
β2-Adrenergic Receptor Signaling Pathway Stimulates the Migration and Invasion of Cancer Cells via Src Activation

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Expression of ADRB2 in Human HCC and BC Cell Lines
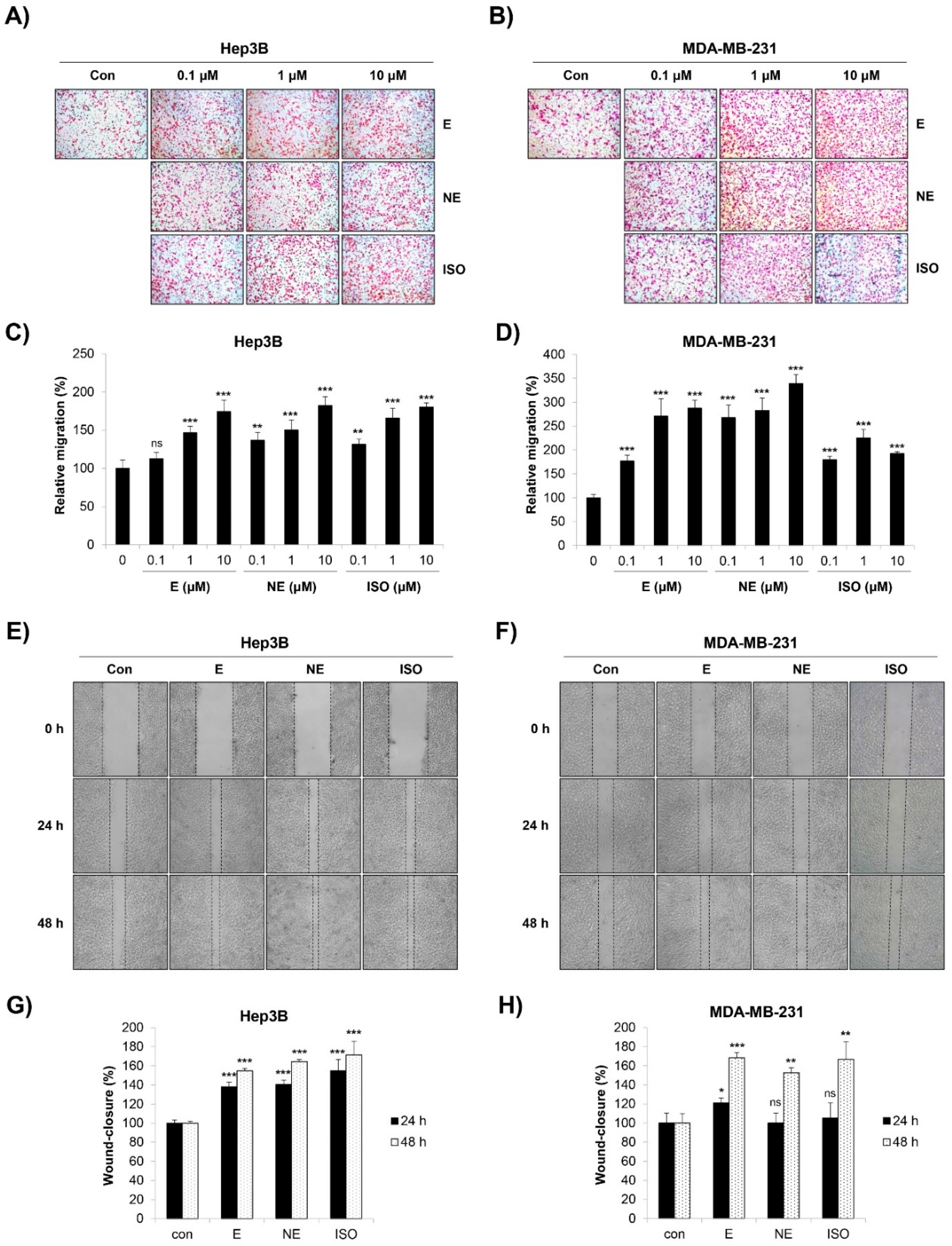
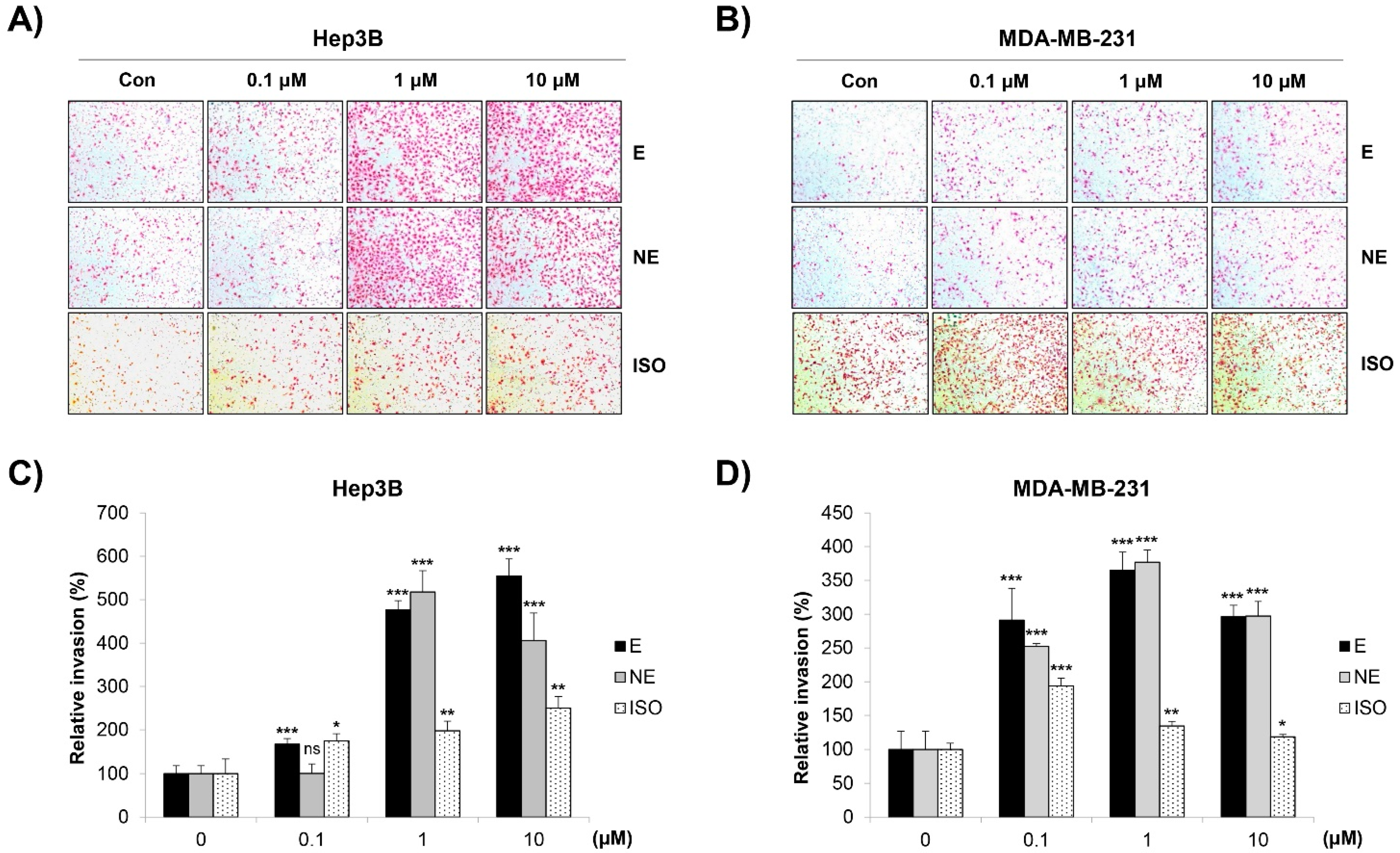
2.2. Effect of β-AR Signaling Pathway on the Migration and Invasion of Human HCC and BC Cells
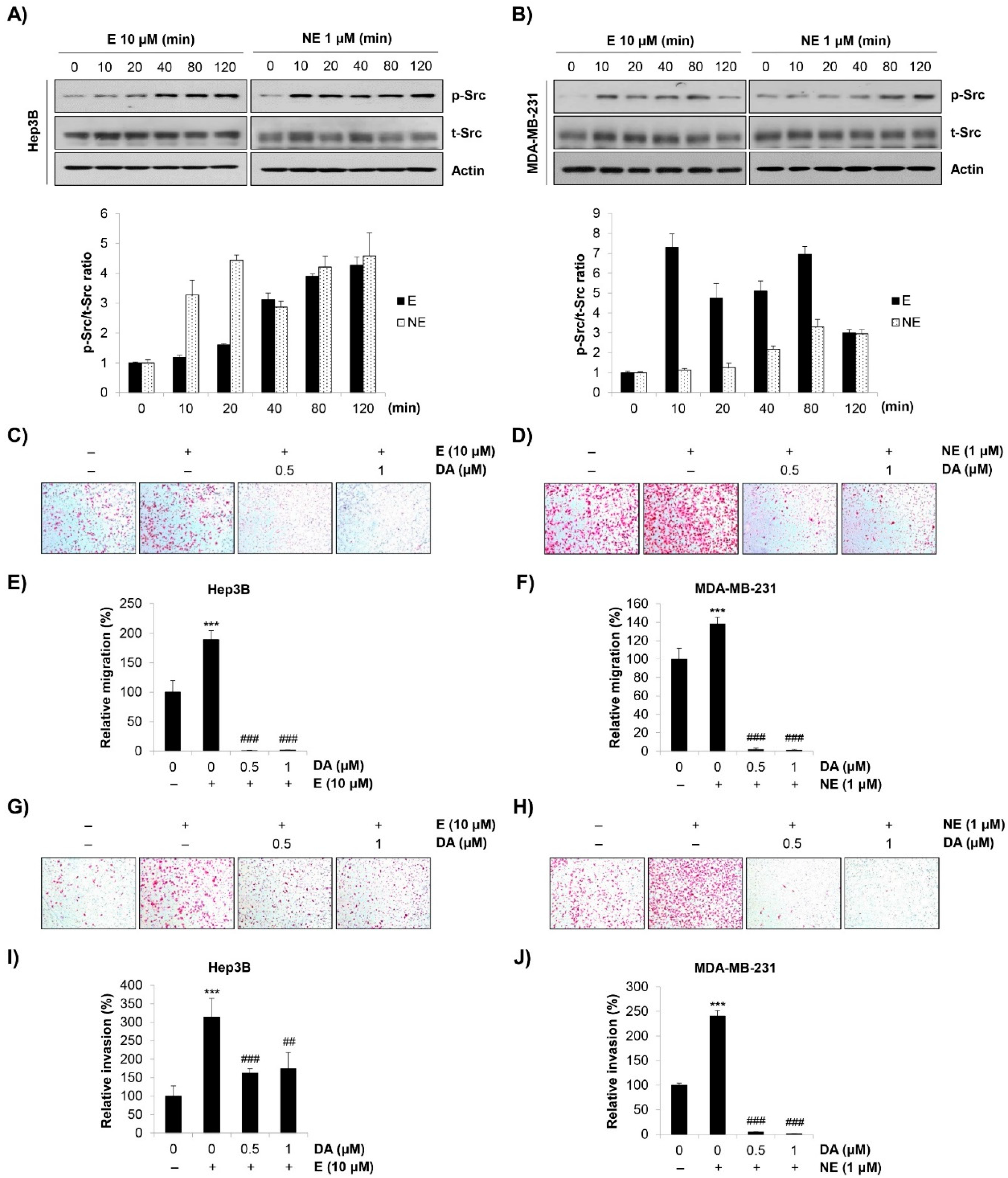
2.3. Role of Src in E/NE-Induced Migration and Invasion in Human HCC and BC Cells
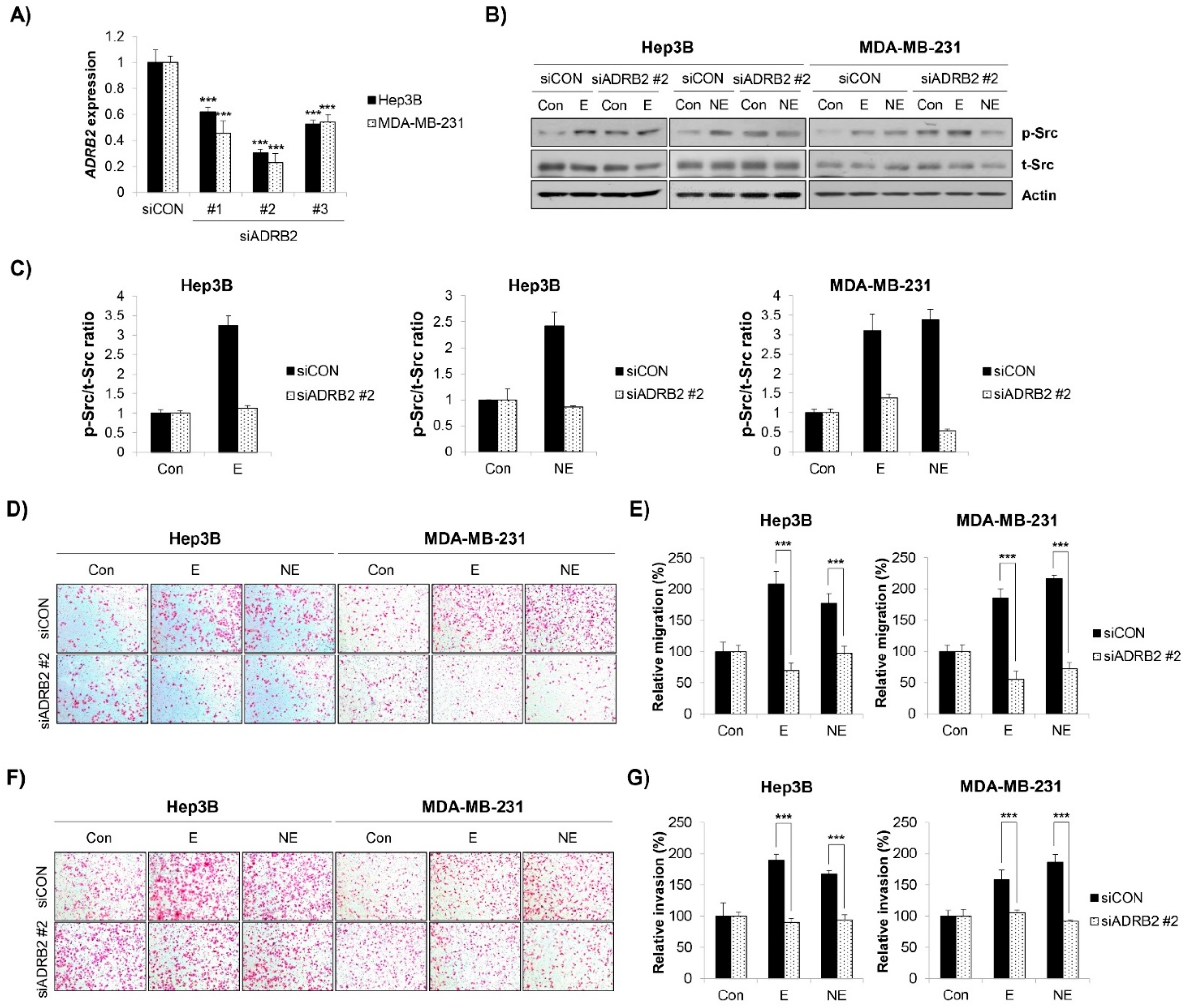
2.4. β2-AR/Src Axis Mediated E/NE-Induced Migration and Invasion in HCC and BC Cells
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Lines and Cell Culture
4.3. Transwell Assay
4.4. Wound-Healing Assay
4.5. Knockdown of ADRB2
4.6. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
4.7. Western Blot Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cui, B.; Peng, F.; Lu, J.; He, B.; Su, Q.; Luo, H.; Deng, Z.; Jiang, T.; Su, K.; Huang, Y.; et al. Cancer and stress: NextGen strategies. Brain Behav. Immun. 2021, 93, 368–383. [Google Scholar] [CrossRef] [PubMed]
- Bernabé, D.G. Catecholamines Mediate Psychologic Stress-Induced Cancer Progression. Cancer Res. 2021, 81, 5144–5146. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.W.; Sood, A.K. Molecular pathways: Beta-adrenergic signaling in cancer. Clin. Cancer Res. 2012, 18, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Partecke, L.I.; Speerforck, S.; Käding, A.; Seubert, F.; Kühn, S.; Lorenz, E.; Schwandke, S.; Sendler, M.; Keßler, W.; Trung, D.N.; et al. Chronic stress increases experimental pancreatic cancer growth, reduces survival and can be antagonised by beta-adrenergic receptor blockade. Pancreatology 2016, 16, 423–433. [Google Scholar] [CrossRef]
- Nkontchou, G.; Aout, M.; Mahmoudi, A.; Roulot, D.; Bourcier, V.; Grando-Lemaire, V.; Ganne-Carrie, N.; Trinchet, J.C.; Vicaut, E.; Beaugrand, M. Effect of long-term propranolol treatment on hepatocellular carcinoma incidence in patients with HCV-associated cirrhosis. Cancer Prev. Res. 2012, 5, 1007–1014. [Google Scholar] [CrossRef]
- Chang, H.; Lee, S.H. Beta-adrenergic receptor blockers and hepatocellular carcinoma survival: A systemic review and meta-analysis. Clin. Exp. Med. 2022, 23. [Google Scholar] [CrossRef]
- Montoya, A.; Amaya, C.N.; Belmont, A.; Diab, N.; Trevino, R.; Villanueva, G.; Rains, S.; Sanchez, L.A.; Badri, N.; Otoukesh, S.; et al. Use of non-selective β-blockers is associated with decreased tumor proliferative indices in early stage breast cancer. Oncotarget 2007, 8, 6446–6460. [Google Scholar] [CrossRef]
- Barron, T.I.; Connolly, R.M.; Sharp, L.; Bennett, K.; Visvanathan, K. Beta-blocker and breast cancer mortality: A population-based study. J. Clin. Oncol. 2011, 29, 2635–2644. [Google Scholar] [CrossRef]
- Kaira, K.; Kamiyoshihara, M.; Kawashima, O.; Endoh, H.; Imaizumi, K.; Sugano, M.; Tanaka, S.; Fujita, A.; Kogure, Y.; Shimizu, A.; et al. Prognostic impact of beta2 adrenergic receptor expression in surgically resected pulmonary pleomorphic carcinoma. Anticancer Res. 2019, 39, 395–403. [Google Scholar] [CrossRef]
- Nilsson, M.B.; Sun, H.; Diao, L.; Tong, P.; Liu, D.; Li, L.; Fan, Y.; Poteete, A.; Lim, S.O.; Howells, K.; et al. Stress hormones promote EGFR inhibitor resistance in NSCLC: Implications for combinations with beta-blockers. Sci. Transl. Med. 2017, 9, eaao4307. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jiang, S. The role of ADRB2 gene polymorphisms in malignancies. Mol. Biol. Rep. 2021, 48, 2741–2749. [Google Scholar] [CrossRef] [PubMed]
- Thaker, P.H.; Han, L.Y.; Kamat, A.A.; Arevalo, J.M.; Takahashi, R.; Lu, C.; Jennings, N.B.; Armaiz-Pena, G.; Bankson, J.A.; Ravoori, M.; et al. Chronic stress promotes tumor growth and angiogenesis in a mouse model of ovarian carcinoma. Nat. Med. 2006, 12, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Ma, J.; Ma, Q.; Guo, K.; Guo, J.; Li, X.; Li, W.; Liu, J.; Huang, C.; Wang, F.; et al. beta2-AR-HIF-1alpha: A novel regulatory axis for 12 stress-induced pancreatic tumor growth and angiogenesis. Curr. Mol. Med. 2013, 13, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; He, Z.; Yin, K.; Li, B.; Zhang, L.; Xu, Z. Chronic stress promotes gastric cancer progression and metastasis: An essential role for ADRB2. Cell Death. Dis. 2019, 10, 788. [Google Scholar] [CrossRef] [PubMed]
- Aleshin, A.; Finn, R.S. SRC: A century of science brought to the clinic. Neoplasia 2010, 12, 599–607. [Google Scholar] [CrossRef]
- Armaiz-Pena, G.N.; Allen, J.K.; Cruz, A.; Stone, R.L.; Nick, A.M.; Lin, Y.G.; Han, L.Y.; Mangala, L.S.; Villares, G.J.; Vivas-Mejia, P.; et al. Src activation by β-adrenoreceptors is a key switch for tumour metastasis. Nat. Commun. 2013, 4, 1403. [Google Scholar] [CrossRef]
- Spini, A.; Roberto, G.; Gini, R.; Bartolini, C.; Bazzani, L.; Donnini, S.; Crispino, S.; Ziche, M. Evidence of β-blockers drug repurposing for the treatment of triple negative breast cancer: A systematic review. Neoplasma 2019, 66, 963–970. [Google Scholar] [CrossRef]
- Wang, F.; Liu, H.; Wang, F.; Xu, R.; Wang, P.; Tang, F.; Zhang, X.; Zhu, Z.; Lv, H.; Han, T.; et al. Propranolol suppresses the proliferation and induces the apoptosis of liver cancer cells. Mol. Med. Rep. 2018, 17, 5213–5221. [Google Scholar] [CrossRef]
- Boulay, G.; Malaquin, N.; Loison, I.; Foveau, B.; Van Rechem, C.; Rood, B.R.; Pourtier, A.; Leprince, D. Loss of Hypermethylated in Cancer 1 (HIC1) in breast cancer cells contributes to stress-induced migration and invasion through β-2 adrenergic receptor (ADRB2) misregulation. J. Biol. Chem. 2012, 287, 5379–5389. [Google Scholar] [CrossRef]
- Boulay, G.; Malaquin, N.; Loison, I.; Foveau, B.; Van Rechem, C.; Rood, B.R.; Pourtier, A.; Leprince, D. β2-Adrenoceptors on tumor cells play a critical role in stress-enhanced metastasis in a mouse model of breast cancer. Brain Behav. Immun. 2016, 57, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yang, X.M.; Wang, Y.H.; Feng, M.X.; Liu, X.J.; Zhang, Y.L.; Huang, S.; Wu, Z.; Xue, F.; Qin, W.X.; et al. Monoamine oxidase A suppresses hepatocellular carcinoma metastasis by inhibiting the adrenergic system and its transactivation of EGFR signaling. J. Hepatol. 2014, 60, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Li, S.; Ma, R.; Guan, S.; Li, J.; Li, H.; Zhang, H.; Lin, Q.; Zhao, Z.; Wang, B. Effects of liver depression and psychological stress on human uterine leiomyoma cells by an AR-cAMP-PKA signal transduction pathway. Taiwan J. Obstet. Gynecol. 2017, 56, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Min, T.R.; Park, H.J.; Ha, K.T.; Chi, G.Y.; Choi, Y.H.; Park, S.H. Suppression of EGFR/STAT3 activity by lupeol contributes to the induction of the apoptosis of human non-small cell lung cancer cells. Int. J. Oncol. 2019, 55, 320–330. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, J.-H.; Park, H.-J.; Park, S.-H.; Choi, Y.-H.; Chi, G.-Y. β2-Adrenergic Receptor Signaling Pathway Stimulates the Migration and Invasion of Cancer Cells via Src Activation. Molecules 2022, 27, 5940. https://doi.org/10.3390/molecules27185940
Jeong J-H, Park H-J, Park S-H, Choi Y-H, Chi G-Y. β2-Adrenergic Receptor Signaling Pathway Stimulates the Migration and Invasion of Cancer Cells via Src Activation. Molecules. 2022; 27(18):5940. https://doi.org/10.3390/molecules27185940
Chicago/Turabian StyleJeong, Jae-Hoon, Hyun-Ji Park, Shin-Hyung Park, Yung-Hyun Choi, and Gyoo-Yong Chi. 2022. "β2-Adrenergic Receptor Signaling Pathway Stimulates the Migration and Invasion of Cancer Cells via Src Activation" Molecules 27, no. 18: 5940. https://doi.org/10.3390/molecules27185940
APA StyleJeong, J.-H., Park, H.-J., Park, S.-H., Choi, Y.-H., & Chi, G.-Y. (2022). β2-Adrenergic Receptor Signaling Pathway Stimulates the Migration and Invasion of Cancer Cells via Src Activation. Molecules, 27(18), 5940. https://doi.org/10.3390/molecules27185940