A. Protein carbonylation in human diseases. Trends Mol. Med. 2003, 9, 169–176. [Google Scholar] [CrossRef]
- Kong, D.; Yan, Y.; He, X.Y.; Yang, H.; Liang, B.Y.; Wang, J.; He, Y.; Ding, Y.; Yu, H. Effects of Resveratrol on the Mechanisms of Antioxidants and Estrogen in Alzheimer’s Disease. Biomed Res. Int. 2019, 2019, 8983752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Paola, D.; Abbate, J.M.; Iaria, C.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Environmental Risk Assessment of Dexamethasone Sodium Phosphate and Tocilizumab Mixture in Zebrafish Early Life Stage (Danio rerio). Toxics 2022, 10, 279. [Google Scholar] [CrossRef] [PubMed]
- Pauli, G.F.; Nauman, M.; Fischer, N.H. Application of Soft Pulse 1D NMR: Sweroside from a Potential Native American Anti-TB Drug. Spectrosc. Lett. 2006, 28, 903–913. [Google Scholar] [CrossRef]
- Damo, J.L.K.; Boiangiu, R.S.; Brinza, I.; Kenko Djoumessi, L.B.; Rebe, R.N.; Kamleu, B.N.; Guedang, S.D.N.; Camdi, G.W.; Bouvourné, P.; Keugong, E.W.; et al. Neuroprotective Potential of Guiera senegalensis (Combretaceae) Leaf Hydroethanolic Extract against Cholinergic System Dysfunctions and Oxidative Stress in Scopolamine-Induced Cognitive Impairment in Zebrafish ( Danio rerio). Plants 2022, 11, 1149. [Google Scholar] [CrossRef] [PubMed]
- Bate, S.T.; Clark, R.A. The Design and Statistical Analysis of Animal Experiments; Cambridge University Press: Cambridge, UK, 2014; ISBN 9781139344319. [Google Scholar]
- du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Cachat, J.; Stewart, A.; Grossman, L.; Gaikwad, S.; Kadri, F.; Chung, K.M.; Wu, N.; Wong, K.; Roy, S.; Suciu, C.; et al. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat. Protoc. 2010, 5, 1786–1799. [Google Scholar] [CrossRef]
- Boiangiu, R.S.; Bagci, E.; Dumitru, G.; Hritcu, L.; Todirascu-Ciornea, E. Angelica purpurascens (Avé-Lall.) Gilli. Essential Oil Improved Brain Function via Cholinergic Modulation and Antioxidant Effects in the Scopolamine-Induced Zebrafish (Danio rerio) Model. Plants 2022, 11, 1096. [Google Scholar] [CrossRef] [PubMed]
- Capatina, L.; Napoli, E.M.; Ruberto, G.; Hritcu, L. Origanum vulgare ssp. hirtum (Lamiaceae) Essential Oil Prevents Behavioral and Oxidative Stress Changes in the Scopolamine Zebrafish Model. Molecules 2021, 26, 7085. [Google Scholar] [CrossRef]
- Brinza, I.; Ayoub, I.M.; Eldahshan, O.A.; Hritcu, L. Baicalein 5,6-Dimethyl Ether Prevents Memory Deficits in the Scopolamine Zebrafish Model by Regulating Cholinergic and Antioxidant Systems. Plants 2021, 10, 1245. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brinza, I.; Raey, M.A.E.; El-Kashak, W.; Eldahshan, O.A.; Hritcu, L. Sweroside Ameliorated Memory Deficits in Scopolamine-Induced Zebrafish (Danio rerio) Model: Involvement of Cholinergic System and Brain Oxidative Stress. Molecules 2022, 27, 5901. https://doi.org/10.3390/molecules27185901
Brinza I, Raey MAE, El-Kashak W, Eldahshan OA, Hritcu L. Sweroside Ameliorated Memory Deficits in Scopolamine-Induced Zebrafish (Danio rerio) Model: Involvement of Cholinergic System and Brain Oxidative Stress. Molecules. 2022; 27(18):5901. https://doi.org/10.3390/molecules27185901
Chicago/Turabian StyleBrinza, Ion, Mohamed A. El Raey, Walaa El-Kashak, Omayma A. Eldahshan, and Lucian Hritcu. 2022. "Sweroside Ameliorated Memory Deficits in Scopolamine-Induced Zebrafish (Danio rerio) Model: Involvement of Cholinergic System and Brain Oxidative Stress" Molecules 27, no. 18: 5901. https://doi.org/10.3390/molecules27185901
APA StyleBrinza, I., Raey, M. A. E., El-Kashak, W., Eldahshan, O. A., & Hritcu, L. (2022). Sweroside Ameliorated Memory Deficits in Scopolamine-Induced Zebrafish (Danio rerio) Model: Involvement of Cholinergic System and Brain Oxidative Stress. Molecules, 27(18), 5901. https://doi.org/10.3390/molecules27185901