
Erdafitinib Inhibits Tumorigenesis of Human Lung Adenocarcinoma A549 by Inducing S-Phase Cell-Cycle Arrest as a CDK2 Inhibitor

 ,
, 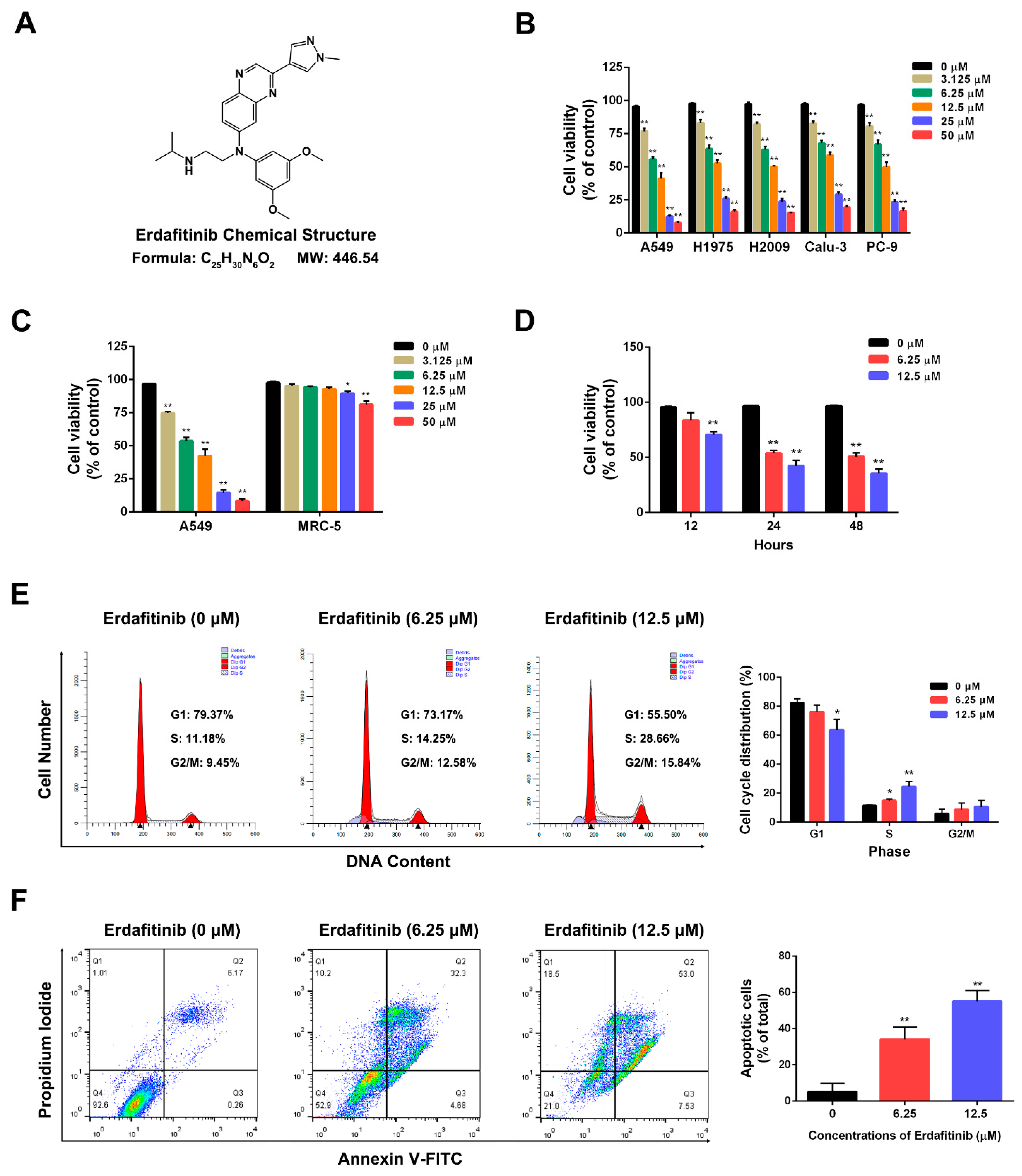{kind=link}
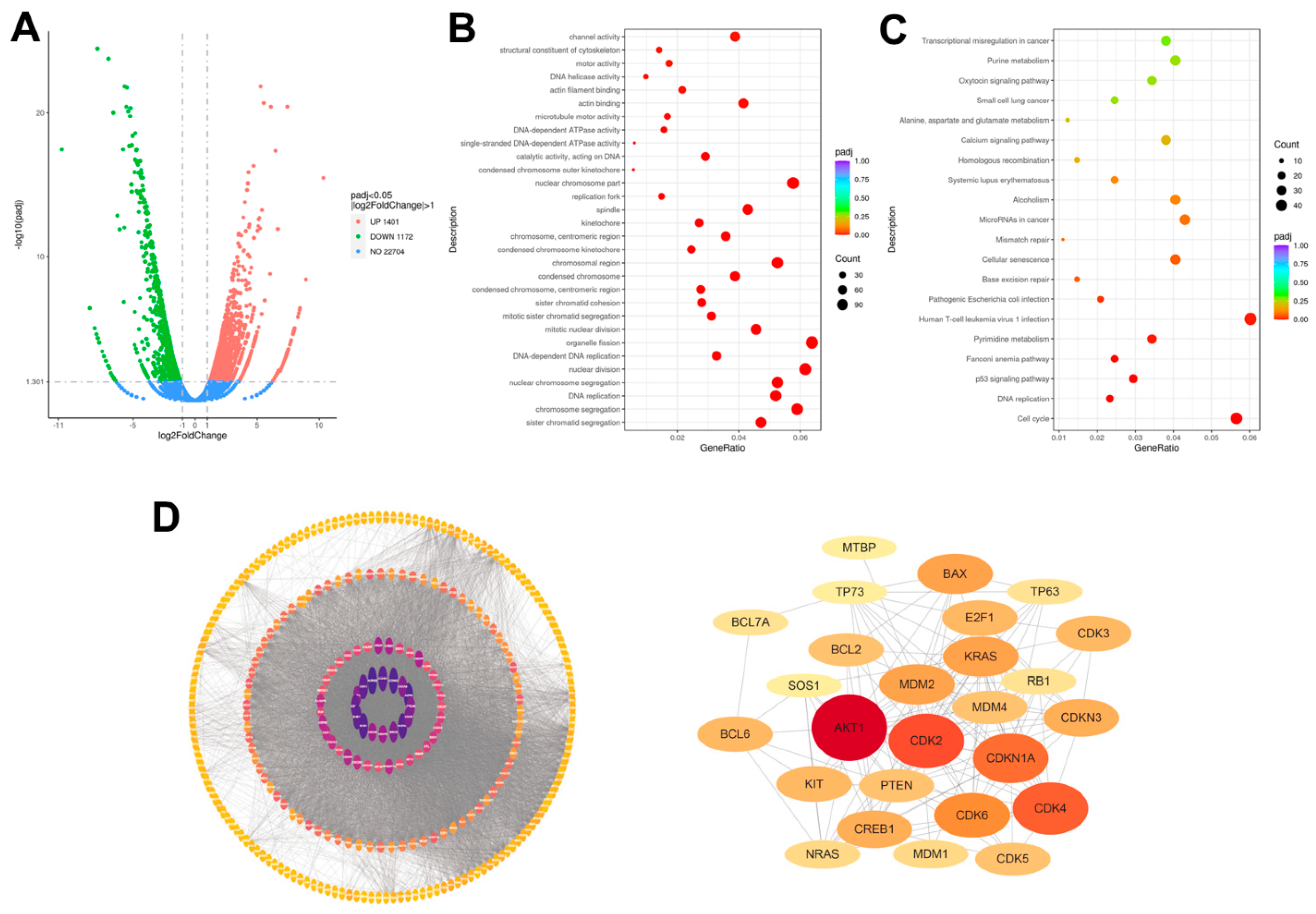{kind=link}
{kind=link}
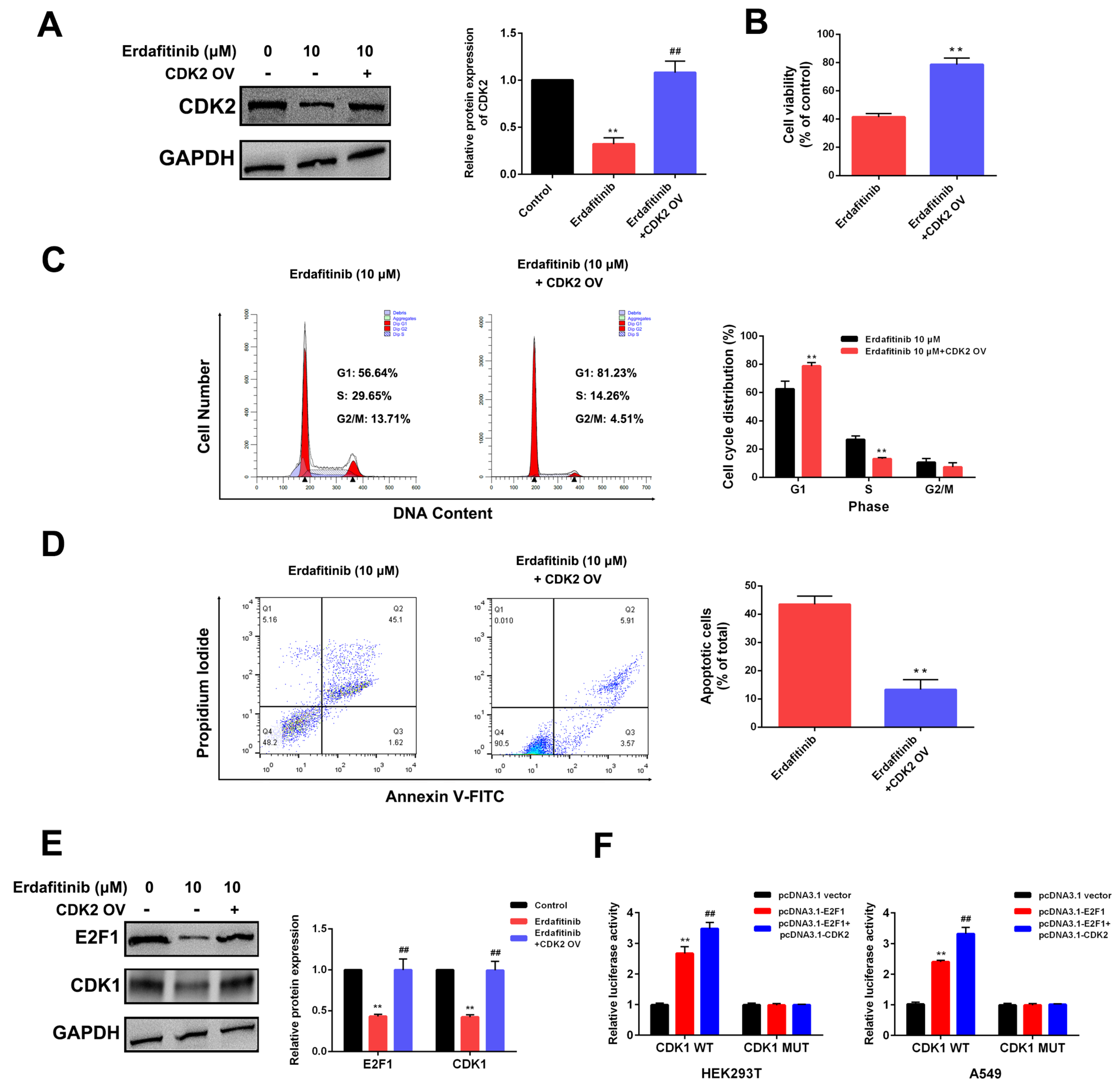{kind=link}
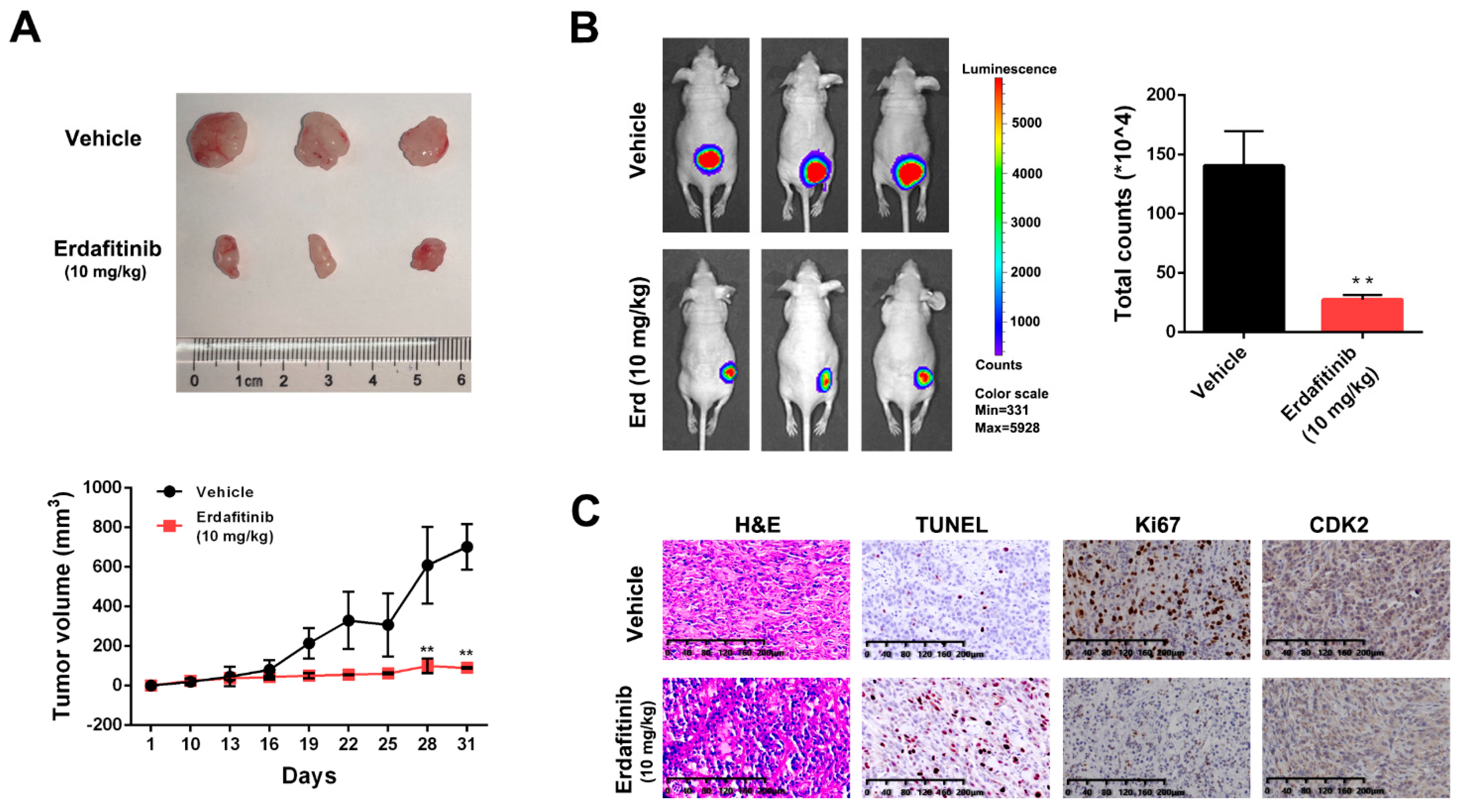{kind=link}
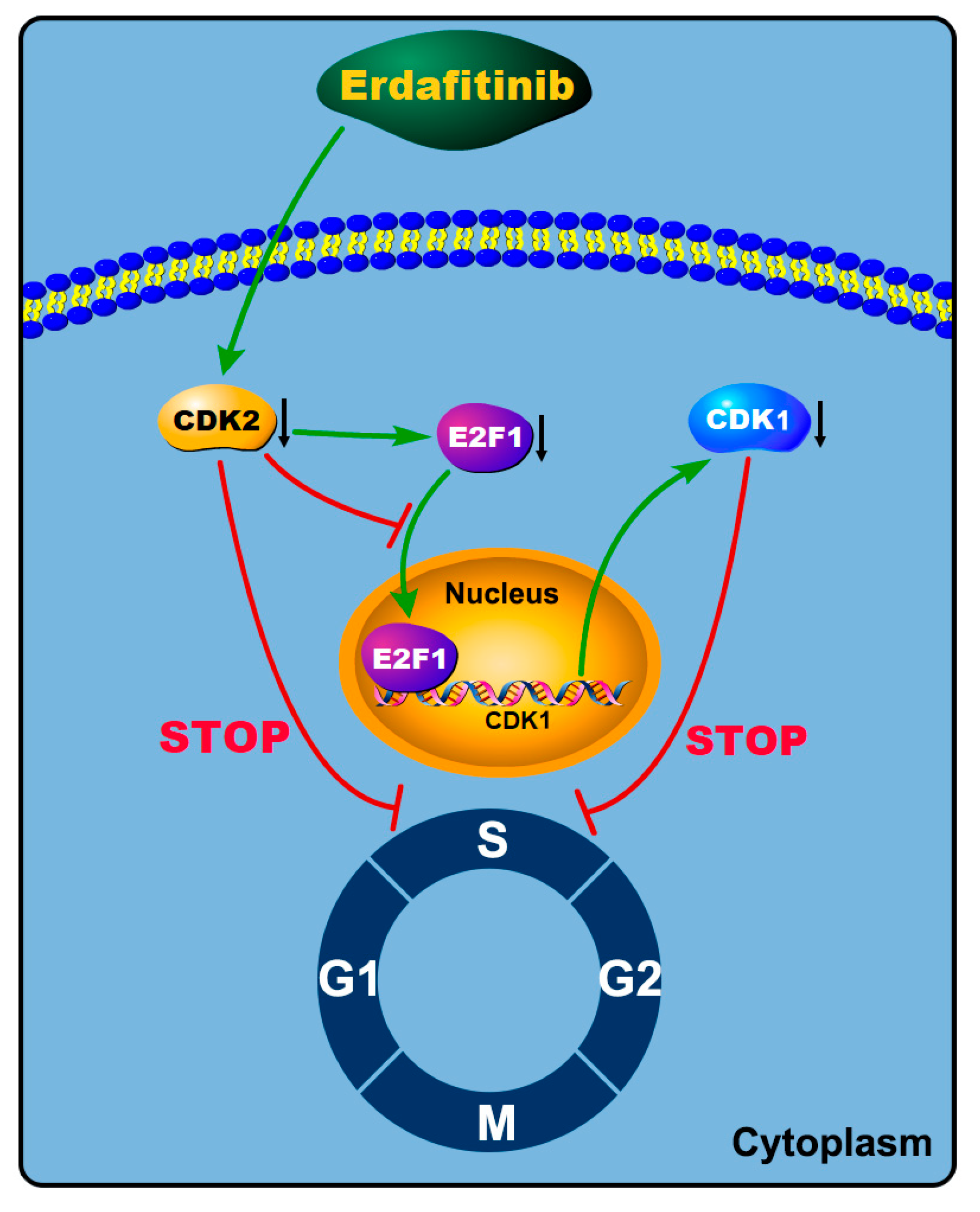{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Erdafitinib Induces S-Phase Cell-Cycle Arrest and Cell Apoptosis in Human Lung Adenocarcinoma A549 Cells
2.2. Transcriptomic Analysis of Human Lung Adenocarcinoma A549 Cells upon Erdafitinib Treatment
2.3. Cell-Cycle Network Analysis of Human Lung Adenocarcinoma A549 Cells upon Erdafitinib Treatment
2.4. CDK2 Is the Target of Erdafitinib in Human Lung Adenocarcinoma A549 Cells
2.5. Erdafitinib Suppresses Tumor Growth in a A549 Xenograft Mouse Model
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Lines and Culture
4.3. Cell Viability Analysis
4.4. Cell-Cycle Analysis
4.5. Cell Apoptosis Analysis
4.6. RNA Sequencing and Analysis
4.7. Quantitative Real-Time PCR (qRT-PCR)
4.8. Western Blot Analysis
4.9. Cell Transfection
4.10. Dual-Luciferase Reporter Assay
4.11. Nude Mice Tumorigenesis Assay
4.12. Histology and Immunohistochemistry
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Thai, A.A.; Solomon, B.J.; Sequist, L.V.; Gainor, J.F.; Heist, R.S. Lung cancer. Lancet Lond. Engl. 2021, 398, 535–554. [Google Scholar] [CrossRef]
- Nooreldeen, R.; Bach, H. Current and Future Development in Lung Cancer Diagnosis. Int. J. Mol. Sci. 2021, 22, 8661. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Denisenko, T.V.; Budkevich, I.N.; Zhivotovsky, B. Cell death-based treatment of lung adenocarcinoma. Cell Death Dis. 2018, 9, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchinson, B.D.; Shroff, G.S.; Truong, M.T.; Ko, J.P. Spectrum of Lung Adenocarcinoma. Semin. Ultrasound CT MR 2019, 40, 255–264. [Google Scholar] [CrossRef]
- Succony, L.; Rassl, D.M.; Barker, A.P.; McCaughan, F.M.; Rintoul, R.C. Adenocarcinoma spectrum lesions of the lung: Detection, pathology and treatment strategies. Cancer Treat. Rev. 2021, 99, 102237. [Google Scholar] [CrossRef]
- Du, X.; Yang, B.; An, Q.; Assaraf, Y.G.; Cao, X.; Xia, J. Acquired resistance to third-generation EGFR-TKIs and emerging next-generation EGFR inhibitors. Innovation 2021, 2, 100103. [Google Scholar] [CrossRef]
- Saito, M.; Shiraishi, K.; Kunitoh, H.; Takenoshita, S.; Yokota, J.; Kohno, T. Gene aberrations for precision medicine against lung adenocarcinoma. Cancer Sci. 2016, 107, 713–720. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Ma, R.; Cao, G.; Liu, H.; He, L.; Tang, L.; Li, H.; Luo, Q. Combined Treatment of Cinobufotalin and Gefitinib Exhibits Potent Efficacy against Lung Cancer. Evid. Based Complement. Alternat. Med. 2021, 2021, 6612365. [Google Scholar] [CrossRef]
- Roubal, K.; Myint, Z.W.; Kolesar, J.M. Erdafitinib: A novel therapy for FGFR-mutated urothelial cancer. Am. J. Health Syst. Pharm. 2020, 77, 346–351. [Google Scholar] [CrossRef]
- Weaver, A.; Bossaer, J.B. Fibroblast growth factor receptor (FGFR) inhibitors: A review of a novel therapeutic class. J. Oncol. Pharm. Pract. 2021, 27, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Montazeri, K.; Bellmunt, J. Erdafitinib for the treatment of metastatic bladder cancer. Expert Rev. Clin. Pharmacol. 2020, 13, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bansal, P.; Dwivedi, D.K.; Hatwal, D.; Sharma, P.; Gupta, V.; Goyal, S.; Maithani, M. Erdafitinib as a Novel and Advanced Treatment Strategy of Metastatic Urothelial Carcinoma. Anticancer Agents Med. Chem. 2021, 21, 2478–2486. [Google Scholar] [CrossRef] [PubMed]
- Chandrani, P.; Prabhash, K.; Prasad, R.; Sethunath, V.; Ranjan, M.; Iyer, P.; Aich, J.; Dhamne, H.; Iyer, D.N.; Upadhyay, P.; et al. Drug-sensitive FGFR3 mutations in lung adenocarcinoma. Ann. Oncol. 2017, 28, 597–603. [Google Scholar] [CrossRef]
- Urrutia Argueta, S.A.; Hanna, N.H. Remarkable Response to Erdafitinib in Metastatic Lung Adenocarcinoma with FGFR Fusion. JCO Precis. Oncol. 2020, 4, 823–824. [Google Scholar] [CrossRef]
- Loriot, Y.; Necchi, A.; Park, S.H.; Garcia-Donas, J.; Huddart, R.; Burgess, E.; Fleming, M.; Rezazadeh, A.; Mellado, B.; Varlamov, S.; et al. Erdafitinib in Locally Advanced or Metastatic Urothelial Carcinoma. N. Engl. J. Med. 2019, 381, 338–348. [Google Scholar] [CrossRef]
- Roskoski, R., Jr. The role of fibroblast growth factor receptor (FGFR) protein-tyrosine kinase inhibitors in the treatment of cancers including those of the urinary bladder. Pharmacol. Res. 2020, 151, 104567. [Google Scholar] [CrossRef]
- Leal-Esteban, L.C.; Fajas, L. Cell cycle regulators in cancer cell metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165715. [Google Scholar] [CrossRef]
- Suski, J.M.; Braun, M.; Strmiska, V.; Sicinski, P. Targeting cell-cycle machinery in cancer. Cancer Cell 2021, 39, 759–778. [Google Scholar] [CrossRef]
- Ding, L.; Cao, J.; Lin, W.; Chen, H.; Xiong, X.; Ao, H.; Yu, M.; Lin, J.; Cui, Q. The Roles of Cyclin-Dependent Kinases in Cell-Cycle Progression and Therapeutic Strategies in Human Breast Cancer. Int. J. Mol. Sci. 2020, 21, 1960. [Google Scholar] [CrossRef]
- You, J.; Dong, R.; Ying, M.; He, Q.; Cao, J.; Yang, B. Cellular Senescence and Anti-Cancer Therapy. Curr. Drug Targets 2019, 20, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, M.; Shen, C. Bauerane Induces S-Phase Cell Cycle Arrest, Apoptosis, and Inhibition of Proliferation of A549 Human Lung Cancer Cells Through the Phosphoinositide 3-Kinase (PI3K)/AKT and Signal Transducer and Activator of Transcription 3 (STAT3) Signaling Pathway. Med. Sci. Monit. 2020, 26, e919558. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.B.; Chen, B.H. Inhibition of lung cancer cells A549 and H460 by curcuminoid extracts and nanoemulsions prepared from Curcuma longa Linnaeus. Int. J. Nanomed. 2015, 10, 5059–5080. [Google Scholar]
- Li, X.; Li, F.; Wang, F.; Li, J.; Lin, C.; Du, J. Resveratrol inhibits the proliferation of A549 cells by inhibiting the expression of COX-2. Onco Targets Ther. 2018, 11, 2981–2989. [Google Scholar] [CrossRef] [Green Version]
- Malumbres, M. Cyclin-dependent kinases. Genome. Biol. 2014, 15, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łukasik, P.; Baranowska-Bosiacka, I.; Kulczycka, K.; Gutowska, I. Inhibitors of Cyclin-Dependent Kinases: Types and Their Mechanism of Action. Int. J. Mol. Sci. 2021, 22, 2806. [Google Scholar] [CrossRef]
- Kciuk, M.; Gielecińska, A.; Mujwar, S.; Mojzych, M.; Kontek, R. Cyclin-dependent kinases in DNA damage response. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188716. [Google Scholar] [CrossRef]
- Goel, S.; DeCristo, M.J.; McAllister, S.S.; Zhao, J.J. CDK4/6 Inhibition in Cancer: Beyond Cell Cycle Arrest. Trends Cell Biol. 2018, 28, 911–925. [Google Scholar] [CrossRef]
- Braal, C.L.; Jongbloed, E.M.; Wilting, S.M.; Mathijssen, R.H.J.; Koolen, S.L.W.; Jager, A. Inhibiting CDK4/6 in Breast Cancer with Palbociclib, Ribociclib, and Abemaciclib: Similarities and Differences. Drugs 2021, 81, 317–331. [Google Scholar] [CrossRef]
- O’Leary, B.; Finn, R.S.; Turner, N.C. Treating cancer with selective CDK4/6 inhibitors. Nat. Rev. Clin. Oncol. 2016, 13, 417–430. [Google Scholar] [CrossRef]
- Ettl, T.; Schulz, D.; Bauer, R.J. The Renaissance of Cyclin Dependent Kinase Inhibitors. Cancers 2022, 14, 293. [Google Scholar] [CrossRef] [PubMed]
- Panagiotou, E.; Gomatou, G.; Trontzas, I.P.; Syrigos, N.; Kotteas, E. Cyclin-dependent kinase (CDK) inhibitors in solid tumors: A review of clinical trials. Clin. Transl. Oncol. 2022, 24, 161–192. [Google Scholar] [CrossRef] [PubMed]
- Mitra, J.; Enders, G.H. Cyclin A/Cdk2 complexes regulate activation of Cdk1 and Cdc25 phosphatases in human cells. Oncogene 2004, 23, 3361–3367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Meerloo, J.; Kaspers, G.J.; Cloos, J. Cell sensitivity assays: The MTT assay. Methods Mol. Biol. 2011, 731, 237–245. [Google Scholar]
- Weng, H.; Tan, Z.J.; Hu, Y.P.; Shu, Y.J.; Bao, R.F.; Jiang, L.; Wu, X.S.; Li, M.L.; Ding, Q.; Wang, X.A.; et al. Ursolic acid induces cell cycle arrest and apoptosis of gallbladder carcinoma cells. Cancer Cell Int. 2014, 14, 96. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-C.; Kim, H.G.; Lee, S.A.; Lim, S.; Park, E.H.; Kim, S.J.; Lim, C.J. Genipin-induced apoptosis in hepatoma cells is mediated by reactive oxygen species/c-Jun NH2-terminal kinase-dependent activation of mitochondrial pathway. Biochem. Pharmacol. 2005, 70, 1398–1407. [Google Scholar] [CrossRef]
- Qiu, J.; Zhang, W.; Zang, C.; Liu, X.; Liu, F.; Ge, R.; Sun, Y.; Xia, Q. Identification of key genes and miRNAs markers of papillary thyroid cancer. Biol. Res. 2018, 51, 45. [Google Scholar] [CrossRef]
- Fiedler, S.D.; Carletti, M.Z.; Christenson, L.K. Quantitative RT-PCR methods for mature microRNA expression analysis. Methods Mol. Biol. 2010, 630, 49–64. [Google Scholar]
- Fido, R.J.; Tatham, A.S.; Shewry, P.R. Western blotting analysis. In Plant Gene Transfer and Expression Protocols; Springer: Berlin/Heidelberg, Germany, 1996; pp. 423–437. [Google Scholar]
- Gao, Y.; Pan, Y.; Wang, T.; Yao, Y.; Yuan, W.; Zhu, X.; Wang, K. MicroRNA-99a-3p/GRP94 axis affects metastatic progression of human papillary thyroid carcinoma by regulating ITGA2 expression and localization. Acta Biochim. Biophys. Sin. 2021, 53, 1650–1661. [Google Scholar] [CrossRef]
- Yin, Y.; Zhang, J.; Ma, T.; Chen, D.; Lu, D. miR-1205/DNAJB1 reverses docetaxel chemoresistance in human triple negative breast carcinoma cells via regulation of mutp53/TAp63 signaling. Acta Biochim. Biophys. Sin. 2022, 54, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zou, W.; Meng, X.; Ji, J.; Wang, X.; Shu, H.; Chen, Y.; Pan, D.; Wang, K.; Zhou, F. Elaiophylin Inhibits Tumorigenesis of Human Uveal Melanoma by Suppressing Mitophagy and Inducing Oxidative Stress via Modulating SIRT1/FoxO3a Signaling. Front. Oncol. 2022, 12, 788496. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, X.; Zhu, X.; Ji, J.; Zhong, H.; Li, X.; Zhao, H.; Xie, G.; Wang, K.; Shu, H.; Wang, X. Erdafitinib Inhibits Tumorigenesis of Human Lung Adenocarcinoma A549 by Inducing S-Phase Cell-Cycle Arrest as a CDK2 Inhibitor. Molecules 2022, 27, 6733. https://doi.org/10.3390/molecules27196733
Meng X, Zhu X, Ji J, Zhong H, Li X, Zhao H, Xie G, Wang K, Shu H, Wang X. Erdafitinib Inhibits Tumorigenesis of Human Lung Adenocarcinoma A549 by Inducing S-Phase Cell-Cycle Arrest as a CDK2 Inhibitor. Molecules. 2022; 27(19):6733. https://doi.org/10.3390/molecules27196733
Chicago/Turabian StyleMeng, Xinmin, Xue Zhu, Jiali Ji, Hongqin Zhong, Xiyue Li, Hongqing Zhao, Guijuan Xie, Ke Wang, Hong Shu, and Xun Wang. 2022. "Erdafitinib Inhibits Tumorigenesis of Human Lung Adenocarcinoma A549 by Inducing S-Phase Cell-Cycle Arrest as a CDK2 Inhibitor" Molecules 27, no. 19: 6733. https://doi.org/10.3390/molecules27196733
APA StyleMeng, X., Zhu, X., Ji, J., Zhong, H., Li, X., Zhao, H., Xie, G., Wang, K., Shu, H., & Wang, X. (2022). Erdafitinib Inhibits Tumorigenesis of Human Lung Adenocarcinoma A549 by Inducing S-Phase Cell-Cycle Arrest as a CDK2 Inhibitor. Molecules, 27(19), 6733. https://doi.org/10.3390/molecules27196733